
大麻素2型受体拮抗剂对激活态BV2小胶质细胞的免疫调节作用
2016-07-10 04:35楼之茵赵忠新
同济大学学报(医学版) 2016年2期
李 琳, 楼之茵, 程 洁, 赵忠新
(1. 上海交通大学医学院附属新华医院神经内科,上海 200092; 2. 第二军医大学附属长征医院神经内科,上海 200003)
·基础研究·
大麻素2型受体拮抗剂对激活态BV2小胶质细胞的免疫调节作用
李 琳1, 楼之茵1, 程 洁1, 赵忠新2
(1. 上海交通大学医学院附属新华医院神经内科,上海 200092; 2. 第二军医大学附属长征医院神经内科,上海 200003)
目的 观察大麻素2型受体(cannabinoid 2 receptor, CB2R)拮抗剂对BV2小胶质细胞免疫调节功能的影响。方法 使用适量浓度的IFN-γ刺激激活BV2小胶质细胞,建立模拟EAE炎性环境的细胞模型,比较静息态BV2细胞组、激活态BV2细胞组和大麻素2型受体拮抗剂SR144528A(SR2)干预组CB2R mRNA和蛋白的表达情况,用ELISA方法检测细胞因子和趋化因子的浓度,MTT法检测细胞增殖率。结果 IFN-γ激活的BV2小胶质细胞CB2R mRNA和蛋白的表达均高于静息组(P<0.05);使用SR2干预激活的BV2小胶质细胞,可降低其CB2R mRNA和蛋白的表达(P<0.05);SR2可显著上调激活态BV2细胞致炎因子IFN-γ、IL-17、IL-6和TNF-α的水平,促进BV2小胶质细胞增殖和NO的释放(P<0.05),同时显著下调IL-4和MCP-1的浓度,对IL-1β、IL-10、CX3CL1无调节作用。结论 CB2R参与了小胶质细胞介导的炎症反应,CB2R在调节Th1/Th17/Th2细胞因子网络平衡中发挥了一定的作用。
大麻素2型受体; 小胶质细胞; 细胞因子; 趋化因子
小胶质细胞是中枢神经系统内最主要的炎症效应细胞,在神经免疫性疾病的发生发展中起重要作用。小胶质细胞在多发性硬化(multiple sclerosis, MS)病理过程中的作用尚不清楚。过去认为小胶质细胞在MS中释放炎性因子,促进白细胞活化和渗入中枢神经系统,促进炎症反应,具有神经毒性作用;然而近来的研究表明小胶质细胞可以分泌神经营养因子、清除变性髓鞘和坏死细胞、修复髓鞘,促进中枢神经的修复[1-2]。因此,抑制小胶质细胞“有害的”免疫应答,加强“有益的”免疫调节功能,使其在MS中充当“保护与修复者”角色,是改善MS临床症状的可能干预机制之一。
大麻素是从印度大麻里发现的一组萜酚类化合物,其神经保护作用最早在1996年被发现,但其神经保护的机制尚不完全清楚。人体内主要存在两种大麻素受体: 大麻素1型受体(cannabinoid 1 receptor, CB1R)和大麻素2型受体(cannabinoid 2 receptor, CB2R)。CB1R主要分布在中枢神经系统,参与运动与体位的控制、感知觉、情感、记忆、认知以及内分泌调节,CB2R大多分布在外周与免疫相关的细胞上,中枢神经系统内小胶质细胞和星形胶质细胞均有表达,参与免疫应答和炎症反应[3]。目前大多数观点认为CB2R介导的是抗炎效应[4],但在一项对大鼠小胶质细胞系的研究中发现,内源性大麻素2-AG可通过CB2R促进小胶质细胞增殖进而促进炎症反应[5]。为了进一步完善CB2R功能的证据,本研究采用细胞培养的方法,选用BV2小胶质细胞,研究CB2受体拮抗剂SR144528A(SR2)对BV2小胶质细胞免疫功能的影响。
1 材料与方法
1.1 材料与试剂
小鼠BV2细胞系购自上海复祥生物科技有限公司,DMEM培养液、胎牛血清购自美国Gibco公司,IFN-γ购自英国Abcam公司,TRIzol试剂购自美国Invitrogen公司,cDNA合成应用反转录试剂盒购自日本TaKaRa公司,成分蛋白抽提试剂盒购自美国Millipore公司,兔抗小鼠CB2多克隆抗体购自美国Cayman Chemical公司,兔抗小鼠GAPDH单克隆抗体购自美国Epitomics公司,MTT购自美国Sigma-Aldrich公司,一氧化氮(nitric oxide,NO)检测试剂盒购自上海碧云天公司。细胞因子和趋化因子ELISA试剂盒: IL-1β ELISA Kit购自美国Invitrogen公司,IL-4、IL-10、IL-17 ELISA Kit购自美国eBioscience公司,IL-6、TNF-α、IFN-γ、MCP-1 ELISA Kit购自美国PeproTech公司,CX3CL1 ELISA Kit购自美国R&D Systems公司。
1.2 BV2细胞培养及实验分组
BV2细胞用含10%胎牛血清的DMEM/高糖培养基,置于37℃、5%CO2和95%空气培养箱培养。当细胞达到95%融合时,以0.25%的胰酶消化细胞并进行传代。调整BV2细胞悬液中细胞密度为3.5×105/ml,接种于相应的细胞培养板中。取培养2~3d,处于对数生长期的BV2细胞进行实验。实验分为3组。静息态BV2细胞组: BV2+溶剂对照(PBS∶DMSO=1∶1);激活态BV2细胞组: BV2+IFN-γ(100U/ml)+溶剂对照;SR2干预组: BV2+IFN-γ(100U/ml)+ SR2(1μmol/L),处理24h。
1.3 Real-Time PCR法检测不同组别BV2细胞CB2R mRNA的表达
采用TRIzol试剂进行总RNA提取。根据Gene bank提供的小鼠CB1R cDNA序列和β-actin cDNA序列,通过Primer Premier 5.0软件设计引物。CB2R上游引物: 5′- CGGCTAGACGTGAG-GTTGGC-3′,下游引物: 5′- GGCTCCTAGGTG-GTTTTCAC -3′,扩增产物长度为 338bp;作为内参的β-actin上游引物: 5′-AGCCATGTACGTAGC-CATCC-3′,下游引物: 5′-CTCTCAGCTGTGGTG-GTGAA-3′,扩增产物长度为228bp。应用反转录试剂盒扩增组织样本cDNA。95℃ 90s变性,60℃ 90s退火,72℃ 120s延伸,共循环40次。以参照基因作为标准进行相对定量,采用参照基因的ΔCT法分析各组CB2R基因表达的相对比率。
1.4 Western印迹法检测不同组别BV2细胞CB2R蛋白的表达
采用成分蛋白抽提试剂盒抽提细胞膜蛋白,二辛可宁酸(bicinchonininc acid, BCA)法进行蛋白定量。电泳的膜蛋白上样量30μg,将蛋白于12%聚丙烯酰胺凝胶电泳分离,电转至PVDF膜上,5%BSA封闭1h后,分别加入兔抗CB2R(1∶300)和兔抗GAPDH(1∶1000),4℃孵育过夜漂洗,加相应荧光偶联的抗兔二抗(1∶1 200),室温孵育1h,洗膜后使用Odyssey近红外双色激光成像系统扫描获取相应WB谱带光密度值,CB2R表达量以CB2R与GAPDH整合光密度比值表示。
1.5 ELISA方法检测细胞因子和趋化因子浓度
使用ELISA试剂盒对干预后细胞上清液IL-1β,IL-4、IL-10、IL-17、IL-6、TNF-α、IFN-γ的含量进行测定。稀释标准品,使得稀释后各孔加样量均为50μl,浓度分别为240、160、80、40、20ng/L。分别设空白孔、待测样品孔。将样品加于酶标板孔底部(5倍稀释),轻晃混匀。经温育、配液、洗涤、加酶、显色等操作,最终加入终止液终止反应,以空白孔调零,测定450nm波长各孔吸光度值(D450)。
1.6 硝酸还原法(Griess法)测定NO浓度
收集各组细胞上清液,以50μl/孔依次加入96孔板,按50μl/孔加入Griess Reagent Ⅰ,室温下避光放置5min;按50μl/孔加入Griess Reagent Ⅱ,室温下避光放置5min;用酶标仪在540nm波长处测定吸光度值(D540),以系列浓度的亚硝酸钠标准溶液绘制标准曲线,计算样品中对应的NO浓度。
1.7 四唑盐(MTT)检测
将BV2细胞以3×106/ml密度接种于96孔细胞培养板,每孔加样200μl,分别给予相应刺激24h,每份样本均进行6复孔检测。各孔加MTT 溶液10μl(终浓度为0.5mg/ml),继续培养4h,然后取出培养板,弃上清液,每孔加100μl DMSO,稍振荡使颗粒完全溶解,置酶标仪以570nm波长读取D570值。细胞增殖率=检测组D570值/对照组D570值×100%。
1.8 统计学处理
2 结 果
2.1 SR2对激活态BV2小胶质细胞CB2R mRNA和蛋白表达的影响
结果显示,IFN-γ激活的BV2细胞中CB2R mRNA 和蛋白的表达分别为静息组的2.59倍和1.83倍(P<0.05)。使用SR2干预IFN-γ激活的BV2细胞,细胞中CB2R mRNA和蛋白的表达分别为激活态细胞的31%和47%,表明SR2可降低其CB2R mRNA 和蛋白的表达(P<0.05),见图1。

图1 不同组别BV2小胶质细胞CB2R mRNA和蛋白的变化Fig.1 Changes of CB2R mRNA and protein expression in BV2 cells of different groupsA: CB2R mRNA表达;B:CB2R蛋白表达1: 静息态BV2细胞组;2: 激活态BV2细胞组;3: SR2干预组
2.2 SR2对激活态BV2细胞的细胞因子与趋化因子的影响
与对照组相比,经IFN-γ(100U/ml)处理后,BV2小胶质细胞细胞因子IFN-γ、IL-17、IL-1β、IL-6、TNF-α和趋化因子MCP-1和CX3CL1的浓度均显著增高(P<0.05);而IL-4的浓度显著降低(P<0.05)。IFN-γ激活的BV2小胶质细胞经SR2(1μmol/L)干预后,与溶剂对照组相比,IFN-γ、IL-17、IL-6和TNF-α的水平均显著升高(P<0.05);IL-4和MCP-1的水平则显著降低(P<0.05);IL-1β、IL-10和CX3CL1的浓度差异无统计学意义(P>0.05)。上述结果显示,SR2在显著上调IFN-γ、IL-17、IL-6和TNF-α水平的同时可显著下调IL-4和MCP-1的浓度,对IL-1β、IL-10、CX3CL1无调节作用,见表1。
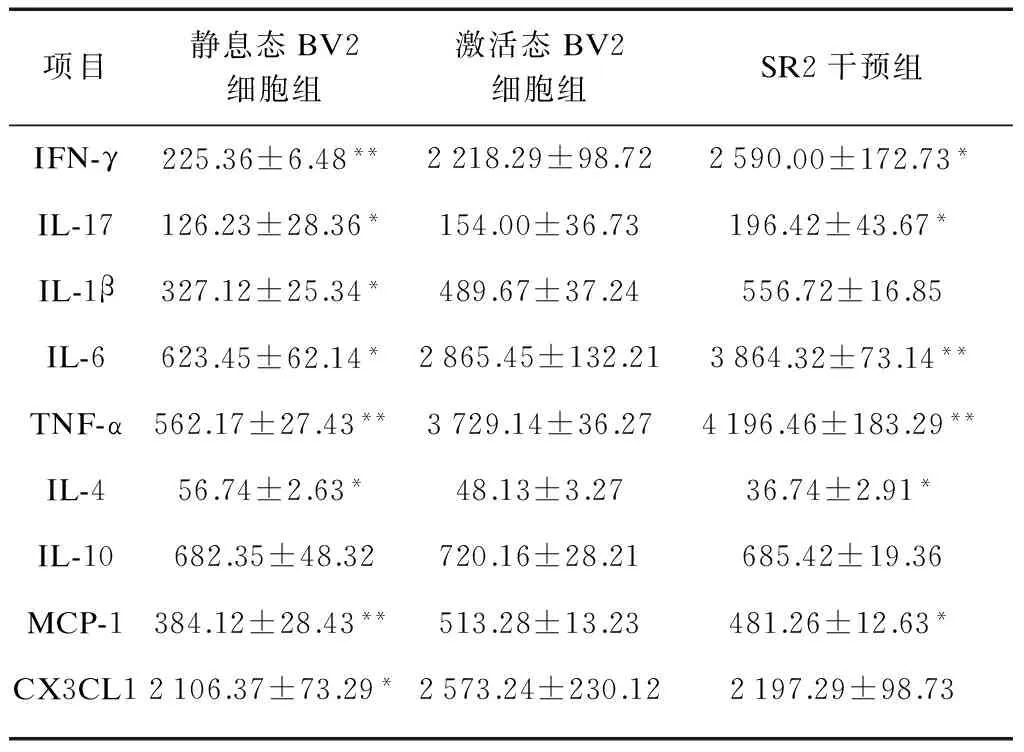
表1 不同干预对BV2小胶质细胞细胞因子和趋化因子的影响
与激活态BV2细胞组相比,*P<0.05,**P<0.01
2.3 SR2对激活态BV2 小胶质细胞中NO 分泌的影响
通过Griess法对激活态不同组别BV2小胶质细胞NO释放的测定结果显示,BV2细胞组NO释放量为(2.98±0.43)μmol/L,激活态BV2细胞组NO释放量为(25.75±1.89)μmol/L,SR2干预组NO释放量为(46.15±1.73)μmol/L,结果表明激活态BV2小胶质细胞NO释放量增加(P<0.05),SR2能显著增加激活态BV2小胶质细胞NO的释放(P<0.05)。
2.4 SR2对激活态BV2小胶质细胞增殖率的影响
通过MTT实验比较不同组别BV2小胶质细胞的增殖率,静息态BV2细胞组D570为0.367±0.031,激活态BV2细胞组D570为0.308±0.024,增殖率为84%;SR2干预组D570为0.444±0.026,增殖率为122%,与IFNγ激活态BV2细胞组相比,差异有统计学意义(P<0.05)。结果提示IFN-γ可促进BV2小胶质细胞增殖,SR2可促进IFN-γ活化的BV2小胶质细胞增殖(P<0.05)。
3 讨 论
病理学研究发现,在MS中枢病灶内,有大量活化的小胶质细胞,提示小胶质细胞在MS的发生发展中起重要作用[6]。最近的研究[7]表明,活化的小胶质细胞可以分为两种表型。(1) 经典的M1型: 发挥致炎作用,释放白介素(IL-1β、IL-17、IL-6)和诱导NO合成,具有神经毒性作用,可引起继发性神经损伤,细胞死亡和脱髓鞘,引起神经变性。(2) 具有神经保护作用的M2型: 可释放抗炎因子如IL-10、IL-4、NGF。经典型小胶质细胞活化往往伴随着MS的恶化,而调节小胶质细胞的功能可能改善疾病的预后[8]。
研究[9]表明,MS中存在内源性大麻素系统(endocannabinoid system, ECS)功能失调,有关ECS在MS中的作用逐渐得到广泛关注并成为新的研究热点。CB2R主要分布在外周与免疫相关的细胞上,在正常的中枢神经系统表达水平较低[10-11],Zhang等[12]对MS患者尸检标本检测发现,在MS慢性非活性斑块中内皮的CB2R表达最多。Malek等[13]发现,用大麻素受体激动剂安南得迈预处理可以降低脂多糖引起的小胶质细胞NO释放,减少小胶质细胞向有神经毒性作用M1表型转变,而这一效应是通过作用于CB2受体实现的。CB2R激动剂可以降低活化的巨噬细胞和脑微血管内皮细胞中多种致炎因子的基因表达[14]。Lau等[15]的研究中,通过对EAE小鼠模型的观察,发现SR2在EAE病程高峰期加重其神经功能缺损;影响脾脏自发性和抗原特异性T细胞增殖,扰乱细胞因子、炎性因子和趋化因子之间的网络平衡[15]。为排除复杂内环境的影响,本研究使用适量的IFN-γ激活BV2小胶质细胞,建立模拟EAE炎性环境的细胞模型,观察激活态BV2小胶质细胞CB2R的表达和免疫炎性反应的变化以及抑制CB2R后激活态BV2小胶质细胞的免疫炎性反应的变化。
本研究表明,IFN-γ刺激可诱导BV2小胶质细胞CB2R的表达显著升高,进一步证实了小胶质细胞表达的CB2R参与了炎症反应。
为了完善CB2R功能的证据,本研究用SR2特异性拮抗了激活态BV2小胶质细胞CB2R的表达,发现CB2R抑制可显著上调激活态BV2细胞分泌的促炎细胞因子IFN-γ、IL-17、IL-6和TNF-α水平,同时显著下调IL-4和趋化因子MCP-1的浓度,对IL-1β、IL-10和CX3CL1无显著调节作用。这些结果提示: CB2R在调节Th1/Th17/Th2细胞因子网络平衡中发挥一定作用,可能成为诱导“致炎性”的小胶质细胞向“抗炎性”小胶质细胞转变的治疗靶点。
在MS炎症病灶中,NO的浓度明显增高。NO主要由可诱导型一氧化氮合酶(inducible form of nitric oxide synthase, iNOS)合成,小胶质细胞、巨噬细胞、激活的星形胶质细胞均表达iNOS。NO能直接影响血脑屏障的通透性、损害少突胶质细胞、介导脱髓鞘、阻滞神经传导、促进神经元的凋亡。Merighi等[16]发现CB2R受体激动剂JWH-015可以抑制小胶质细胞产生NO。通过对激活态不同组别BV2小胶质细胞NO释放的观察,本研究发现CB2R受体拮抗剂SR2能显著增加BV2小胶质细胞NO的释放,提示CB2R可能与减少小胶质细胞NO分泌有关。
小胶质细胞活化和聚集是中枢神经系统关键的致病因素[17],研究发现CB2R可控制神经细胞和非神经细胞的增殖、分化和存活,激活CB2R可诱导小胶质细胞的抗炎表型,减少LPS刺激诱导的小胶质细胞迁移。Tang等[18]发现,激活小胶质细胞的CB2R受体可以显著降低原代大鼠胶质细胞p-ERK和p-p38 MAPK蛋白表达,抑制小胶质细胞的增殖与活化[18]。本研究发现,SR2可促进IFN-γ活化的BV2小胶质细胞增殖,CB2R可能通过抑制BV2小胶质细胞的增殖发挥其免疫调节作用。
由于ECS对机体的影响广泛而复杂,本实验针对CB2R的免疫调节作用做了初步探讨,我们将在CB2R细胞内信号转导通路中展开深入研究。
[1] Bogie JF1, Stinissen P, Hendriks JJ. Macrophage subsets and microglia in multiple sclerosis[J]. Acta Neuropathol, 2014,128(2): 191-213.
[2] Napoli I, Neumann H. Protective effects of microglia in multiple sclerosis[J]. Exp Neurol, 2010,225: 24-28.
[3] Cabral GA, Ferreira GA, Jamerson MJ.Endocann-abinoids and the immune system in health and disease[J]. Handb Exp Pharmacol, 2015,231: 185-211.
[4] Malfitano AM, Basu S, Maresz K, et al. What we know and do not know about the cannabinoid receptor 2(CB2)[J].Semin Immunol, 2014,26(5): 369-379.
[5] Carrier EJ, Kearn CS, Barkmeier AJ, et al. Cultured rat microglial cells synthesize the endocannabinoid 2-arachidonylglycerol, which increases proliferation via a CB2 receptor-dependent mechanism[J]. Mol Pharm-acol, 2004,65: 999-1007.
[6] Bogie JF, Stinissen P, Hendriks JJ. Macrophage subsets andmicroglia in multiple sclerosis[J]. Acta Neuropathol, 2014,128(2): 191-213.
[7] Tam WY, Ma CH. Bipolar/rod-shaped microglia are proliferating microglia with distinct M1/M2 phenotypes[J]. Sci Rep, 2014,4: 7279.
[8] Gao Z, Tsirka SE. Animalmodels of MS reveal multiple roles of microglia in disease pathogenesis[J]. Neurol Res Int, 2011,2011: 383087.
[9] Rossi S1, Bernardi G, Centonze D. The endocann-abinoid system in the inflammatory and neurodegene-rative processes of multiple sclerosis and of amyotr-ophic lateral sclerosis[J].Exp Neurol, 2010,224(1): 92-102.
[10] Martín-Couce L, Martín-Fontecha M, Palomares O, et al. Chemical probes for the recognition of cannabinoid receptors in native systems[J]. Angew Chem Int Ed Engl, 2012,51(28): 6896-6899.
[11] Li Y, Kim J. Neuronal expression of CB2 cannabinoid receptor mRNAs in the mouse hippocampus[J]. Neuroscience, 2015,311: 253-267.
[12] Zhang H, Hilton DA, Hanemann CO, et al. Cannabinoid receptor and N-acyl phosphatidyletha-nolamine phospholipase D-evidence for altered expre-ssion in multiple sclerosis[J]. Brain Pathol, 2011,21(5): 544-557.
[13] Malek N, Popiolek-Barczyk K, Mika J,et al. Anandamide, acting via CB2 receptors, alleviates LPS-induced neuroinflammation in rat primary microglial cultures[J]. Neural Plast, 2015,2015: 130639.
[14] Persidsky Y, Fan S, Dykstra H,et al. Activation of cannabinoid type two receptors (CB2) diminish inflammatory responses in macrophages and brain endothelium[J].J Neuroimmune Pharmacol, 2015,10(2): 302-308.
[15] Lou ZY, Chen C, He Q,et al. Targeting CB(2) receptor as a neuroinflammatory modulator in experime-ntal autoimmune encephalomyelitis[J].Mol Immunol, 2011,49(3): 453-461.
[16] Merighi S,Gessi S, Varani K, et al.Cannabinoid CB(2) receptors modulate ERK-1/2 kinase signalling and NO release in microglial cells stimulated with bacterial lipopolysaccharide[J]. Br J Pharmacol, 2012,165(6): 1773-1788.
[17] Supramaniam V, Vontell R, Srinivasan L,et al. Microglia activation in the extremely preterm human-brain[J]. Pediatr Res, 2013,73(3): 301-309.
[18] Tang J, Tao Y, Tan L,et al. Cannabinoid receptor 2 attenuates microglial accumulation and brain injury following germinal matrix hemorrhage via ERK dephosphorylationinvivoandinvitro[J]. Neuropha-rmacology, 2015,95: 424-433.
Immunoregulation of cannabinoid 2 receptor antagonist on activated microglia BV2 cells
LILin1,LOUZhi-yin1,CHENGJie1,ZHAOZhong-xin2
(1. Dept. of Neurology, Xinhua Hospital, Shanghai Jiaotong University School of Medicine, Shanghai 200092, China;2. Dept. of Neurology, Changzheng Hospital, Second Military Medical University, Shanghai 200003, China)
Objective To study the effect of cannabinoid 2 receptor (CB2R) antagonist on the activated microglia BV2 cells. Methods Stimulated by IFN-γ, BV2 cells were exposed to CB2R antagonist SR144528A (SR2). The expression of CB2R mRNA and protein were measured by SYBR Green-based Real-Time RT-PCR and Western-blotting, respectively; the levels of cytokines and chemokines were measured with ELISA; and cell proliferation was tested by MTT. Results Stimulated by IFN-γ, the expression of CB2R in BV2 cells increased significantly (P<0.05). SR2 decreased the expression of CB2R, increased the production of inflammatory cytokines IFN-γ, IL-17, IL-6 and TNF-α as well as NO, promoted cell proliferation, and the production of IL-4 and MCP-1 were decreased; but had no effects on the secretion of IL-1β, IL-10 and CX3CL1. Conclusion CB2R is involved in the inflammatory response mediated by microglia BV2 cells, and CB2R plays a role in the regulation of Th1/Th17/Th2 cytokine network balance.
cannabinoid 2 receptor; microglia; cytokines; chemokines
2015-12-13
国家自然科学青年基金(81200921)
李 琳(1974—),女,主治医师,硕士研究生.E-mail: lilin7505@126.com
赵忠新.E-mail: zhaozx@medmail.com.cn
R 744.5
A
1008-0392(2016)02-0014-05
猜你喜欢
生物化工(2022年4期)2022-09-20
昆明医科大学学报(2021年2期)2021-03-29
世界最新医学信息文摘(2021年55期)2021-01-07
神经损伤与功能重建(2020年10期)2020-12-23
神经损伤与功能重建(2020年11期)2020-12-01
神经损伤与功能重建(2018年2期)2018-02-01
儿童故事画报·发现号趣味百科(2017年1期)2017-06-01
华声(2016年20期)2016-11-19
中国医药生物技术(2015年4期)2015-12-26
中华胰腺病杂志(2015年5期)2015-12-08
