
基于MaxEnt模型的可可潜在适宜分布研究
2016-05-30 07:04:05秦晓威郝朝运李付鹏谭乐和宋应辉赖剑雄
热带作物学报 2016年5期
秦晓威 郝朝运 李付鹏 谭乐和 宋应辉 赖剑雄

摘 要 根据野外采样和文献查阅,系统整理了可可的地理分布记录,并利用MaxEnt生态位模型和ArcGIS软件对可可的潜在适宜分布范围进行预测。结果表明:北美洲南部、中南美洲北部、非洲西部、亚洲东南部以及太平洋美拉尼西亚群岛地区均是可可的潜在适宜分布区域。其中,中国海南、台湾南部、云南西双版纳、广东雷州半岛也属于可可的适生范围。经ROC(Receiver operating characteristic)曲线分析法验证,MaxEnt模型的AUC(Area under curve)值为0.977,表明预测结果具有较高的可信度。各环境变量重要性的Jacknife检验表明,极端最低温度、年降雨量、年温度变化范围、最暖季降雨量对可可的潜在分布影响最大。
关键词 可可;MaxEnt模型;适宜分布区;ArcGIS地理信息系统
中图分类号 S571.3 文献标识码 A
Abstract Theobroma cacao L, the chocolate tree, is an important tropical crop. In order to understand the possible distribution areas of T. cacao L., a prediction of potential geographical distribution of T. cacao L. was conducted by the maximum entropy(MaxEnt)species distribution model combined with geographic information system(ArcGIS). The results showed that the potential distribution of T. cacao L. mainly concentrated in Southern North America, Northern South America, Western Africa, South-Eastern Asia, and Melanesian Islands in the South Pacific. The suitable areas for T. cacao L. cover only some areas in Hainan, Southern Taiwan, Xishuan Banna in Yunnan, Leizhou Peninsula in Guangdong in China. The checked result of receiver operating characteristic(ROC)curve analysis showed that the area under curve(AUC)value of MaxEnt model reached 0.977, indicating that the predicted results was fairly satisfactory. A Jackknife test in MaxEnt showed that minimum temperature of coldest month, annual precipitation, temperature annual range, and precipitation of warmest quarter were the most important environmental variable affecting the distribution of the cocoa species.
Key words Theobroma cacao L.; MaxEnt ecological niche model; Potential geographic distribution; Geographic information system ArcGIS
doi 10.3969/j.issn.1000-2561.2016.05.027
可可(Theobroma cacao L.)为梧桐科(Sterculiaceas)可可属(Theobroma)多年生热带经济作物。据Henderson等[1]考古证实,可可约有3 000多年的食用历史。目前,世界可可收获面积达1 000万hm2,产量约460万t,贸易额达139亿美元[2]。其中,非洲占总产量的57%、北美洲16%、南美洲12%和亚洲15%。中国可可引种历史相对较短,1922年首次由印度尼西亚和爪哇地区引入台湾嘉义、高雄等地试种。20世纪50年代又先后从越南、泰国、马来西亚、巴布新几内亚、厄瓜多尔、科特迪瓦、斯里兰卡、哥斯达尼加等国家引进可可种质,在海南万宁等地试种成功。目前,主要栽培于海南和台湾地区,云南、广东、福建等地也有零星分布[3]。可可在世界热区的广泛传播,已成为发展中国家热区农业经济收入的特殊组成,对于发展热带经济,提高边远地区农民收入和生活水平具有重要意义。
可可的引种传播与推广种植在很大程度上依赖于农业气候资源状况[4-8]。可可为中南美洲亚马逊河流域热带雨林的特有物种,地域分布狭窄。16世纪中期以来,由美洲大陆陆续传入亚洲的东南部和非洲的西部。据国际可可组织统计(ICCO),已有60多个国家和地区种植可可,其种植区域将由美洲向非洲、亚洲东南部及其太平洋岛屿进一步扩大。目前,可可收获种植面积已由1960年的400万hm2增至2014年的1 000万hm2[2]。可可在世界热区的传播佐证了可可对不同气候区域的适应性。然而,ICCO统计表明,2013~2014年度可可产量比预期减产7.5万t。La··derach等[9]研究认为,可可供需赤字与科特迪瓦、加纳和印度尼西亚等可可主产区极端气候频发密切相关[10-12]。而温度升高、降雨减少等气候变化导致西非可可种植区由干燥的东部向湿润的西南部气候区转移[13]。此外,Wanger[14]、Larson[15]等研究认为环境条件制约着可可作物的空间分布格局与产量波动。气候条件成为限制可可产业提升的重要因素[16-18]。但诸多研究以局域尺度试验为主,其研究的空间尺度和气象因子选择存在很大差异,大多影响因子的选择以主观经验为主,缺乏客观定量的评价[18]。因此,如何合理广泛的利用丰富多样的气候资源,评估适宜引种与种植的气候区域,以便获取更高的产量,成为可可精细化气候区划的热点问题。
近年来,基于植物物种地理分布位点与气候环境因子之间的紧密联系为探索目标物种潜在地理分布格局及其气候适宜性区划提供了技术途径[9,19-21]。国内外的众多学者利用机理模型、回归模型和生态位模型结合GIS、GPS、RS技术对目标物种进行气候适宜性评价[22-24]。最大熵模型(MaxEnt)是一种根据已知物种分布信息预测该物种潜在分布区域的生态模型,广泛用于生物地理、生物入侵、园林引种以及农作物种植区区划等领域[19,25-26]。该模型采取一种可以最大程度满足约束未知分布信息的环境变量的运算方法,即“最大熵原则”,可以更多地保留物种已有分布数据的有用信息。与其他方法相比,MaxEnt模型所具备的优势使其预测表现始终是最好的[27]。因此,本研究采用MaxEnt模型和气候环境图层建模分析可可的潜在种植区分布和主要影响因子,以期为可可作物种质资源保护及种植生产提供科学策略和依据。
1 研究方法
1.1 可可地理分布位点数据采集
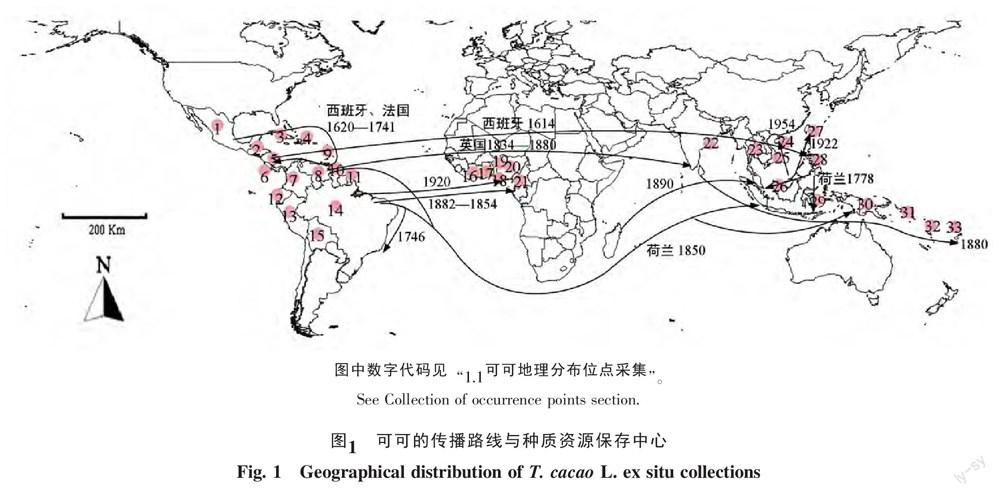
查阅标本、种质资源保存中心和文献确定可可现已知的地理分布位点。腊叶标本信息主要通过密苏里植物园(http://www.mobot.org)、美国国家植物标本馆(http://www.nmnh.si.edu/botany)、亚马逊哥伦比亚植物标本馆(http://www.sinchi.org.co)、哥斯达黎加国家生物多样性机构(http://www.inbio.ac.cr)、墨西哥生物多样性全国委员会(http://www.biodiversidad.gob.mx)、菲尔德自然历史博物馆(http://www.fieldmuseum.org)、加纳大学(http://www.ug.edu.gh)、iNaturalist(http://www.inaturalist.org)、Discover Life(http://www.discoverlife.org)、中国数字植物标本馆(http://www.cvh.org.cn)、中国物种信息系统(http://www.chinabiodiversity.com)、华南植物园标本馆(http://www.scib.ac.cn)、西双版纳热带植物园(http://www.xtbg.ac.cn)以及中国热带农业科学院香料饮料研究所(http://xys.xlrdzwy.com)等标本信息平台查询获得。
活体保存信息主要通过可可种质资源保存中心获得(图1),包括墨西哥(1,INIFAP)、洪都拉斯(2,FHIA)、古巴(3,EIC/ECICC)、多米尼加(4,IDIAF)、尼加拉瓜(5,UNAN)、哥斯达黎加(6,CATIE)、哥伦比亚(7,CORPOICA)、委内瑞拉(8,INIA)、特立尼达和多巴哥(9,CRU/UWI)、圭亚那(10,MHOCGA)、法属圭亚那(11,CIRAD)、厄瓜多尔(12,INIAP)、秘鲁(13,CEPICAFE-ICT-UNAS-UNSAAC)、巴西(14,CEPEC-SUEPA-SUERO-ICA)和玻利维亚(15,EI Ceibo Cooperative)等15个拉丁美洲国家,科特迪瓦(16,CNRA)、加纳(17,CRIG)、多哥(18,CRAF)、贝宁(19,CRA/SB)、尼日利亚(20,CRIN)和喀麦隆(21,IRAD)等6个西非国家以及印度(22,CPCRI)、泰国(23,CHRC)、中国(24,CATAS;27,CAS)、越南(25,Nong Lam University)、马来西亚(26,MCB)、菲律宾(28,USMARC/PICRI)、印度尼西亚(29,Bah Lias-ICCRI)、巴布新几内亚(30,CCI)、所罗门群岛(31,BPCU)、瓦努阿图(32,VARTC)、斐济(33,Dobuilevu)等11个亚洲国家。去除经、纬度缺失和重复以及标本信息缺失的条目后,共得到288个有效记录地理信息位点。按MaxEnt模型要求,将可可分布记录数据的物种名和分布点经、纬度顺序录入CSA格式的Excel文件,其中东经和北纬标为正值,西经和南纬标为负值。
1.2 方法
1.2.1 环境数据采集 参考郝朝运[19]、胡秀[20]和Hijmans[28]等研究,环境数据采用WorldClim-Global Climate Data气候数据资料(http://www.worldclim.org),数据年份为1950~2000年,空间分辨率为10 km。环境变量包括了对植物生长具有重要影响的19个生物气候变量(Bio1~Bio19),较适用于植物的潜在适宜区预测。分别为:
反映平均温度及其变异幅度的指标:年均大气温度(Bio1)、温度季节性变化系数(Bio4)。
反映极端温度影响的指标:极端最高温度(Bio5)、极端最低温度(Bio6)、最热季平均温度(Bio10)、最冷季平均温度(Bio11)。
反映降雨量及其季节性分布的指标:年降雨量(Bio12)、降雨量季节性变化(Bio15)。
反映极端水分条件的指标:最湿月降雨量(Bio13)、最干月降雨量(Bio14)、最干季降雨量(Bio16)、最湿季降雨量(Bio17)。
反映水热是否同步的指标:最湿季平均温度(Bio8)、最干季平均温度(Bio9)、最暖季降雨量(Bio18)和最冷季降雨量(Bio19)。
反映温差特点的指标:平均月温度变化(Bio2)、等温性(Bio3)、年温度变化范围(Bio7)。
1.2.2 模型的建立与结果检验 本研究应用最大熵法则(Maximum entropy, MaxEnt)的神经网络模拟方法进行可可潜在适宜分布区的预测。MaxEnt 3.33软件在http://www.cs.princeton.edu/~schapire/maxent网站注册并免费下载,关于软件的介绍和详细信息可以参考Phillips[23]、Elith[27]等文献。将可可的分布数据和19个生物气候变量导入MaxEnt,参数设置为:25%的分布点作为测试集,剩余的分布点为训练集,选择Jackknife法分析影响因子,输出格式为ASCII。MaxEnt软件通过分布数据和生物气候数据建立预测模型,再根据模型进行模拟计算,得到研究区域每个象元可可的适生概率,取值范围为0~100%。图形处理:ASCII格式文件导入ArcGIS进行格式转换,并根据适宜分布概率大小进行适宜性等级分类。
2 结果与分析
2.1 可可在全球的潜在适宜分布区分析
采用ROC(Receiver operating characteristic)曲线分析法对应用MaxEnt模型预测的可可适宜分布区结果进行精度检验。结果表明,可可MaxEnt模型训练集的AUC(Area under curve)值为0.991,测试集的AUC值为0.977,显著高于随机模型的AUC 0.5值,表明预测结果具有较高的精度,即模型预测分布区与可可实际分布区的拟合度较好。
根据可可在全球的分布记录数据,使用MaxEnt和Arc-Gis结合分析可可在全球的适宜分布区域(图2)。在当前气候条件下,可可在全球的适宜分布区位于17°S~24°N、100°W~118°E的热带、亚热带之间,主要集中在北美洲南部、南美洲北部、非洲西部、亚洲东南部及太平洋群岛地区,包括北美南部的墨西哥、巴拿马、哥斯达黎加、尼加拉瓜、洪都拉斯、萨尔瓦多、特立尼达、多米尼加,南美北部的委内瑞拉、哥伦比亚、厄瓜多尔、秘鲁、玻利维亚、法属圭亚那、圭亚那,西非的加纳、科特迪瓦、尼日利亚、贝宁、多哥,亚洲东南部的中国台湾、中国海南、越南、缅甸、泰国、马来西亚、印度尼西亚、菲律宾、斯里兰卡以及太平洋美拉尼西亚群岛的巴布新几内亚、所罗门群岛、瓦努阿图群岛、斐济群岛地区。
为了细致区分各区域的适生程度并方便应用,参照可可在世界各地区引种与推广种植的描述,通过选定合适的阈值划分适生等级。根据预测发生概率p值进行设定:p<0.19为非适宜区,0.19≤p<0.40为低适宜区,0.40≤p<0.60为中适宜区,0.60≤p<0.80为高适宜区,0.80≤p<1.00为极高适宜区。
极高适宜区(0.80~1.00):主要集中在北美洲墨西哥的南部、南美洲北部的秘鲁、哥伦比亚和厄瓜多尔交界处以及玻利维亚的拉巴斯和特立尼达地区。
高适宜区(0.60~0.80):主要位于极高适宜分布区域的外缘,包括北美洲南部的洪都拉斯、萨尔瓦多、尼加拉瓜、哥斯达黎加、巴拿马、巴西亚马逊州、委内瑞拉奥里诺科河流域,西非的科特迪瓦西南部、斯里兰卡东部沿海,亚洲东南部的越南、菲律宾、马来半岛的西马来西亚、以及太平洋美拉尼西亚群岛的巴布新几内亚地区。
中适宜区(0.40~0.60):主要位于高适宜分布区域的外缘,包括南美洲东北部的法属圭亚那、圭亚那、特立尼达和多巴哥,西非的尼日利亚、贝宁和加纳,亚洲的中国海南、台湾、印度尼西亚爪哇岛。
低适宜区(0.19~0.40):主要分布于南美洲的巴西北部、西非的喀麦隆、亚洲东南部的东马来西亚、印度尼西亚苏门答腊岛、越南河内、中国广东、福建以及太平洋美拉尼西亚群岛的斐济、所罗门群岛、瓦努阿图群岛等。
非适宜区(0.00~0.19):除以上4级区域的其他地区为可可的非适宜区。
2.2 可可在中国的潜在适宜分布区分析
由图3可见,可可在中国适宜分布范围较狭窄,主要分布在海南、台湾、云南等热带北缘气候区。其中,极高适宜区和高适宜区主要在海南东部、台湾南部,主要包括海南文昌、琼海、万宁、保亭,台湾高雄以及广东雷州半岛等沿海区域。潜在适宜分布范围有所扩大,主要分布于海南澄迈、定安、屯昌、琼中、五指山、乐东、三亚、东方、儋州,云南西双版纳、广东的湛江,福建的厦门和台湾的台南、台东、嘉义等地区。
2.3 可可的适宜性分布与环境变量的关系
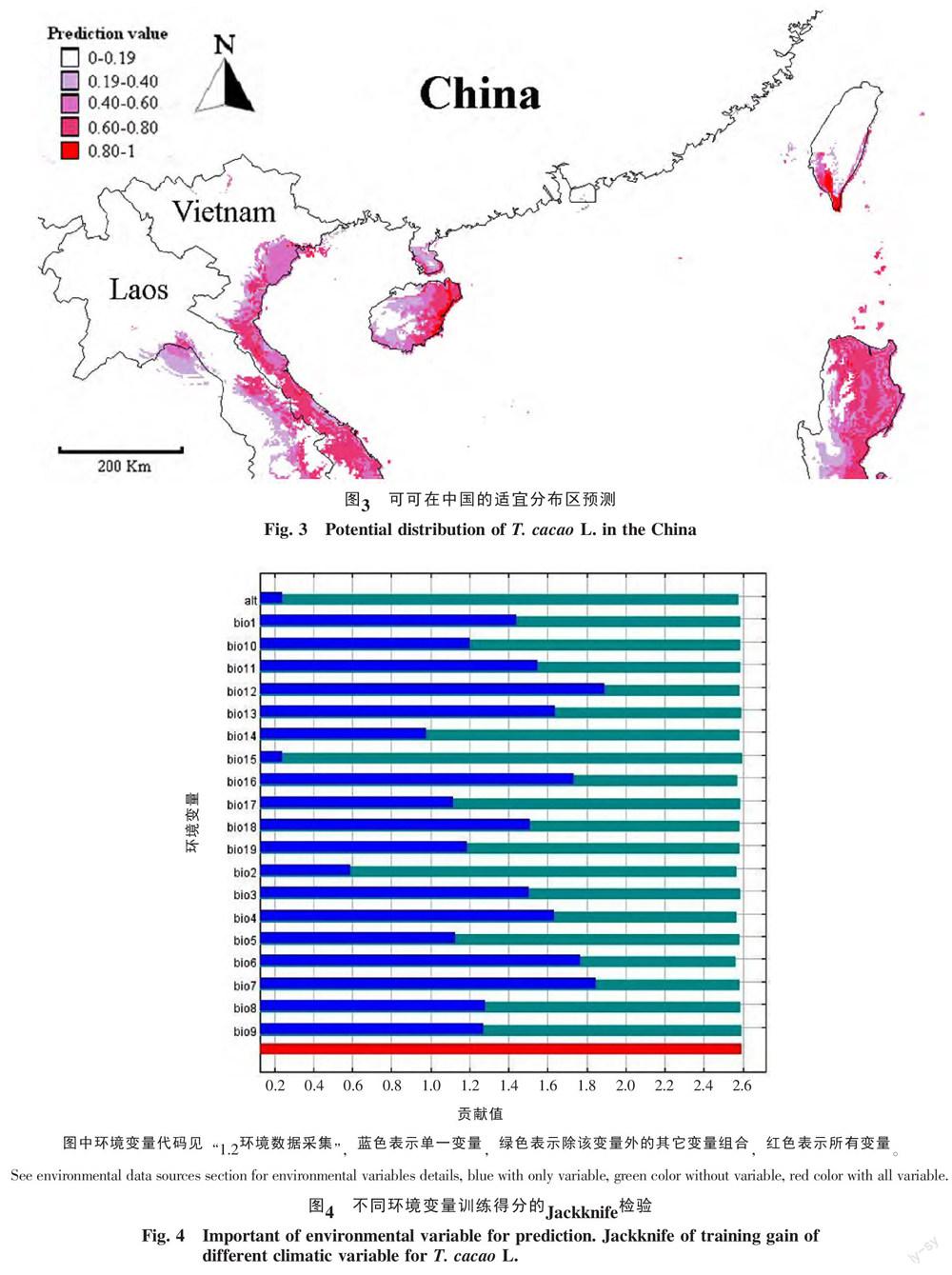
用刀切法(Jackknife Test)检测生物气候变量对于分布增益的贡献,结果显示(图4),年降雨量(bio12)、年温度变化范围(bio7)、极端最低温度(Bio6)、最干季降雨量(Bio16)和温度季节性变化系数(Bio4)等共10个环境因子在Jacknife检验中获得了较高得分(>1.4),说明这些因子在模拟时起到了重要作用;其他环境变量,如海拔、平均月温度变化(Bio2)、等温性(Bio3)、温度季节性变化系数(Bio4)、极端最高温度(Bio5)等的得分较低,表明这些因子起到了次要作用。由于各环境因子之间往往具有很强的相关性,很容易导致过拟合,本研究利用MaxEnt软件对因子间的相关性进行分析,以移除对模拟具有相似影响的因子。结果表明,共有4个环境因子对模拟的影响贡献率大于5%,由高到低依次为极端最低温度(Bio6,34.4%)、年降雨量(bio12,24.2%)、年温度变化范围(bio7,14.8%)和最暖季降雨量(Bio18,6.9%),其累积贡献率达80%,是最主要的影响因子。
绘制极端最低温度、年降雨量、年温度变化范围和最暖季降雨量与分布值之间的响应曲线,以分布值0.2为阈值(用训练数集数据中随机选取10%来计算的分布阈值),然后划出适合可可分布的生物气候变量范围。结果显示,极端最低温度的适宜值为12.5~22.3 ℃,最适宜值为19.8 ℃,在12.5~19.8 ℃时,随温度的升高分布值增大,在19.8~22.3 ℃时,随温度升高分布值降低(图5-a);年降雨量的适宜值为>1 500 mm,最适宜值为3 210 mm(图5-b);年温度变化范围的适宜值为9.2~18.5 ℃,最适宜值为11.1 ℃,在9.2~11.2 ℃时,随温度的升高分布值增大,在11.2~18.5 ℃时,随温度的升高分布值降低(图5-c);最暖季降雨量的适宜值为>400 mm,最适宜值为750 mm(图5-d)。
3 讨论
基于已有的研究成果,本研究通过检索可可腊叶标本记载信息以及可可种质资源活体保存资料,利用基于最大熵理论的MaxEnt模型并结合地理信息系统,综合分析了可可的潜在适宜性分布及主导环境因子阈值。结果表明,北美洲南部、中南美洲北部、非洲西部、亚洲东南部以及太平洋美拉尼西亚群岛地区均是可可的潜在适宜分布区域。该结论与Bartley等[29]描述的泛性结论比较吻合,也验证了MaxEnt模型预测可可适宜分布区的可靠性。此外,本研究发现可可的适宜分布区为15° S~24° N,比Monteiro等[30]记载的纬度分布范围15° S~18° N广。该研究结果进一佐证了16世纪中期以来,可可在世界范围的引种与传播呈现“北扩南移”的发展趋势。同时,ROC曲线分析法验证结果表明,MaxEnt模型的AUC值为0.977(非常接近1),预测结果具有较高的精度。
根据Jackknife检验,一些降雨和温度相关的环境变量被认为对可可的潜在适宜分布有重要影响。基于可可已知分布位点信息,结合最大熵模型,根据气候因子的贡献程度确定了影响可可潜在分布的4个主导气候因子:极端最低温度、年降雨量、年温度变化范围、最暖季降雨量,其累积贡献率达80%。各主导因子的阈值:极端最低温度12.5~22.3 ℃,年降雨量>1 500 mm,年温度变化范围9.2~18.5 ℃,最暖季降雨量>400 mm。这在一定程度上证实了降雨量和温度是影响可可生产的主要气候因素[9,11-13]。根据可可的存在概率,本研究将可可预测图的适生性划分为5个等级。在预测图中,区划等级在0.4以上的区域可以进行可可的商品生产,为种植的适宜区。例如,以北美洲的洪都拉斯、南美洲的巴西、委内瑞拉、西非的科特迪瓦、亚洲东南部的马来西亚、菲律宾、太平洋美拉尼西亚群岛的巴布新几内亚为代表的高适宜区,构成了可可生产的核心区,约占世界可可收获面的37%和总产量的45%[2];同时,以南美洲的法属圭亚那、圭亚那、特立尼达和多巴哥、西非的尼日利亚、加纳、亚洲的印度尼西亚、中国海南、中国台湾为代表的中适宜区,为可可生产的辐射区,约占世界可可收获面积的39%和总产量的46%。而以北美洲的墨西哥、南美洲的秘鲁、哥伦比亚、厄瓜多尔等为代表的极高适宜区,Dias[31]的研究结果认为,这些区域是可可的起源中心。此外,本研究找出了更多适合可可引种与推广种植的区域,如亚洲东南部的东马来西亚、印度尼西亚苏门答腊岛、越南河内、中国广东、福建以及太平洋美拉尼西亚群岛的斐济、所罗门群岛、瓦努阿图群岛等地区,为新兴的可可引种试种区域。该划分结果基本符合当前可可的种植现状,可为可可的生产布局及制定可可应对气候变化对策提供参考。
需要指出的是,可可的潜在适宜性分布不仅取决于气候条件,还取决于社会经济结构、生产技术水平等因素的综合影响,同时还受到包括土壤类型、地理特征、遗传类型、人为活动、历史原因等因素的影响[32-33]。因此,在实际生产活动中,可可引种试种、种植还需要综合考虑各种因素的作用,特别是要考虑产量因素及其经济要素对可可种植的影响。此外,由于该模型软件目前还不能考虑海洋暖流和局部地形的影响,如中国云南西双版纳、越南河内、瓦努阿图等地成功引种,但落在预测区划图的低适宜区。因此,在区划等级为0.4以下的区域,根据可可对几个环境因子的适宜取值范围,选择良好的小气候环境也可尝试种植可可。另外,在本研究的可可适宜分布预测图的一些适宜区域,由于受到台风侵扰,要慎重评估台风可能带来的损失。
参考文献
[1] Hendderson J S, Joyce R A, Hall G R, et al. Chemical and archaeological evidence for the earliest cacao beverages [J]. PNAS, 2007, 104: 18937-18940.
[2] FAO Statistics[EB/OL]. http://faostat3.fao.org/search/cocoa/E, 2015-01-01.
[3] 秦晓威, 郝朝运, 吴 刚, 等. 可可种质资源多样性与创新利用研究进展[J]. 热带作物学报, 2014, 35(1): 188-194.
[4] Jacobi J, Schneider M, Bottazzi P, et al. Agroecosystem resilience and farmers' perceptions of climate change impacts on cocoa farms in Alto Beni, Bolivia[J]. Renewable Agriculture and Food Systems, 2013, 30: 170-183.
[5] Amos T T, Thompson O A. Climate change and the cocoa production in the tropical rain forest ecological zone of Ondo State, Nigeria[J]. Journal of Environment and Earth Science, 5: 36-41.
[6] Asante F A, Amuakwa-Mensah F. Climate change and variability in Ghana: stocktaking [J]. Climate, 2015, 3: 78-99.
[7] Oyekale A S. Climate change induced occupational stress and reported morbidity among cocoa farmers in South-Western Nigeria[J]. Annals of Agricultural and Environmental Medicine, 2015, 22: 357-361.
[8] Laliberte B, Cryer N C, Daymond A J, et al. A global strategy for the conservation and use of cacao genetic resources, as the foundation for a sustainable cocoa economy[J]. 17th Internaitonal Cocoa Research Conference, Yaoundé, Cameroon, 2012: 15-20.
[9] Laderach P, Martinez-Valle A, Schroth G, et al. Predicting the future climatic suitability for cocoa farming of the world's leading producer countries, Ghana and Cote d'Ivoire[J]. Climate Change, 2013, 119: 841-854.
[10] Lawai J O, Omonona B T. The effects of rainfall and other weather parameters on cocoa production in Nigeria[J]. Comunicata Scientiae, 2014, 5: 518-523.
[11] Ofori-Boateng K,Insah B. The impact of climate change on cocoa production in West Africa[J]. International Journal of Climate Change Strategies and Management, 2014, 6: 296-314.
[12] Agbongiarhuoyi A E, Abdulkarim I F, Fawole O P, et al. Analysis of farmers' adaptation strategies to climate change in cocoa production in Kwara State[J]. Journal of Agricultural Extension, 2013, 17: 10-22.
[13] Ruf F, Schroth G, Doffangui K.Climate change, cocoa migrations and deforestation in West Africa: what does the past tell us about the future?[J]. Sustainability Science, 2015, 10: 101-111.
[14] Wanger T C. Pollination curbs climate risk to cocoa[J]. Nature, 2014, 511: 155.
[15] Larson C. The endangered palates list[J]. Science, 2015, 348: 954.
[16] Codjoe F N Y, Ocansey C K, Boateng D O, et al. Climate change awareness and coping strategies of cocoa farmers in rural Ghana[J]. Journal of Biology, Agriculture and Healthcare, 2013, 3: 19-29.
[17] Ajewole D O, Sadiq I. The effect of climate change on cocoa yield in Cocoa Research Institute of Nigeria(CRIN)farm, Oluyole Local Government Ibadan Oyo State[J]. Journal of Sustainable Development in Africa, 2010, 12: 350-358.
[18] Oyekale A S, Bolaji M B, Olowa O W. The effects of climate change on cocoa production and vulnerability assessment in Nigeria[J]. Agricultural Journal, 2009, 4: 77-85.
[19] 郝朝运, 谭乐和, 范 睿, 等. 利用最大熵模型预测药用植物海南蒟的潜在地理布局[J]. 热带作物学报, 2011, 32(8): 1 561-1 566.
[20] 胡 秀, 吴福川, 郭 徽, 等. 基于MaxEnt生态学模型的檀香在中国的潜在种植区预测[J]. 林业科学, 2014, 50: 27-33.
[21] 何奇瑾, 周广胜. 我国玉米种植区分布的气候适宜性[J]. 科学通报, 2012, 57: 267-275.
[22] Thorn J S, Nijman V, Smith D, et al. Ecological niche modeling as a technique for assessing threats and setting conservation priorities for Asian slow lorises(Primates: Nycticebus)[J]. Diversity and Dsitributions, 2009, 15: 289-298.
[23] Phillips S J, Anderson R P, Schapire R, E. Maximum ntropy modeling of species geographic distributions[J]. Ecological Modelling, 2006, 190: 231-259.
[24] 周卫红, 杨正英, 万 帆, 等. 基于GIS的中国春小麦(Triticum aestiyum L.)潜在适宜分布研究[J]. 兰州大学学报(自然科学版), 2009, 45: 1-5.
[25] 胡 秀, 吴福川, 郭 徽. 基于MaxEnt生态学模型的毛姜花潜在园林引种区预测[J]. 中国城市林业, 2013, 4: 29-31.
[26] Elith J, Philips S J, Hastie T, et al. A statistic explanation of MaxEnt for ecologists[J]. Diversity and Distributions, 2011, 17: 43-57.
[27] Elith J, Graham C H, Anderson R P, et al. Novel methods improves prediction of species' distribution from occurrence data[J]. Ecography, 2006, 29: 129-151.
[28] Hijmans R J, Cameron S E, Parra J L, et al. Very high resolution interpolated climate surfaces for global land areas[J]. International Journal of Climatology, 2005, 25: 1 965-1 978.
[29] Bartley B G D. The genetic diversity of cacao and its utilization[M]. U K, Wallingford: CABI Publishing, 2005: 50-55.
[30] Monteiro W R, Lopez U V, Clement D. Genetic Improvement in Cocoa. In: Jain SM, Priyadarshan D(eds.), Breeding Plantation Tree Crops: Tropical Species[M]. Springer Science, Business Media, 2009: 589-626.
[31] Dias LAS. Origin and distribution of Theobroma cacao L: A new scenario. In: Dias LAS(ed.), Genetic improvement of cacao[EB/OL]. http://ecoport.org/, 2004.
[32] Sánchez P A, Jaffé K. Rutas de migraciones humanas precolombinas a la amazonia sugeridas por la distribucion del cacao[J]. Interciencia, 1992, 17, 28-34.
[33] Motamayor J C, Lopez P A, Ortiz C, et al. Sampling the genetic diversity of Criollo cacao in Central and South America[J]. INGENIC Newsletter, 1998, 4: 14-15.
猜你喜欢
疯狂英语·新读写(2021年8期)2021-11-05 08:44:26
数学物理学报(2021年4期)2021-08-30 08:27:50
数学小灵通·3-4年级(2021年6期)2021-07-16 06:54:58
中等数学(2020年1期)2020-08-24 07:57:42
文化创新比较研究(2020年14期)2020-01-02 19:25:56
文化创新比较研究(2020年8期)2020-01-02 04:45:23
小学生优秀作文(高年级)(2018年4期)2018-09-11 01:23:24
少儿科学周刊·儿童版(2016年1期)2016-03-14 04:00:33
山东青年(2016年1期)2016-02-28 14:25:28
湖南水利水电(2015年2期)2015-12-24 02:18:46
