
农杆菌介导的芦笋遗传转化体系的建立
2016-05-30 15:47:21鹿志伟侯晓婉高建明张燕梅杨子平陆军迎李俊峰赵艳龙周文钊易克贤
热带作物学报 2016年8期
关键词:芦笋
鹿志伟 侯晓婉 高建明 张燕梅 杨子平 陆军迎 李俊峰 赵艳龙 周文钊 易克贤


摘 要 以芦笋“井岗701”胚状体为试验材料,在构建pCAMBIA3300-35S-hevein-NOS植物表达载体基础上,采用农杆菌介导的转基因方法,探究菌液浓度、AS浓度、侵染时间和共培养时间等4个因素对芦笋转基因效率的影响,以期建立高效的芦笋转基因体系。结果表明:用菌液浓度OD600=0.6,AS终浓度为200 μmol/L的农杆菌菌液进行侵染,侵染10 min后,暗培养4 d的转基因效率最佳,经PCR检测,阳性转化率达21%,获得了转基因幼苗。本实验构建了完整的农杆菌介导的芦笋遗传转化体系。
关键词 芦笋;农杆菌介导;遗传转化体系
中图分类号 S644.6 文献标识码 A
Abstract Embryoids derived from asparagus“Jinggang 701”were chosen as the experimental materials. With construction of plant expression vector-pCAMBIA3300-35S-hevein-NOS, the effects of liquid bacterial concentration, AS concentration, infection time and co-culture time on asparagus transgenic efficiency were explored by the agrobacterium-mediated transgenic method. The high-efficiency transgenic system of asparagus was expected to be established in this study. The result showed that the optimal infection conditions were liquid bacterial concentration OD600=0.6, adding AS up to final concentration of 200 μmol/L, infecting for 10min and culturing under darkness for 4 d, which made the transformation rate up to 21% and got transgenic plant.
Key words Asparagus(Asparagus officinalis L.);Agrobacterium-mediated;Genetic transformation system
doi 10.3969/j.issn.1000-2561.2016.08.009
芦笋(Asparagus officinalis L.)属于天门冬属的多年生草本植物,在欧洲、亚洲、澳大利亚以及美洲的分布尤为广泛,中国是芦笋的主要生产国之一[1]。芦笋除了具有重要的食用价值外,还对多种疾病具有良好的治疗效果,如降血糖、抗癌等,享有“蔬菜之王”的美誉[2-4]。当前市场上流行的芦笋品种虽然高产但是均易感病,尤其是茎枯病严重影响着芦笋的产量,高产、抗病芦笋新品种的选育已成为当务之急。新品种选育主要有杂交育种和转基因育种2种方式,杂交育种的随机性、过程繁琐、育种周期长等特征致使新品种培育具有很大的不可控性,短期内难以得到抗病高产的芦笋新品种。而转基因育种则不同,其可以对植株性状进行定向的改变,品种选育周期短、可控性高,日益成为品种选育的首选方式。
Hevein基因又称为橡胶树凝集因子基因,其表达产生的橡胶树凝集因子是一个富含Cys和Gly的小分子单链蛋白质,具有结合几丁质的作用,是乳胶中橡胶粒子凝集的主要影响因素,也是乳胶中黄色物质的主要蛋白质之一[5]。后来从甜菜叶片、小麦等植物种子中也发现了类似的hevein-like蛋白,经研究发现hevein和hevein-like蛋白在植物体内和体外还表现出广谱的抗菌性,对细菌、真菌等病原菌都具有良好的抑菌性能[6-8]。
目前,国内外有关芦笋转基因育种的相关研究较少,在国内尚未有相关报道,而国外仅3人对芦笋转基因有过报道:Hernalsteens等[9]将空的根癌农杆菌转入芦笋茎中,并成功地在转化植株中检测到胭脂碱和农杆菌素碱,证明根癌农杆菌成功地转入了芦笋中。Bytebier等[10]将含有NOS-APHⅡ基因的根癌农杆菌成功转入芦笋中;Limanton-Grevet等[11]将含有uidA和nptⅡ基因农杆菌AGL1成功转入芦笋胚性系中,并对后代进行了遗传分析。虽然研究人员已成功将外源基因转入芦笋植株中,但是转化体系中转化率均较为低下,转化周期长,且均无目的基因转入。笔者在前人研究的基础上,以芦笋胚状体作为外植体,在构建含有35S-hevein-NOS表达元件的pCAMBIA3300植物表达载体基础上,探究菌液浓度、AS浓度、侵染时间和共培养时间等4个因素对农杆菌介导的芦笋转基因效率的影响,以期进一步完善农杆菌介导的芦笋遗传转化体系,提高遗传转化率,并导入目的基因,为后期芦笋转基因抗病高产新品种选育奠定研究基础。
1 材料与方法
1.1 材料
植物表达载体:pBI121、pCAMBIA3300,均由中国热带农业科学院热带生物技术研究所张树珍实验室惠赠。农杆菌EHA105菌株和Puc57-hevein由本实验室保存。限制性内切酶均为fermentas公司产品,DNA聚合酶和连接酶为NEB公司产品,其它分析纯药品及试剂盒。芦笋“井岗701”购自江西省农业科学院。
1.2 方法
1.2.1 外植体制备 外植体制备方法参照鹿志伟等[12]的研究方法。
1.2.2 植物表达载体构建 设计5′TGCTCTAGAAT
GAAATACTGTACTATGTTTAT3′(添加XbaⅠ酶切位点)和5′CGAGCTCTCAGTTGGCACCGC3′(添加SacⅠ酶切位点)引物对hevein基因进行扩增,随后对扩增得到的目的片段进行电泳检测。准确无误后,对扩增得到的目的基因和pBI121进行XbaⅠ和SacⅠ双酶切,分别使用PCR产物回收试剂盒和胶回收试剂盒对二者进行目的片段回收,将回收得到的目的基因和pBI121大片段进行T4 DNA连接酶连接,获得含hevein基因的pBI121载体(图1)。随后分别对改造后的pBI121载体和pCAMBIA3300进行HindⅢ和EcoRⅠ双酶切,使用胶回收试剂盒对pBI121进行小片段回收,而对pCAMBIA3300则进行大片段回收,对得到的2个片段进行T4 DNA连接酶连接,从而完成含有35S-hevein-NOS基因表达元件的pCAMBIA3300植物表达载体构建(图2),最后采用冻融法将构建好的植物表达载体转入农杆菌EHA105中,-20 ℃备用。
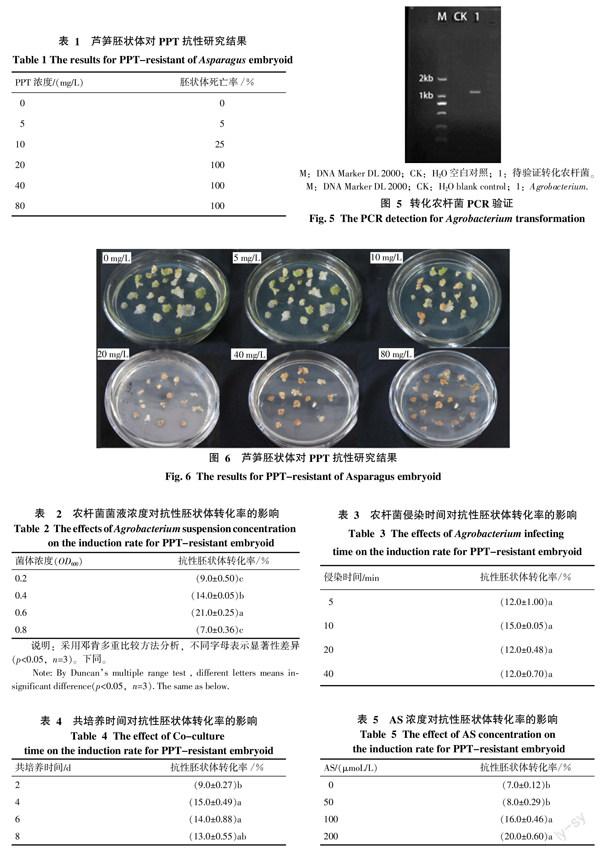
1.2.3 芦笋胚状体对PPT的抗性实验 将芦笋胚状体切成0.6 cm2大小,添加至含0、5、10、20、40、80 mg/L PPT的转化培养基中,每组实验添加20块外植体,重复3次,培养20 d后,观察芦笋胚状体生长状况并统计死亡率。
1.2.4 预培养 将培养好的芦笋胚状体切成0.6 cm2左右的小块,置于预培养基上,26 ℃光培养2 d。预培养培养基:MS+4%蔗糖+800 mg/L谷氨酰胺+500 mg/L酸水解酪素+0.70 mg/L嘧啶醇+0.10 mg/L NAA+0.50 mg/L kinetin,pH5.8。
1.2.5 转化菌液的制备 取-70 ℃保存含pCAMBI
A3300-35S-Hevein-NOS质粒的转化农杆菌菌液20 μL,置于YEP固体培养基(含Kanamycin 50 mg/L和Rifampicin100 mg/L)中,涂布均匀。静置30 min,28 ℃倒置培养2 d。挑取单菌落接种至含Kanamycin 50 mg/L和Rifampicin 100 mg/L 的YEP液体培养基中,28 ℃、250 r/min震荡培养直至对数期(OD600约为0.55左右),5 500 r/min,-4 ℃离心8 min,收集菌体,液体转化培养基悬浮。
1.2.6 侵染和共培养 使用液体转化培养基将上述菌体OD600值分别调节为0.2、0.4、0.6、0.8。将预培养2d芦笋胚状体置于150 mL无菌锥形瓶中,同时锥形瓶中添加50 mL含转化农杆菌菌体和乙酰丁香酮(浓度分别为0、50、100、200 μmol/L)的液体转化培养基,从而对胚状体进行农杆菌侵染,侵染时间为5、10、20、40 min,26 ℃ 150 r/min振荡。无菌滤纸擦干芦笋胚状体表面液体,转移至固体转化培养基中,26 ℃,暗培养2、4、6、8 d。
1.2.7 脱菌与选择培养 使用200 mg/L Timentin对共培养后的芦笋胚状体进行脱菌10 min,100 r/min轻轻振荡,然后无菌水清洗3次。重复3次。无菌滤纸擦干芦笋胚状体表面液体。最后转移至含Timentin和PPT的固体转化培养基中,26 ℃,光培养,直至原有胚状体表面有新的胚状体生成,上述每组实验添加80块胚状体,重复3次,统计抗性胚状体转化率,抗性胚状体转化率/%=抗性胚状体个数/胚状体处理总个数×100。
1.2.8 分子水平检测 PCR检测是一种非常简单、快速和直接的转基因苗检测方法,本实验用此方法对抗性胚状体进行检测。胚状体经PPT筛选后进行PCR验证以及植株再生。采用OMEGA植物DNA提取试剂盒对抗性胚状体进行DNA提取,并进行PCR检测,统计阳性转化率。
1.2.9 抗性植株再生 将选择培养后新长出的胚状体转移到生苗培养基中,进行苗诱导。
1.3 数据分析
使用SPSS19.0和Excel2007软件对试验数据进行统计分析[13-14]。
2 结果与分析
2.1 植物表达载体构建
2.1.1 pBI121-Hevein载体的构建 对pBI121-hevein植物表达载体进行PCR扩增以及电泳分析,发现有且仅有一条约276 bp目的基因条带,与目的基因hevein相符(图3)。
2.1.2 植物表达载体pCAMBIA3300-35S-Hevein-NOS的构建 对pCAMBIA3300-35S-Hevein-NOS植物表达载体进行双酶切以及电泳分析,发现经双酶切后载体被分为2个片段,其中小片段大小为1 100 bp左右,与目的基因表达元件35S-Hevein-NOS相符(图4)。
2.1.3 植物表达载体pCAMBIA3300-35S-Hevein-NOS转化农杆菌及其验证 对pCAMBIA3300-35S-Hevein-NOS植物表达载体进行菌落PCR验证,发现在PCR目的条带大小为1 100 bp左右,与目的基因表达元件35S-Hevein-NOS相符(图5)。
2.2 芦笋胚状体对PPT抗性的研究
由表1和图6可以看出,随着PPT浓度的增加,芦笋胚状体发生萎缩、变黄直至死亡,死亡率逐渐增加,PPT浓度为20 mg/L时胚状体全部死亡。
2.3 不同农杆菌菌液浓度和侵染时间对芦笋遗传转化的影响
由表2可知,不同菌体浓度对抗性胚状体的转化率存在显著性差异(p<0.05),当菌体浓度为0.6时,转化率最高。由表3可知,不同侵染时间对抗性胚状体的转化率不存在显著性差异(p<0.05),为达到侵染时间最短以及转化率较高的目标,侵染时间选择10 min为佳。因此,农杆菌菌液浓度OD600=0.6,侵染时间10 min时为最佳作用条件。
2.4 不同共培养时间对芦笋遗传转化的影响
由表4可知,当共培养时间为4 d和6 d时,抗性胚状体转化率与其它处理相比差异显著,转化率分别为15.0%和14.0%。为了节省培养时间,同时又使转化率达到最高,共培养时间选择4 d为佳。
2.5 不同浓度AS对芦笋遗传转化的影响
由表5可知,不同AS浓度对抗性胚状体转化率存在显著性差异(p<0.05)。当AS浓度为100、200 μmoL/L时,抗性胚状体转化率与其它处理相比差异显著,转化率分别为16.0%和20.0%。为了使转化率达到最高,AS添加浓度选择200 μmoL/L作为最佳。
2.6 转基因植株的PCR检测
由图7可知,阳性对照和再生芦笋植株PCR结果中目的条带大小一致,约为1 100 bp,大小与基因表达元件35S-Hevein-NOS相符。整个转基因过程见图8。
3 讨论与结论
农杆菌介导的植物遗传转化是一个非常复杂的过程,受外植体类型、菌液浓度、菌株类型、侵染时间、共培养时间、以及乙酰丁香酮浓度等多种因素影响,而且不同植物基因型、外植体其对应的农杆菌最佳侵染条件不同,转化率也具有很大差异。农杆菌作为转化效果最好的植物转化方式之一,目前为止已有很多相关研究,如周月等[15]通过农杆菌介导法将LJAMP2基因成功导入“红阳”泥猴桃中,其使用的最佳农杆菌侵染条件为:共培养时间为2 d,农杆菌菌液浓度为OD600=0.5,侵染时间为10 min,AS浓度为100 μmol/L,转化率达到5.11%;Gnasekaran等[16]农杆菌介导的兰花转基因中,最佳的农杆菌侵染条件为:OD600=0.8,侵染时间30 min,共培养4 d,AS浓度为200 μmol/L,转化率高达33.6%。
农杆菌介导的芦笋遗传转化体系的研究已有部分工作,如Bytebier等[17]使用愈伤组织作为外植体,经过侵染、选择培养以及植株再生等得到了转基因芦笋植株,但是转化周期较长。Bruno等[18]使用C58农杆菌菌株,同时对3个基因型的芦笋体细胞进行转化,检测得到阳性植株,但是转化率较低,为0.6%~4%。Limanton-Grevet等[19]使用AGL1Gin农杆菌菌株,选用5个胚性系作为侵染对象,结果发现转化率为0.8%~12.8%,转化率得到提高。本研究在综合对比分析前人研究的基础上,以芦笋胚状体作为侵染对象,采用EHA105农杆菌菌株,并对菌体浓度、侵染时间、共培养时间等侵染条件进行优化。结果表明,芦笋转化率大大提高,最高可达21%。转化周期得到有效地缩短,遗传效率得到极大地提高。另外,实验中成功将hevein基因和bar基因转入芦笋植株中,有利于后期抗病、抗除草剂芦笋新品种的选育。
参考文献
[1] Huang X F, Lin Y Y, Kong L Y. Steroids from the roots of Asparagus officinalis and their cytotoxic activity[J]. Journal of Integrative Plant Biology, 2008, 50(6): 717-722.
[2] Nindo C I, Sun T, Wang S W, et al. Evaluation of drying technologies for retention of physical quality and antioxidants in Asparagus(Asparagus officinalis L.)[J]. Food Science and Technology, 2003, 36(5): 507.
[3] Hafizur R M, Kabir N,Chishti S. Asparagus officinalis extract controls blood glucose by improving insulin secretion and β-cellfunction in streptozotocin -induced type 2 diabetic rats[J]. British Journal of Nutrition, 2012, 108: 1 586-1 595.
[4] Jang D S, Cuendet M, Fong H H S, et al. Constituents of Asparagus officinalis evaluated for inhibitory activity against cyclooxygenase-2[J]. J Agric Food Chem, 2004, 52: 2 218-2 222.
[5] Gidrol X, Chrestin H, Tan H L, et al. Hevein, a lectin-like protein from Hevea brasiliensis (rubber tree) is involved in the coagulation of latex[J]. J BIOl Chem, 1994, 269: 9 278.
[6] 刁现民, 王永芳, 李 伟, 等. 抗除草剂基因Bar转化谷子研究[C]. 全国作物遗传育种学术研讨会论文集, 2003: 613-618. [7] Davis E G, Sang Y M, Blecha F. Equine β-defensin-1:full-length cDNA sequence and tissue expression[J]. Vet Immunol Immunopathol, 2004, 99: 127-132.
[8] De Leeuw E, Li C Q, Zeng P Y, et al. Functional interaction of human neutrophil peptide-1 with the cell wall precursor lipidⅡ[J]. FEBS Lett, 2010, 584(8): 1 543-1 548.
[9] Hernalsteens J P, Thia-Toong L, Schell J, et al. An Agrobacterium-transformed cell culture from the monocot Asparagus officinalis[J]. The EMBO Journal, 1984, 3(13): 3 039-3 041.
[10] Bytebier B, Deboeck F, Greve H D, et al. T-DNA organization in tumor cultures and transgenic plants of the monocotyledon Asparagus officinalis[J]. Proc Nati Acad Sci, 1987, 84(15): 5 345-5 349.
[11] Limanton-Grevet A, Jullien M. Agrobacterium-mediated transformation of Asparagus officinalis L.: molecular and genetic analysis of transgenic plants[J]. Molecular Breeding, 2001, 7(2): 141-150.
[12] 鹿志伟, 高建明, 侯晓婉,等. 芦笋组培快繁体系的建立[J]. 北方园艺, 2015(9): 83-86.
[13] 刘瑞江, 张业旺,闻崇炜,等. 正交试验设计和分析方法研究[J]. 实验技术与管理, 2010, 27(9): 52-55.
[14] 高忠江, 施树良, 李 钰. SPSS方差分析在生物统计的应用[J]. 现代生物医学进展, 2008, 8(11): 2 116-2 120.
[15] 周 月, 赵许朋, 吴秀华,等. 农杆菌介导LJAMP2基因导入‘红阳猕猴桃及分子鉴定[J]. 生物工程学报, 2014, 30(6):931-942.
[16] Gnasekaran P, Uddain J, Subramaniam S, et al. Agrobacterium-mediated transformation of the recalcitrant Vanda Kasems Delight Orchid with higher efficiency[J/OL]. The Scientific World Journal, 2014: http://dx.doi.org/10.1155/2014/583934.
[17] Bytebier B, Deboeck F, Greve H D, et al. T-DNA organization in tumor cultures and transgenic plants of the monocotyledon Asparagus officinalis[J]. Proc Nati Acad Sci, 1987, 84(15): 5 345-5 349.
[18] Bruno Delbreil, Phifippe Guerche t, M. Jullien. Agrobacterium-mediated transformation of Asparagus officinalis L. long-term embryogenic callus and regeneration of transgenic plants[J]. Plant Cell Reports, 1993, 12: 129-132.
[19] Limanton-Grevet A, Jullien M. Agrobacterium-mediated transfo-
rmation of Asparagus officinalis L.: molecular and genetic analysis of transgenic plants[J]. Molecular Breeding, 2001, 7(2): 141-150.
猜你喜欢
今日农业(2022年15期)2022-09-20 06:55:30
中老年保健(2022年5期)2022-08-24 02:35:50
——防口干
饮食科学(2019年2期)2019-11-22 05:06:09
发明与创新·中学生(2019年4期)2019-04-19 12:06:36
发明与创新(2019年14期)2019-03-18 07:41:24
今日农业(2019年14期)2019-01-04 08:57:40
现代园艺(2017年19期)2018-01-19 02:50:04
世界热带农业信息(2016年9期)2017-04-14 13:21:16
老友(2016年5期)2016-04-02 23:07:27
发明与创新(2015年29期)2015-02-27 10:39:41
