
普通小麦与粗山羊草正反杂交研究
2016-06-15 11:59张海泉
西北农林科技大学学报(自然科学版) 2016年4期
关键词:小麦
张海泉,杨 虹,郎 杰
(河北经贸大学 生物科学与工程学院,河北 石家庄 050061)
普通小麦与粗山羊草正反杂交研究
张海泉,杨虹,郎杰
(河北经贸大学 生物科学与工程学院,河北 石家庄 050061)
[摘要]【目的】 研究普通小麦与粗山羊草杂交后的成胚率、成苗率及F1代的田间表现,为提高小麦的杂交成功率提供参考。【方法】 以小麦矮败不育系(RH/CS)、中国春、辽春10号3个春性小麦及济南17和济麦20 2个冬性小麦为供试小麦材料,Y122、Y212、Y215为3个粗山羊草材料,先以小麦为母本,粗山羊草为父本构建正交组合,并在去雄后1~5 d连续授粉,分析授粉次数对授粉率的影响,再以粗山羊草为母本,普通小麦为父本构建反交组合,分析各组合下胚、胚乳的形成情况;对正交组合进行幼胚拯救后,统计各组合的成胚率和成苗率;并将幼苗移至大田中,观察F1代的田间表现。【结果】 小麦反交组合的授粉率(70.32%)远高于正交组合(8.51%),但正交和反交组合均未能形成种子,需要幼胚拯救才能获得后代;重复授粉可以提高授粉率;正交时春麦或者冬麦做母本对胚的形成和成苗率没有影响;胚培养最佳时间为13~18 d;组培苗移入大田后成活率极低,仅为5.20%;杂交F1代成活单株有效分蘖数平均为263个。【结论】 普通小麦×粗山羊草杂交后授粉2~3次,并于13~18 d进行胚培养,可以提高杂交成功率。
[关键词]小麦;粗山羊草;正反杂交;幼胚拯救
我国虽然是小麦生产大国,但国内优质的小麦品种较少,选育优质、抗病、高产的小麦新品种一直是育种工作者面临的紧迫任务。在实际育种工作中,优异基因的短缺成为制约小麦育种的限制因子之一。
粗山羊草(AegilopstauschiiCosson,2n=2x=14,DD),又称节节麦,为禾本科小麦族山羊草属粗山羊草种,粗山羊草为二倍体,是小麦D染色体组的提供者,也是许多含有D染色体组多倍体的亲本。粗山羊草中蕴涵大量可以改良小麦抗逆性、抗病性、抗虫性、抗寒性和提高小麦品质的基因[1],可丰富小麦种质资源,拓宽小麦育种途径。
小麦与粗山羊草远缘杂交,主要有两种途径:一是小麦与粗山羊草直接杂交,经过幼胚拯救和染色体加倍获得稳定的材料;二是用四倍体小麦与粗山羊草杂交,再经过染色体加倍,获得能够稳定遗传的材料。Olson等[2]用直接杂交、回交的方法,将粗山羊草的2个新的抗小麦秆锈病基因导入普通小麦中。Wang等[3]用圆锥小麦与粗山羊草杂交,合成的2个人工六倍体都具有抗小麦黑森瘿蚊基因,并证明这2个抗虫基因都来自于粗山羊草。Laikova等[4]用一个春面包小麦与提莫菲维×粗山羊草合成的六倍体杂交,育成小麦新品种,此品种小麦可抗叶锈病、茎锈病、白粉病和散黑穗病,并具备良好的高谷物和面包制作品质。川麦38、川麦42、川麦43、川麦47都是由四川省农科院作物所以硬粒小麦/节节麦人工合成种为母本、栽培小麦为父本杂交,并通过杂交、连续回交选育而成的高产、高抗条锈病的优质小麦新品种[5-10],已经取得了非常可观的经济效益。
本研究以小麦矮败不育系(RH/CS)、中国春、辽春10号3个春性小麦及济南17和济麦20 2个冬性小麦为供试小麦材料,同时选Y122、Y212、Y215 3个粗山羊草材料,先以小麦为母本,粗山羊草为父本构建正交组合,并在去雄后1~5 d连续授粉,观察授粉次数对授粉率的影响,再以粗山羊草为母本,普通小麦为父本构建反交组合,观察各组合胚、胚乳的形成情况;对正交组合进行幼胚拯救后,统计各组合的成胚率和成苗率;并将幼苗移至大田中,观察F1代的田间表现,以期为提高小麦远缘杂交成功率提供参考。
1材料与方法
1.1供试材料
选取小麦矮败不育系(RH/CS)、中国春、辽春10号、济南17和济麦20等5个普通小麦品种(系),Y212、Y215和Y122粗山羊草3份材料,材料来源及基本信息见表1。将所有材料以穴播方式播种,行距0.5 m,株距0.1 m。为延长授粉时间,粗山羊草以5 d为间隔时间错期播种;3~4叶期定苗,每穴1株,每株只留穗中部小花去雄,套袋隔离。

表 1 供试材料及来源
1.2杂交及幼胚拯救
以普通小麦为母本,粗山羊草为父本分别杂交,构建15个正交组合,在去雄后1~5 d连续授粉,观察重复授粉对授粉率的影响。同时以粗山羊草为母本,普通小麦为父本,构建3个反交组合。
将已经消毒的幼胚,放入一步成苗N6培养基上,于恒温光照培养箱(25 ℃、12 h/d光照)中培养,适时转移到壮苗培养基(4 ℃、12 h/d光照)上继续培养,并视情况再转移3~4次,第二年春季移栽到大田。
1.3统计项目及方法
每个杂交组合授粉后3~30 d观察子房膨大情况,解剖子房,观察并统计形成的胚和胚乳,统计正常授粉及1~5次重复授粉后的授粉率,计算成胚率和成苗率。计算公式如下:
授粉率=(子房膨大数/授粉小花数)×100%;
成胚率=(获得胚的个数/授粉小花数)×100%;
成苗率=(最终获得的组培苗数/胚拯救数)×100%。
1.4数据处理
数据统计、分析与处理采用EXCEL2003和SAS9.1软件完成。
2结果与分析
2.1普通小麦与粗山羊草的正反杂交结果
从表2可见,以普通小麦为母本,以粗山羊草为父本授粉率非常低,共杂交20 422朵小花,子房膨胀小穗只有1 738个,平均授粉率仅为8.51%;而以粗山羊草为母本,以普通小麦为父本授粉的成功率非常高,仅做了1 388朵杂交,就有976朵小花子房出现膨大,平均授粉率高达70.32%。
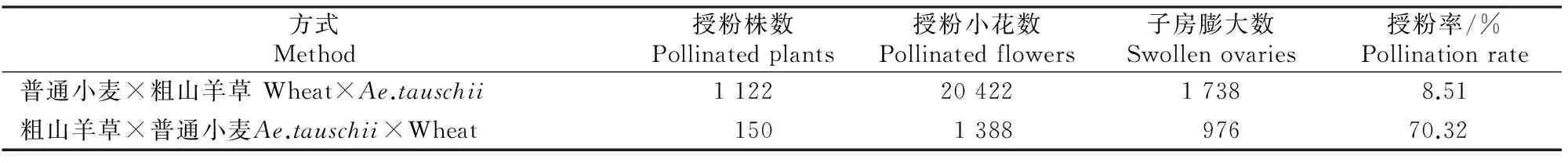
表 2 普通小麦与粗山羊草正反交授粉率统计结果
2.2重复授粉对普通小麦×粗山羊草授粉率的影响
在正交试验中,进行了1~5次重复授粉试验,从表3可见,重复授粉可以提高普通小麦×粗山羊草的授粉成功率,重复授粉4次的授粉率达到峰值,重复授粉5次则略有下降,但下降幅度很小,与重复授粉3次基本相同;重复授粉4次的授粉率比重复2次和1次授粉分别提高6.58%和37.63%。为了比较授粉次数之间的差异,随机选取不同授粉次数杂交小花各100朵,重复3次,进行方差分析,结果见表3和表4。从表3和表4可以看出,不同授粉次数之间差异极显著,其中有重复授粉杂交组合的授粉率极显著高于只进行1次授粉的杂交,但2~5次重复授粉之间的授粉率差异不显著。
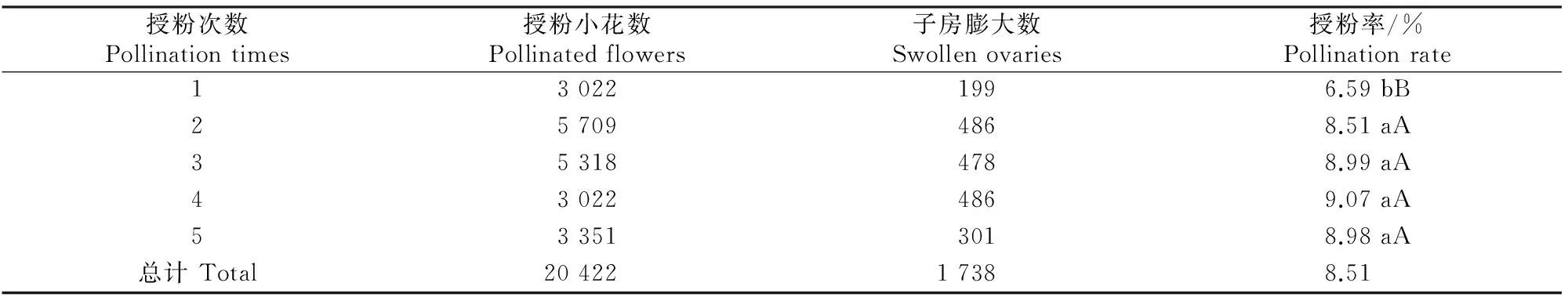
表 3 重复授粉对普通小麦×粗山羊草授粉率的影响
注:小写字母代表在α=0.05水平差异显著;大写字母代表在α=0.01水平差异极显著。
Note:Lowercase letters denote significant level ofα=0.05 while capital letters denote extremely significant level ofα=0.01.
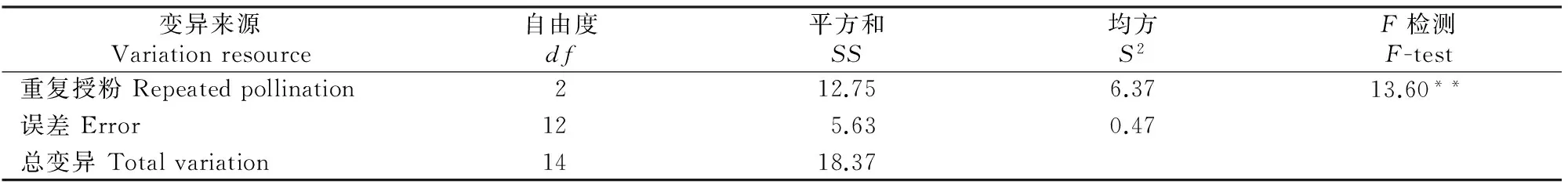
表 4 重复授粉对普通小麦×粗山羊草授粉率影响的方差分析
注:**表示在α=0.01水平差异极显著。
Note:**denotes extremely significant level ofα=0.01.
2.3普通小麦与粗山羊草正反交的成胚率情况
观察普通小麦与粗山羊草的正反交受精结果(表5)发现,无论是正交还是反交,0~8 d均未见胚和胚乳形成,子房没有膨大;9 d以后,子房逐渐膨大,解剖子房发现:膨大的子房是因胚的形成并膨大而成,胚乳一直没有形成,只有一汪清水;23 d以后,胚逐渐干瘪,失去活性。

表 5 普通小麦与粗山羊草杂交F1代胚和胚乳的形成状况
从表6可见,以春性、冬性小麦为母本与粗山羊草杂交,子房(胚)膨大的比例不同,成胚率最高的是济麦20×Y215组合,为10.27%;最低的是济麦20×Y122组合,为7.38%;即成胚率最高和最低都来自于冬麦济麦20组合。以春麦为母本的杂交组合的平均成胚率(8.34%)略低于冬麦组合(8.72%),但二者差异不显著。在胚拯救形成幼苗时,春麦杂交组合和冬麦组合也没有显著差异,成苗率最低的是冬麦济麦20×Y122杂交组合,为7.27%,最高的为春麦矮败×Y122杂交组合,为9.80%(表6)。因以粗山羊草为母本远缘杂交的F1代细胞质来自于粗山羊草,在生产上应用价值较低,所以未做幼胚拯救。
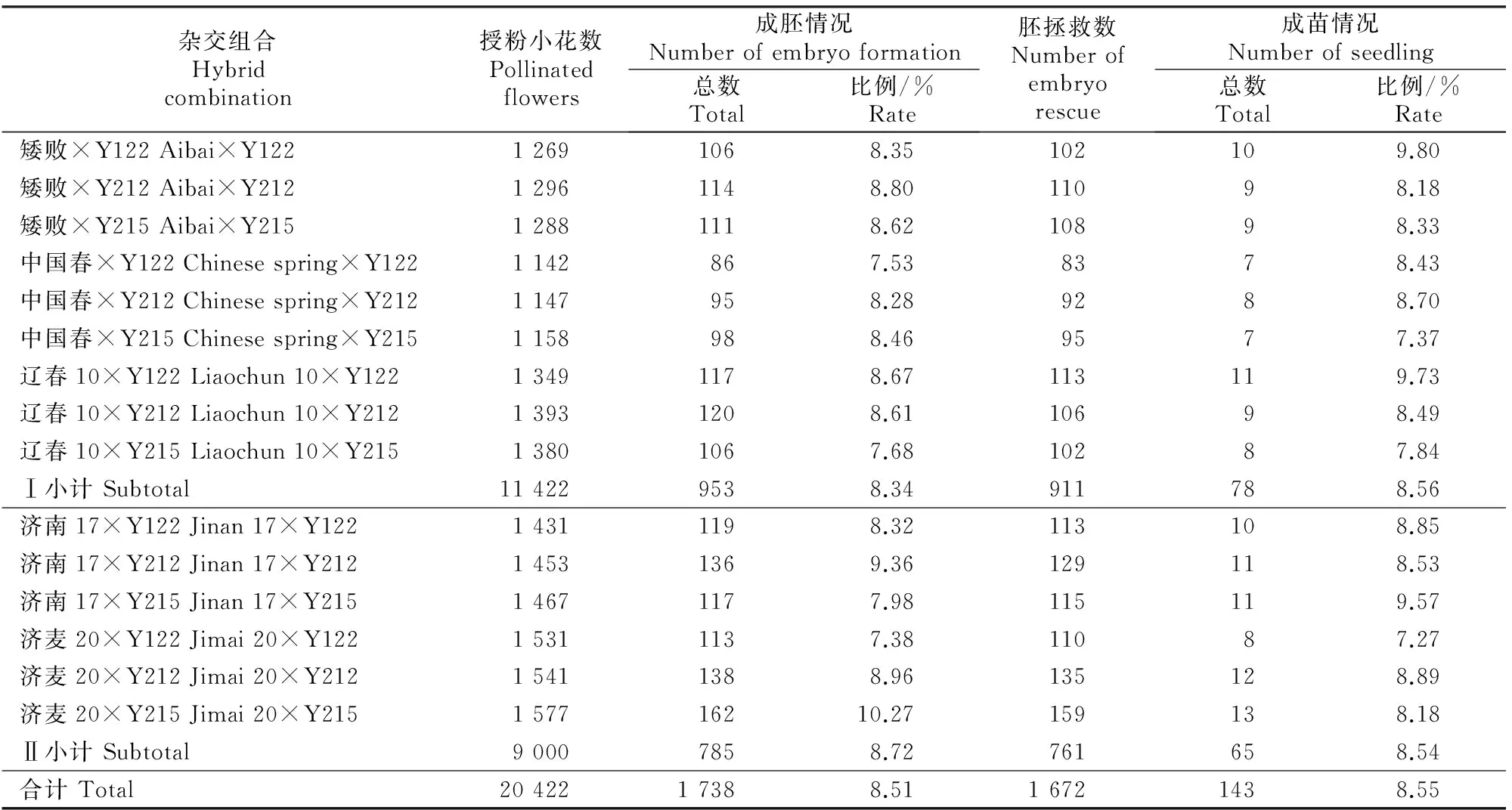
表 6 春性、冬性小麦与粗山羊草杂交的成胚率和成苗率比较
注:Ⅰ为春性小麦与粗山羊草杂交;Ⅱ为冬性小麦与粗山羊草杂交。
Note:Ⅰis cross combination of spring wheat andAe.tauschii,Ⅱis cross combination of winter wheat andAe.tauschii.
2.4普通小麦×粗山羊草杂种F1代的田间表现
幼胚拯救苗在实验室转移过程中,因消毒及其他原因,最初胚拯救成功的有143株苗,存活97株,栽入大田后,最后只存活5株,分别是矮败×Y215组合2株、中国春×Y215组合1株和济南17×Y212组合2株。
杂种F1代的农艺性状表现介于普通小麦和粗山羊草之间,外部形态接近于普通小麦。分蘖力强,单株平均分蘖为297个,有效分蘖263个(图1)。自交不结实,与普通小麦回交后,可以结实,但结实率非常低。

图 1矮败(RH/CS)×粗山羊草Y215杂交F1代的田间表现
Fig.1F1of Aibai(RH/CS)×Ae.tauschiiY215
3结论与讨论
本研究表明,普通小麦×粗山羊草远缘杂交,只有通过幼胚拯救才能获得杂交后代,与袁汉民等[11]“不用胚拯救即可获得普通小麦与山羊草组柱穗山羊草(Ae.cylindricaHost,CD,2n=2x=14)远缘杂交后代种子”的研究结果相反。其原因是因为本研究中所有的杂交组合都不能形成胚乳,而胚没有得到胚乳的滋养很快就会干瘪,失去生命力。普通小麦与粗山羊草杂交授粉率很低,多次授粉的授粉率明显高于一次授粉,二者差异极显著;但2~5次授粉间的差异不显著,因此从经济成本角度考虑,以重复授粉2~3次为宜。以粗山羊草为母本的杂交组合,授粉率非常高,高达70.32%,也需要幼胚拯救;授粉1 388朵小花后,有976个子房膨大并形成胚,但因其应用价值较低,未做胚拯救,因此未获得种子。反交授粉率显著高于正交的原因可能是:在正交组合中,雌配子体为三倍体(ABD),其花柱较长、胚囊较大,而雄配子体(花粉)为一倍体(D),生活力相对较弱,多数花粉的花粉管难以伸长到胚囊中与卵子受精结合为合子,进而发育成胚;相反,在反交组合中,雌配子体(胚囊)为一倍体(D),其花柱较短、胚囊较小,而雄配子体则为三倍体(ABD),生活力强,其花粉管极易伸长到胚囊中与卵子结合形成胚,因此可能导致反交组合的成胚率比正交组合高得多。另一方面,也可能是在正交组合中,三倍体胚囊(ABD)对一倍体雄配子体(D)的容忍性要比反交组合中一倍体胚囊(D)对三倍体雄配子体(ABD)的容忍性大,可能导致正交组合的卵细胞与精细胞结合难,但胚的发育较好、质量较高,更易做幼胚拯救。
通过正交和反交种子形成过程可见,由于均是六倍体与二倍体的远缘杂交,正交和反交都不能形成初生胚乳核,也就不能进一步发育成胚乳。胚由于得不到胚乳的滋养,因此经历了“胚形成→胚膨大→胚干瘪→失去生命力”的过程。授粉后9~22 d是胚有活力的时间,超过22 d胚逐渐干瘪,胚培养的最佳时间是13~18 d,生命力强,获得组培苗的可能性比较大。
以普通小麦为母本、粗山羊草为父本的不同组合,其杂交授粉率不同,变幅为7.38%~10.27%;春性小麦的平均授粉率(8.34%)低于冬性小麦(8.72%),但二者之间差异不显著。通过胚培养获得的普通小麦×粗山羊草F1代可以形成苗,但成苗率很低,平均为8.55%;春性小麦的成苗率为8.56%,冬性小麦的成苗率为8.54%,差异不明显。
F1代组培苗转移培养及移栽大田过程中成活率非常低,幼胚拯救获得143株组培苗,经继代培养得到97株苗,成活率为67.80%;移入大田后,只成活5株,成活率仅为5.20%。继代培养过程中组培苗大量死亡的原因可能是:F1细胞核染色体组为ABDD,进行减数分裂产生配子时会发生核染色体数目混乱,难以产生可育配子,因而通常不结实;组培苗A、B属于单倍性,植株羸弱,生命力不强;消毒不严格出现病菌感染等。移栽大田后的死亡原因可能是:(1)组培苗一直在恒温培养箱中的培养基上生长,比较瘦弱,移栽到大田后外部环境变得非常恶劣,幼苗不适应环境而死亡;(2)组培苗是在无菌条件下培养,一旦移栽到大田中,空气和土壤中的有害病原菌大量侵入,导致死亡等。
F1代在大田中的形态表现接近小麦,但分蘖率极高,平均分蘖297个,有效分蘖263个,分蘖数较多,可能与粗山羊草F1代基因导入有关,也可能与周围没有其他植物竞争有关。F1代自交不结实,通过与普通小麦回交可以获得种子,但结实率很低。
另有研究表明,小麦的kr基因能抑制异源花粉的亲和性[12],且对不同的远缘杂交影响不同,如对小麦与黑麦杂交的抑制作用非常明显,使其杂交亲和性降低,却对小麦与玉米杂交没有影响[13]。但也有研究指出,kr基因不能影响远缘杂交的亲和性[14-15]。本研究中“中国春”品种有隐性的kr1kr1kr2kr2kr3kr3基因,杂交结果显示该品种授粉率比其他杂交组合无明显增加,暗示可能kr基因对小麦与粗山羊草远缘杂交的亲和性没有影响。
[参考文献]
[1]Jia J,Zhao S,Kong X,et al.Aegilopstauschiidraft genome sequence reveals a gene repertoire for wheat adaptation [J].Nature,2013,496:91-95.
[2]Olson E L,Rouse M N,Pumphrey M O,et al.Introgression of stem rust resistance genes SrTA10187 and SrTA10171 fromAegilopstauschiito wheat [J].Theor Appl Genet,2013,126:2477-2484.
[3]Wang T,Xu S S,Harris M O,et al.Genetic characterization and molecular mapping of Hessian fly resistance genes derived fromAegilopstauschiiin synthetic wheat [J].Theor Appl Genet,2006,113:611-618.
[4]Laikova L I,Belan I A,Badaeva E D,et al.Development and study of spring bread wheat variety Pamyati Maystrenko with introgression of genetic material from synthetic hexaploidTriticumtimopheeviiZhuk.×AegilopstauschiiCoss [J].Genetika,2013,49(1):103-112.
[5]张颙,杨武云,郑有良,等.源于硬粒小麦:节节麦人工合成种的小麦新品种川麦38抗条锈性及遗传分析 [J].西南农业学报,2006,19(1):14-18.
Zhang Y,Yang W Y,Zheng Y L,et al.Analysis of agronomic characters of new wheat variety Chuanmai 38 derived from synthetics (Triticumdurum×Aegilopstauschii) [J].Southwest China Journal of Agricultural Sciences,2006,19(1):14-18.(in Chinese)
[6]张颙,杨武云,彭云良,等.小麦新品种川麦42抗条锈病性遗传分析 [J].植物保护学报,2006,33(3):287-290.
Zhang Y,Yang W Y,Peng Y L,et al.Inheritance of resistance for Chinese wheat stripe rust races in a new common wheat variety Chuanmai 42 derived from synthetics betweenTriticumdurum×Aegilopstauschii[J].Acta Phytophylacica Sinica,2006,33(3):287-290.(in Chinese)
[7]谭禺,杨武云.人工合成小麦衍生品种川麦43 [J].四川农业科技,2008(10):23.
Tan Y,Yang W Y.Synthetic derivative Chuanmai 43 [J].Science and Technology of Sichuan Agriculture,2008(10):23.(in Chinese)
[8]李俊,魏会廷,胡晓蓉,等.人工合成小麦衍生品种川麦47的抗条锈病SSR分子标记定位 [J].农业生物技术学报,2007,15(2):318-322.
Li J,Wei H T,Hu X R,et al.Molecular mapping of stripe rust resistance gene in synthetic derivative Chuanmai 47 [J].Journal of Agricultural Biotechnology,2007,15(2):318-322.(in Chinese)
[9]张颙,杨武云,邹裕春.源于CIMMYT人工合成种的小麦新品种川麦42的选育与研究 [J].西南农业学报,2007,20(2):199-202.
Zhang Y,Yang W Y,Zou Y C.Characterisation of the new wheat cultivar Chuanmai 42 derived from CIMMYT hexaploid wheat [J].Southwest China Journal of Agricultural Sciences,2007,20(2):199-202.(in Chinese)
[10]张颙,杨武云,谭禺.高产、高抗条锈病小麦新品种川麦47 [J].四川农业科技,2008(11):20.
Zhang Y,Yang W Y,Tan Y.High-yield,high resistance to stripe rust new wheat cultivar Chuanmai 47 [J].Science and Technology of Sichuan Agriculture,2008(11):20.(in Chinese)
[11]袁汉民,王小亮,陈东升,等.普通小麦远缘杂交F1代表现型研究 [J].植物遗传资源学报,2005,6(4):377-380.
Yuan H M,Wang X L,Chen D S,et al.Study on F1phenotype of wheat crosses involving wild relatives and triticale [J].Journal of Plant Genetic Resources,2005,6(4):377-380.(in Chinese)
[12]Manickavelu A,Koba T,Mishina K,et al.Molecular characterization of crossability genekrfor intergeneric hybridization inTriticumaestivum(Poaceae:Triticeae) [J].Plant Systematics and Evolution,2009b,278(12):125-131.
[13]Torres L E,Bima P,Maich R,et al.Production of haploid pla-nts from ten hybrids of bread wheat(TriticumaestivumL.)through wide hybridization with maize(ZeamaysL.) [J].Agriscientia,2010,2:79-85.
[14]Boyes J W,Thompson W P.The development of the endospe-rm and embryo in reciprocal interspecific crosses in Cereal [J].J Genet,1937,34:203-207.
[15]Julian B,Thomas P J,Kaltsikes R,et al.Relation between wh-eat-rye cross ability and seed set of common wheat after pollination with other species in Hordeae [J].Euphytica,1981,30:121-127.
Reciprocal crosses ofTriticumaestivumandAegilopstauschii
ZHANG Hai-quan,YANG Hong,LANG Jie
(CollegeofBiologyEngineering,EconomicsandBusinessofHebeiUniversity,Shijiazhuang,Hebei050061,China)
Abstract:【Objective】 This study investigated embryo rate,seedling rate and F1 performance in field of reciprocal crosses of common wheat and Aegilops tauschii to provide reference for improving hybridization success rate.【Method】 Three spring wheat varieties (the wheat dwarf male sterile lines (RH/CS),Chinese spring,and Liaochun 10) and two winter wheat varieties (Jinan 17 and Jimai 20) were selected to cross with three tauschii materials (Y122,Y212,and Y215).First,common wheat varieties were used as female parent and Aegilops tauschii varieties were used as male parent to construct orthogonal combinations.Within the 1-5 consecutive days of after emasculation,pollination was conducted and the effects of pollination times on pollination rate were analyzed.Then,common wheat varieties were used as male parent and Aegilops tauschii varieties were used as female parent.The formation of embryo and endosperm was analyzed.After embryo rescue,the embryo rate and seedling rate were calculated and the seedlings were moved to field for observation of F1 generation performance.【Result】 Reciprocal cross combinations had much higher pollination rate (70.32%) than orthogonal combinations (8.51%).Both orthogonal and reciprocal crosses failed to form a seed,and embryo rescue was required to obtain offsprings.Repeated pollination improved the pollination rate.Female parent had no effects on embryo formation and seedling rate of orthogonal combinations.Best time for embryo culturing was 13-18 days.The survival rate of tissue culture plantlets moved to field was as low as 5.20%.The number of tillers per plant was 263.【Conclusion】 Hybridization of common wheat and Aegilops tauschii could be improved by pollination for 2-3 times and culturing embryo for 13-18 d.
Key words:wheat;Aegilops tauschii;reciprocal crosses;embryo rescue
DOI:网络出版时间:2016-03-1408:4510.13207/j.cnki.jnwafu.2016.04.005
[收稿日期]2014-08-21
[基金项目]河北省科学技术研究与发展指导计划项目“微卫星标记小麦野生资源抗病基因及抗病基因的利用”(05225510)
[作者简介]张海泉(1964-),男,辽宁锦州人,教授,博士,主要从事分子生物学研究。E-mail:hqzhang188@126.com
[中图分类号]S512.1;S332.2
[文献标志码]A
[文章编号]1671-9387(2016)04-0033-06
网络出版地址:http://www.cnki.net/kcms/detail/61.1390.S.20160314.0845.010.html
猜你喜欢
今日农业(2022年16期)2022-11-09
今日农业(2021年12期)2021-11-28
今日农业(2021年8期)2021-11-28
今日农业(2021年8期)2021-11-28
金桥(2021年10期)2021-11-05
今日农业(2021年13期)2021-08-14
河北农机(2020年10期)2020-12-14
今日农业(2020年13期)2020-08-24
作文小学中年级(2020年4期)2020-06-11
今日农业(2019年11期)2019-08-13
