
3种香薷属植物染色体数目与核型分析
2016-07-04 08:01:24巴罗菊李志敏陈光富邓洪平
西北植物学报 2016年5期
巴罗菊,李志敏,陈光富,邓洪平*
(1 西南大学 生命科学学院,三峡库区生态学环境教育部重点实验室,重庆 400715;2 云南师范大学 生命科学学院,云南呈贡 650500)
3种香薷属植物染色体数目与核型分析
巴罗菊1,李志敏2,陈光富2,邓洪平1*
(1 西南大学 生命科学学院,三峡库区生态学环境教育部重点实验室,重庆 400715;2 云南师范大学 生命科学学院,云南呈贡 650500)
摘要:参照植物根尖细胞学研究的方法标准,对香薷属3种(5个居群)植物进行核形态学分析。结果表明:(1)从染色体数目看,密花香薷2居群染色体数目2n=16;野苏子2居群染色体数目2n=20,染色体数目和倍性与前人报道的一致;毛穗香薷染色体数目2n=10为首次报道。(2)聚类分析结果显示,3种(5居群)植物中野苏子和密花香薷亲缘关系较近;结合现有报道数据分析表明,该属植物仅有2种倍性(二倍体和四倍体),且二倍体占主导地位。(3)核型参数分析表明:密花香薷的稻城无名山居群1核型公式为2n=2x=16=14 m+2 sm,居群2为2n=2x=16=16 m,着丝粒指数(CI)分别为39.57和42.32,不对称系数AI值分别为2.75和2.87,核型不对称性都为1A型;毛穗香薷的核型公式为2n=2x=10=10 m,着丝粒指数(CI)为41.76,不对称系数AI值为5.25,核型不对称性为1B型;野苏子的昆明西山居群核型公式为2n=2x=20=14 m+6 sm,聂拉木樟木沟居群为2n=2x=20=16 m+4 sm,着丝粒指数(CI)分别为38.49和40.97,不对称系数AI值为4.20和4.30,核型不对称性为1B型和2B型。
关键词:香薷属;核型;染色体数目;聚类分析;AI值
香薷属(Elsholtzia)在分类上隶属唇形科(Lamiaceae)野芝麻亚科下的刺蕊草族,全属约40种,是以中国为主要分布区的类群,中国有33种、15变种和3变型[1]。香薷属植物为重要的中药植物,开发前景十分广阔[2-3]。此外,在园艺、食用等方面也有重要的价值[4],少部分香薷属植物还是良好的蜜源植物[5]。
关于香薷属植物的研究在植物化学与药理学、植物分类学、生理学和功能生态学上有相关报道[6-8]。就该属植物的分类学研究,多集中于形态分类方面。植物染色体核型作为植物分类中的细胞学数据,对植物区系起源及进化具有重要意义[9],也是分类和鉴别的重要依据[10],且Inceer等在2004年提出,以核型数据为基础的聚类分析能更好地考察物种间的亲缘关系[11]。目前已有部分香薷属类群染色体数据的报道(表1),香薷[E. ciliata (Thunb.)Hyland.]、海州香薷(E. splendensNakai)等4种香薷属植物也有染色体长度范围记录[15],但就本属染色体核型参数并未进行分析比较。本实验从核型数据和聚类分析2个方面分别对野苏子[E. flava (Benth.)Benth.]、毛穗香薷(E. eriostachyaBenth.)和密花香薷(E. densaBenth.)3种(5居群) 香薷属植物进行了研究,且核型数据均为首次报道。这为研究香薷属植物系统演化提供了重要的细胞学数据,并为该属植物物种鉴定、引种栽培、杂交和药理成分分析等方面奠定了基础。
1材料和方法
1.1材料
实验材料为野生种,均采自高海拔地区(表2),分布于云贵及川藏高原,具有明显的横断山脉植物地理区系特征[21]。采样地海拔梯度及纬度差异明显,具一定代表性。其中,密花香薷居群1、居群2与毛穗香薷由中国科学院昆明植物研究所孙航研究员采集及鉴定、野苏子昆明居群由云南师范大学陈光富老师采集及鉴定、野苏子樟木沟居群由昆明植物研究所周卓老师采集及鉴定,在此感谢几位老师对本实验的帮助。凭证标本保存于中国科学院昆明植物研究所标本馆(KUN)。
1.2方法
常规实验方法包括种子发芽、取材,预处理、固定、解离、染色、镜检,拍照。针对本实验材料的特征,对实验处理进行了一定改良,具体如下:种子在25~30 ℃条件下培养萌发,切取生长旺盛的植物根尖,长度一般为0.5~1.5cm,常温下用0.002~0.003mol/L8-羟基喹啉混合液避光处理1~2h,用卡诺液(冰乙酸: 无水乙醇=1∶3)在4 ℃冰箱中固定18~24h。在60 ℃恒温水浴锅中用1mol/LHCl解离13~16min,卡宝品红染色液染色24h以上,但染色时间不宜过长,否则达不到预期效果。常规染色体压片法压片,在显微镜下观察并拍下较好的间期、前期、中期细胞图片。
依据Tanaka的标准,判断细胞间期核与前期核的类型[22-23]。统计45个以上中期细胞,85%以上的细胞具有相同染色体数目,从而确定染色体数目[24]。至少取5个分散较好的细胞进行测量,染色体类型按Levan等的方法分析[25],核型不对称性参照Stebbins及Paszko提出的较为精确的AI值[26-27],分析参照李懋学等的方法[24]。对核型数据处理参考Hamideh等提出的常规统计分析方法[28],而亲缘关系分析采用Inceer等的方法对11个核型参数进行聚类分析[11]:倍性水平、随体染色体数目、染色体相对长度(RL)、染色体绝对长臂(LA)、染色体绝对短臂(SA)、染色体绝对长度(CL)、染色体组长(TCL)、臂比值(AR)、染色体内不对称系数(A1)、染色体间不对称系数(A2)、着丝粒指数(CI),以上分析均使用SPSS17.0软件包。

表1 已有报道的香薷属植物染色体数目和数据来源文献

表2 材料采集信息
2结果与分析
图1,A、B所示为间期核和前期核,根据Tanaka标准[25],间期核中有一些着色较深的染色体,得到该物种间期核为复杂染色体,有丝分裂前期染色体为中间型。
中期染色体形态如图1,C~G所示,中期染色体核型模式图如图1所示,对3种(5居群)植物分别挑选5个染色体分散较好的中期细胞进行核型分析。得到的核型参数如表4所示,结果表明:3种香薷属植物染色体属于sm或m型,核型不对称性有1A、1B和 2B等3种类型。染色体长度比在1.67~2.67之间变化;染色体平均长度(CL)值在1.65~2.75之间变化,以野苏子昆明西山居群为最短,毛穗香薷为最长。着丝粒指数(CI)值在38.49~42.32之间变化,不对称系数AI值在2.75~5.25之间变化,以毛穗香薷为最高,密花香薷为最低。本研究中,同一物种具有相同的染色体倍性和染色体数目,但不同居群间核型参数存在差异;首次发现香薷属植物中存在染色体数目2n=10;3种植物的核型数据均为首次报道,且在香薷属中首次使用AI值进行种间系统学分析。
2.1密花香薷(居群1和居群2)
密花香薷为直立草本,花、果期7~10月,生于林缘、高山草甸、林下、河边及山坡荒地。密花香薷稻城无名山居群1和居群2的植物体细胞染色体数目都为2n=16,但核型参数和核型特征存在差异(表3、表4)。其中,居群1核型公式2n=2x=14m+2sm,体细胞中期染色体由14条中部(m)和2条近中部(sm)染色体组成,从相对长度组成2n=8M2+8M1来看,密花香薷居群1由中长染色体(M2)和中短染色体(M1)2种染色体组成,这8组染色体相对长度范围为9.53%~ 15.41%、臂比范围为1.30~1.79、染色体长度比为1.78、染色体平均长度为1.81、着丝粒指数为39.57、不对称系数AI值为2.75、核型为1A型;居群2核型公式2n=2x=16m,体细胞中期染色体由16条中部(m)染色体组成,相对长度组成与居群1相同,相对长度范围为10.12%~15.29%、臂比范围为1.21~1.59、染色体长度比为1.67、染色体平均长度为1.74、着丝粒指数为42.32、不对称系数AI值为 2.87、核型与居群1相同为1A型。
2.2毛穗香薷
毛穗香薷为一年生草本,简单或自基部向上在节处均具短分枝,分枝能育,花、果期7~9月,中国产甘肃、四川、西藏、云南,生于高海拔地区的山坡草地。毛穗香薷四川稻城三堆居群植物体细胞染色体数目为2n=10,核型公式为2n=2x=10m,核型参数与核型特征见表3和表4。从核型公式看出,体细胞中期染色体全部由10条中部(m)染色体组成,从相对长度组成2n=2L+1M1+2S来看,毛穗香薷由长染色体(L)、中短染色体(M1)和短染色体(S)3种染色体组成,5组染色体相对长度范围为12.26%~30.20%、臂比范围为1.22~1.67、染色体长度比为2.67、染色体平均长度为2.75、着丝粒指数为41.76、不对称系数AI值为5.25、核型为1B型。
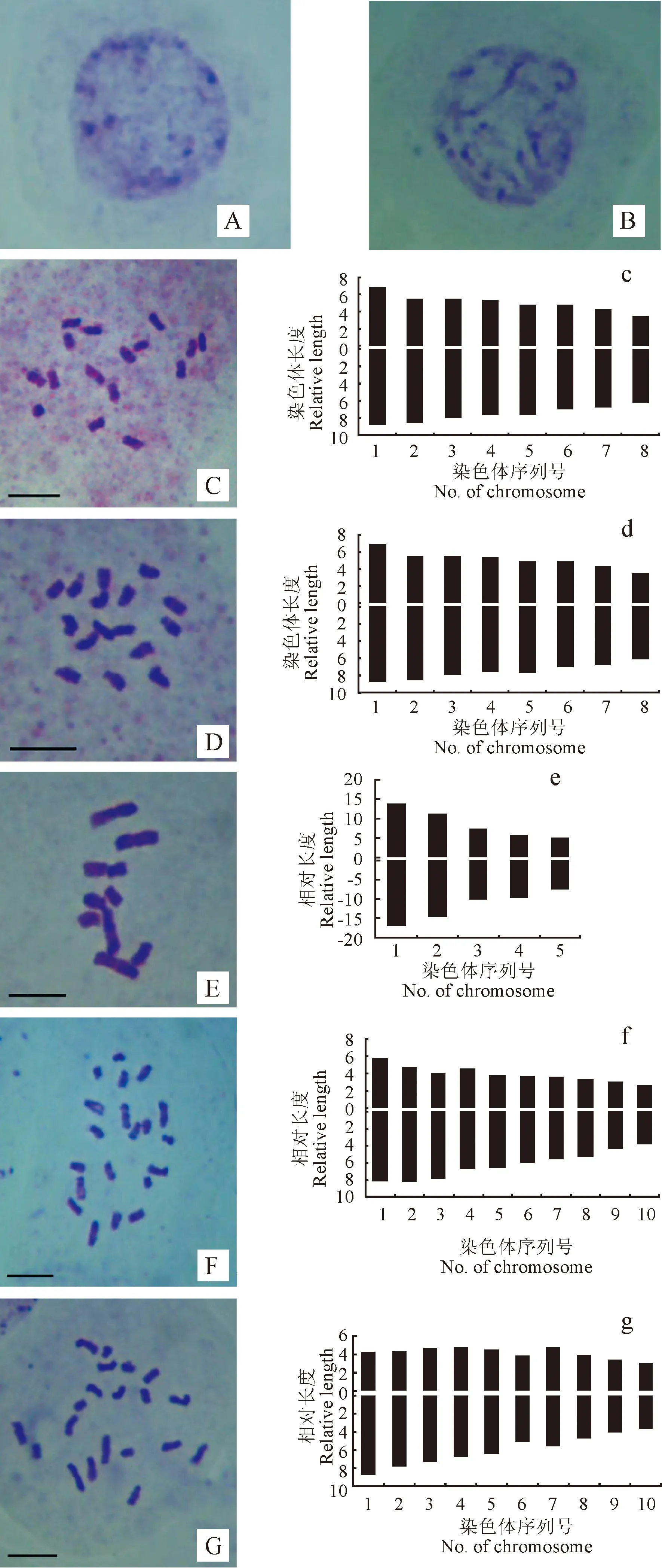
A.前期核;B.有丝分裂前期染色体;C~G.中期染色体:(C.密花香薷居群1,D.密花香薷居群2,E.毛穗香薷,F.野苏子居群1,G.野苏子居群2;)c~g.染色体核型模式图:(c.密花香薷居群1,d.密花香薷居群2,e.毛穗香薷,f.野苏子居群1,g.野苏子居群2。)标尺=5 μm图1 香薷属的前期核、有丝分裂前期染色体和中期染色体形态及核型模式图A. Mitotic interphase nuclei; B. Mitotic prophase; C-G were somatic metaphase: [C. Elsholtzia densa (population 1), D. E. densa (population 2), E. E. eriostachya; F. E. flava (population 1), G. E. flava (population 2);] c-g were chromosome ideogram: [c. E. densa (population 1), d. E. densa (population 2), e. E. eriostachya, f. E. flava (population 2), g. E. flava (population 2).] Scale bar=5 μmFig. 1 Cytological features and chromosome ideograms of Elsholtzia
2.3野苏子(居群1和居群2)
野苏子为直立半灌木,茎分枝,密被灰白色短柔毛,花期7~10月、果期9~11月,生于开阔耕地、路边、沟谷旁、灌丛中或林缘。野苏子昆明西山居群1和聂拉木樟木沟居群2的植物体细胞染色体数目都为2n=20,但核型参数和核型特征也存在差异(表3、表4)。其中,居群1核型公式为2n=2x=14m+6sm,体细胞中期染色体由14条中部和6条近中部染色体组成,染色体相对长度范围为6.18%~14.01%、臂比范围为1.46~1.99、染色体长度比为2.47、染色体平均长度为1.65、着丝粒指数为38.49、不对称系数AI值为4.20、核型为1B型;居群2核型公式为2n=2x=16m+4sm,体细胞中期染色体由16条中部和4条近中部染色体组成,相对长度范围为6.52%~12.79%、臂比范围为1.19~2.10、染色体长度比为2.37、染色体平均长度为2.04、着丝粒指数为40.97、不对称系数值AI为4.30、核型为2B。从相对长度组成来看,野苏子居群1为2n=4L+6M2+6M1+4S、居群2为2n=2L+10M2+4M1+4S,两居群染色体长度组成相同,都是由长染色体、中长染色体(M2)、中短染色体和短染色体4种染色体组成。
2.43种(5居群)香薷属植物聚类分析
对5居群植物进行聚类分析,讨论5居群间的进化关系,分析结果(图2)表明:阈值取20时聚为两支,野苏子和密花香薷的4个居群聚为一支,且密花香薷居群1与野苏子有更近一步的亲缘关系,毛穗香薷单独为一支。
3讨论
香薷属已有染色体数目报道的物种染色体数目有:2n=2x=16、18、20、32,2n=4x=32,基数为x=8、9、10、16,可见香薷属中,有二倍体和四倍体存在,且二倍体类群占据主导地位。而本属中的香薷有2n=2x=16、18,2n=4x=32的报道,染色体数目及倍性水平具不稳定性。Lewis等指出染色体可以通过重排、易位等发生进化[29],这与海拔、经纬度有着密切的关联。本研究中野苏子2居群染色体均为2n=20与Saggoo等所报道的一致,密花香薷2居群染色体均为2n=16与Zhang等报道的一致,而毛穗香薷2n=10和Saggoo等所报道的2n=16存在差异[13,15],说明毛穗香薷种内染色体数据存在不稳定性。

图2 香薷属3种(5居群)染色体形态特征聚类分析图Fig. 2 Cluster analysis dendrogram of three Elsholtzia species five populations investigated

分类群Taxon染色体序号Chromosomenumber相对长度Relativelengths/%臂比Armratio类型Form相对长度指数Relativelengthindex(I.R.L)密花香薷居群1E.densapopulation1115.411.30m1.23(M2)213.881.59m1.11(M2)313.301.46m1.06(M2)412.871.44m1.03(M2)512.371.62m0.99(M1)611.701.47m0.94(M1)710.941.59m0.88(M1)89.531.79sm0.76(M1)密花香薷居群2E.densapopulation2115.291.31m1.22(M2)214.251.43m1.14(M2)313.441.59m1.08(M2)412.651.30m1.01(M2)512.151.42m0.97(M1)611.451.30m0.92(M1)710.641.29m0.85(M1)810.121.21m0.81(M1)毛穗香薷E.eriostachya130.201.22m1.51(L)225.491.28m1.27(L)317.051.41m0.85(M1)415.011.67m0.75(S)512.261.46m0.61(S)野苏子居群1E.flavapopulation1114.011.47m1.40(L)212.711.73sm1.27(L)311.761.99sm1.18(M2)411.051.47m1.10(M2)510.221.76sm1.02(M2)69.521.68m0.95(M1)79.041.53m0.90(M1)88.381.58m0.84(M1)97.221.46m0.72(S)106.181.47m0.62(S)野苏子居群2E.flavapopulation2112.792.10sm1.28(L)211.891.90sm1.19(M2)311.851.70m1.19(M2)411.411.50m1.14(M2)510.771.47m1.08(M2)68.821.35m0.88(M1)710.141.19m1.01(M2)88.501.23m0.85(M1)97.241.23m0.72(S)106.521.26m0.65(S)

表4 香薷属3种(5居群)植物的核型参数
研究结果表明,野苏子和密花香薷各居群植物体细胞染色体数目均一致,但其染色体类型与相对长度组成却存在一定差异,密花香薷无名山居群1、居群2核型公式分别为2n=2x=16=14m+2sm和2n=2x=16=16m,野苏子昆明西山居群和聂拉木樟木沟居群分别为2n=2x=20=14m+6sm和2n=2x=20=16m+4sm;从各居群所处环境看,居群间海拔梯度差异较大,生境也存在明显差异,染色体类型组成上的差异可能是由此生境差异所造成;此外,Paszko[27]提出了更为精确的不对称性量化参数AI值,该参数综合考虑了染色体长短及着丝粒位置的变异情况,认为AI值越高,不对称性越强,植物居群越趋于进化。本实验采用AI值来表述该属植物的染色体不对称性,得出密花香薷海拔3 740m居群AI=2.75,海拔4 324m居群AI=2.87;野苏子中昆明居群1 950mAI=4.20,樟木沟居群2 230mAI=4.30 ,也不难看出类似的结果。这似乎表明海拔较高的居群具有较进化的特征,可能是环境(高海拔)恶劣所造成的。
3种植物核型不对称类型有密花香薷1A型、毛穗香薷1B型、野苏子1B型和2B型,早有学者提出“核型对称——原始,核型不对称——更进化”的观点[9,27,30]。因此核型类型为1A的密花香薷较其余2种更原始。对该属已有染色体数目报道的植物进行染色体资料统计发现,共有12种香薷属植物有染色体数目报道,从中不难发现该属仅存在二倍体和四倍体2种倍性,且多倍体所占比例较小(16.6%)。由于多倍体具有更高的基因杂合度,故在适应更广的生态幅时较有利[31],这可能是该属植物仅在中国广布的原因之一。当然,关于香薷属的研究还需结合分子生物学、植物地理学和古生物学等方面的手段进行深入研究。
此外,根据聚类分析可以看出,香薷属3种植物中,密花香薷和野苏子的亲缘关系相对较近,密花香薷2居群和野苏子2居群在种内也存在一定差异。由于缺乏国产香薷属植物其它种类的核型资料,而无法在分类和进化上作更进一步分析比较。但对3种(5居群)香薷属植物群染色体数目和核型分析的研究,可为香薷属植物细胞分类学研究积累资料及填补空缺,也为研究中国香薷属植物群落分布与地理位置差异的关系提供理论基础。
参考文献:
[1]中国科学院中国植物志编辑委员会. 中国植物志(66卷)[M]. 北京: 科学出版社, 1977.
[2]龚慕辛. 香薷的药理研究概况[J]. 北京中医,1997,16(6): 46-48.
GONGMX.OverviewofpharmacologicalresearchofElsholtzia[J]. Beijing Journal of Traditional Chinese Medicine, 1997, 16(6): 46-48.
[3]石进丽, 朱甘培. 中国香薷属植物的药用及开发前景[J]. 中药材, 1994, 17(12): 10-13.
SHIJL,ZHUGP.MedicinalUtilizationandDevelopingPromiseofGenusElsholtziainChina[J]. Journal of Chinese Medicinal Materials, 1994, 17(12): 10-13.
[4]李保印, 周秀梅, 郝峰鸽, 等. 我国香薷属植物研究进展[J]. 河南科技学院学报, 2012, 40(1): 37-44.
LIBY,ZHOUXM,HAOFG, et al.AdvancesandperspectivesinstudiesonElsholtziagenus[J]. Journal of Henan Institute of Science and Technology, 2012, 40(1): 37-44.
[5]瞿纯, 董霞. 香薷属蜜源植物的研究与利用[J]. 蜜蜂杂志, 2009, (8): 41-42.
[6]赵勇, 赵焱, 陈业高. 云南产香薷属植物挥发油的研究进展[J]. 云南师范大学学报(自然科学版), 2004, 25(1): 31-33.
ZHAOY,ZHAOY,CHENYG.ResearchprogressonessentialoilsofElsholtziainYunnanProvince[J]. Journal of Yunnan Normal University (NaturalSciencesEdition), 2004, 25(1): 31-33.
[7]胡珊梅, 范崔生. 四种香薷的花粉和果实形态扫描电镜比较观察[J]. 武汉植物学研究, 1995, 13(1): 95-96.
HUSM,FANCS.Thescanningelectronmicroscopeofpollenandfruitoffourspecies“Xiangru”[J]. Journal of Wuhan Botanical Research, 1995, 13(1): 95-96.
[8]曹素珍. 青藏高原两种香薷的适应性表型可塑性研究[D]. 兰州:兰州大学, 2008.
[9]洪德元. 植物分类学[M]. 北京: 科学出版社, 1990: 1-11.
[10]张雪婷, 杨文雄, 杨芳萍, 等. 6个紫花苜蓿材料的核型及其亲缘关系分析[J]. 西北植物学报,2011,31(4): 671-676.
ZHANGXT,YANGWX,YANGFP, et al.Karyotypeandclusteranalysisof6alfalfamaterials[J]. Acta Botanica Boreali-Occidentalia Sinica, 2011, 31(4): 671-676.
[11]INCEERH,BEYAZOGLUO.KaryologicalstudiesinTripleurospermum (Asteraceae,Anthemideae)fromnorth-eastAnatolia[J]. Botanical Journal of the Linnean Society, 2004, 146: 427-438.
[12]GILLLS.TheincidenceofpolyploidyintheWest-HimalayanLabiatae[J]. Revue de Cytologie et de Biologie Végétales, le Botaniste, 1984, 7: 5-16.
[13]SAGGOOMI,BIRSS.MeioticstudiesonsomeeastHimalayanmembersoffamilyLabiatae[J]. Journal of the Indian Botanical Society, 1986, 56: 304-309.
[15]张义贤, 上官铁梁, 平俊爱, 等. 山西产9种野生植物的染色体观察[J]. 广西植物, 1993, 13(2): 159-163.
ZHANGYX,SHANGGUANTL,PINGJA, et al.Chromosomeobservationof9wildplantspeciesfromShanXi[J]. Guihaia,1993, 13(2): 159-163.
[16]NISHIKAWAT.ChromosomecountsoffloweringplantsofHokkaido[J]. Journal of Hokkaido University of Education, 1985, 36: 25-40.
[17]ADAMKIEWICZE,BIJOKK.KaryologicalinvestigationsofElsholtziapatrini(Lep.)Garche[J]. Zeszyty Naukowe Wydzialu Biologii Nauko Ziemi Biologia, 1980, 2: 77-80.
[18]马兴华, 覃若林, 邢文斌. 新疆20种药用植物的染色体观察[J]. 西北植物学报, 1985, 5(2): 149-154.
MAXH,QINRL,XINGWB.ChromosomeobservationoftwentyspeciesofdrugplantsinXingjiang[J]. Acta Botanica Boreali-Occidentalia Sinica, 1985, 5(2): 149-154
[20]GILLBS,BIRSS,BEDIYS.CytologicalstudiesonwoodyEuphorbiaceaefromNorthandCentralIndia[J]. New Botanist, 1981, 8: 35-44.
[21]李锡文,李捷.横断山脉地区种子植物区系的初步研究[J]. 云南植物研究, 1993, 15(3): 217-231.
LIXW,LIJ.ApreliminaryfloristicstudyontheseedplantsfromtheregionofHengduanMountain[J]. Acta Botanica Yunnanica, 1993, 15(3): 217-231.
[22]TANAKAR.TypesofrestingnucleiinOrchidaceae[J]. Botanical Magazine Tokyo, 1971, 84: 118-122.
[23]TANAKAR.ThekaryotypetheoryandwidecrossingasanexampleinOrchidaceae[M]//:HONGDY.Plantchromosomeresearch, 1989.Hiroshima:ProceedingsoftheSino-JapaneseSymposiumonPlantChromosomes, 1987: 1-10.
[24]李懋学, 陈瑞阳. 关于植物核型的标准化问题[J]. 武汉植物学研究, 1985, 3(4): 297-302.
LIMX,CHENRY.ASuggestionontheStandardizationofKaryotypeAnalysisinPlants[J]. Journal of Wuhan Botanical Research, 1985, 3(4): 297-302.
[25]LEVANA,FEDGAK, et al.Nomenclatureforcentromericpositiononchromosomes[J]. Hereditas, 1964, 52: 201-220.
[26]STEBBINSGL.ChromosomalEvolutioninHigherPlants[M].ChromosomalEvolutioninHigherPlants, 1971: 72-123.
[27]PASZKOB.Acriticalreviewandanewproposalofkaryotypeasymmetryindices[J]. Plant Systematics and Evolution, 2006, 258: 39-48.
[28]HAMIDEHJ,HESAMZADEHSM,BABAYEVMS.KaryotypicstudiesofthreeThymus (Lamiaceae)speciesandpopulationsinIran[J].Caryologia International Journal of Cytology Cytosystematics & Cytogenetics, 2009, 62: 316-325.
[29]LEWISKR,JOHNB.ChromosomeMarker[M].London:Churchill, 1963.
[30]STEBBINSGL.Longevity,habitatandreleaseofgeneticvariabilityinthehigherplants[J]. Cold Spring Harbor Symposia on Quantitative Biology, 1958, 23: 365-378.
(编辑:宋亚珍)
Karyotype and Chromosome Numbers of Three Species fromElsholtzia
BALuoju1,LIZhimin2,CHENGuangfu2,DENGHongping1*
(1SchoolofLifeScience,KeyLaboratoryofEco-environmentsinThreeGorgesReservoirRegion,SouthwestUniversity,Chongqing400715,China; 2LifeScienceSchool,YunnanNormalUniversity,Chenggong,Yunnan650500,China)
Abstract:Standard plant root tip cytology research methods were used to study the chromosome number and karyotype of 5 populations of 3 Elsholtzia species. The result showed that:(1)the chromosome number of E. densa was 2n=16, and the chromosome number of E. flava was 2n=20, which were consistent with previous reports. Chromosome number of E. eriostachya was 2n=10, which is first reported here.(2) Cluster analysis results showed that E. flava has close relationship withE. densa. Only diploid and tetraploid were found in this genus and dominated by diploid.(3)Three conclusions were drawn by analysis of karyotype parameters: the karytypic formulae of E. densa Daocheng Wumingshan population 1 was 2n=2x=16=14m+2sm, and population 2 was 2n=2x=16=16m. CI of population 1 and population 2 were 39.57 and 42.32, AI were 2.75 and 2.87, respectively. The two populations have the same type of karyotype: 1A. The karytypic formulae of E. eriostachya was 2n=2x=10=10m, CI was 41.76, AI was 5.25 and the type of karyotype was 1B. The karytypic formulae of E. flava was 2n=2x=20=14m+6sm (Kunming population) or 2n=2x=20=16m+4sm (Nielamu population). CI of Kunming population and Nielamu population were 38.49 and 40.97, AI were 4.20 and 4.30, respectively. The karyotype types of Kunming population and Nielamu population were 1B and 2B.
Key words:Elsholtzia; karyotype; chromosome number; cluster analysis; AI
文章编号:1000-4025(2016)05-0923-07
doi:10.7606/j.issn.1000-4025.2016.05.0923
收稿日期:2016-03-11;修改稿收到日期:2016-05-17
基金项目:科技部国家科技基础平台国家标本平台——教学标本子平台运行服务项目(2005DKA21403-JK)
作者简介:巴罗菊(1990-),女,在读硕士研究生,主要从事植物系统进化与保护生物学研究。E-mail:luoju63@163.com。 *通信作者:邓洪平,教授,主要从事植物系统进化与保护生物学研究。E-mail:denghp@swu.edu.cn。
中图分类号:Q343.2+2
文献标志码:A
猜你喜欢
宁夏医学杂志(2020年3期)2021-01-21 08:23:24
商情(2016年39期)2016-11-21 08:45:54
新媒体研究(2016年19期)2016-11-18 19:48:34
大经贸(2016年9期)2016-11-16 16:16:46
中国市场(2016年33期)2016-10-18 12:16:58
科技视界(2016年20期)2016-09-29 12:32:48
企业导报(2016年9期)2016-05-26 21:08:14
哈尔滨医药(2015年2期)2015-12-01 03:57:21
中国当代医药(2015年9期)2015-03-01 02:02:12
郑州大学学报(医学版)(2015年1期)2015-02-27 14:50:35
