
施氮肥对东北帽儿山云杉人工林土壤微生物生物量和群落结构的影响1)
2016-05-30 05:57尉建埔张洁王文娜王政权谷加存
东北林业大学学报 2016年5期
尉建埔 张洁 王文娜 王政权 谷加存
(东北林业大学,哈尔滨,150040)
施氮肥对东北帽儿山云杉人工林土壤微生物生物量和群落结构的影响1)
尉建埔张洁王文娜王政权谷加存
(东北林业大学,哈尔滨,150040)
摘要采用磷脂脂肪酸(PLFA)方法,研究了施氮肥对东北帽儿山地区云杉人工林的土壤微生物生物量和群落结构的影响。结果表明:与对照相比,施肥使土壤微生物总生物量在表层(0~10 cm)和亚表层(>10~20 cm)分别下降了28.5%和11.5%。施肥也使细菌、真菌、革兰氏阳性菌生物量比对照显著降低,但是革兰氏阴性菌比对照组显著增加,而对放线菌无显著影响。施肥导致土壤部分理化因子也发生了显著改变,特别是施肥处理下硝态氮质量分数在表层和亚表层分别提高了10倍和6倍,而pH值在表层和亚表层显著下降。因此,施氮肥可以显著改变云杉人工林土壤的理化性质,导致土壤微生物生物量下降和土壤微生物群落结构的改变。
关键词云杉人工林;群落结构;土壤微生物;生物量;磷脂脂肪酸
分类号S725.5
Effect of Nitrogen Fertilization on Soil Microbial Biomass and Community Structure ofPiceakoraiensisPlantation in Maoershan Mountains of Northeastern China//
Wei Jianpu, Zhang Jie, Wang Wenna, Wang Zhengquan, Gu Jiacun
(Northeast Forestry University, Harbin150040, P. R. China)//Journal of Northeast Forestry University,2016,44(5):52-56.

KeywordsPicea koraiensis; Community structure; Soil microbe; Biomass; PLFA
土壤微生物在陆地生态系统碳(C)和养分循环过程中发挥重要作用(参与凋落物分解过程、调节土壤有机质的储存和矿化等)[1-3]。土壤微生物的组成是土壤健康的重要指标,对维持陆地生态系统的稳定、林木生长以及森林净初级生产力具有重要意义[4-7]。
土壤微生物对生境变化反应敏感,受气候和土壤状况等影响显著[8]。大量研究表明,氮(N)添加会导致林地土壤微生物生物量、群落结构和功能发生变化[9]。He Yating et al.[10]研究证实,长期施N肥可导致阔叶林土壤微生物生物量显著降低。Treseder[11]在总结82个研究案例的Meta分析中发现,在施N肥条件下土壤微生物生物量平均降低15%,长期或大量施N肥可导致细菌和真菌丰富度显著降低。在野外林地和实验室培养条件下,施N肥均对土壤微生物生长产生抑制作用[9],这主要是因为土壤理化性质发了显著的变化[9-12]。土壤N增加还对土壤微生物群落结构产生重要影响,如导致真菌/细菌比值[8]及革兰氏阳性细菌(G+)/革兰氏阴性细菌(G-)比值发生变化[13]。这些微生物群落结构的改变可能会对一系列的土壤生态学过程有重要的影响,如影响凋落物和土壤有机质的分解速率[2],改变硝化和反硝化的微生物过程[14],增加温室气体N2O的排放[15]等。因此,研究N添加对土壤微生物生物量和群落结构的影响对于了解土壤健康状况,评估土壤肥力及有针对性的进行土壤改良和优化林地管理措施等有重要的意义。
红皮云杉(Piceakoraiensis)是东北地区常见的针叶树种,其蓄积量占黑龙江省天然林总面积的23%[16],目前已营造大面积人工林。虽然以往针对东北地区云杉人工林的土壤特征进行了较多研究[6,14],但是施N肥对云杉林土壤微生物的影响了解甚少。为此,本实验通过对28年生云杉人工林进行短期(1 a)的硝酸铵施肥处理,研究土壤N有效性增加对土壤理化性质、微生物(包括细菌、真菌和放线菌)生物量和群落结构(G+和G-)的影响,目的是为优化东北地区云杉人工林林地营养管理措施提供参考。
1研究区概况
试验地位于东北林业大学帽儿山实验林场尖砬沟试验站(127°30′~127°34′E,45°21′~45°25′N)。该地区气候属温带大陆性季风气候,年均降水量723.8 mm,年均气温2.8 ℃,其中1月份平均气温为-19.6 ℃,7月份平均气温为20.9 ℃,年无霜期120~140 d。该地区平均海拔300 m,土壤类型为暗棕壤,土层平均厚度在40~50 cm,有机质丰富,排水状况良好。
2研究方法
样地设置:2014年5月,在云杉人工林内随机设置6块10 m×10 m的样地,在每块样地核心区设置1块4 m×4 m的施肥样方和1个4 m×4 m对照样方(每个处理3个重复)。分别于2014年5、7、8月份在试验样方内施N肥(硝酸铵)。总施N量为10 g·m-2·a-1,根据季节温度变化调节施肥量,其中5月份施30%、7月份施40%、8月份施30%。


土壤微生物测定:采用磷脂脂肪酸分析法(PLFA)测定土壤微生物生物量总量、细菌、真菌和放线菌生物量以及群落结构[18]。主要步骤为:精确称取1.00 g冻干土样,经过脂类抽提、固相抽提柱层析分离脂类、磷脂的碱性甲醇水解和甲基化之后,应用气相色谱—质谱联用仪(GC-MS:Varian 450-GC,Varian 240-MS,美国)上机测试。磷脂脂肪酸的定性根据与标准物质(十九酸甲酯)的相对保留时间及质谱图与数据库进行对比确定。磷脂脂肪酸的含量由内标法确定。代表微生物各种群的特征磷脂脂肪酸主要为:饱和脂肪酸及单不饱和脂肪酸代表细菌(12:0、i14:0、14:0、i15:0、a15:0、15:0、i16:0、a16:0、16:0、i17:0、17:0、16:1ω9,cy17:0、18:0、cy19:0、21:0,22:0);其中i15:0、a15:0、15:0、i16:0、a16:0、i17:0代表革兰氏阳性菌,cy17:0、cy19:0、16:1ω9代表革兰氏阴性菌;18:2ω6、18:3ω6、18:1ω9c、18:1ω9t代表真菌;10-Me分支饱和脂肪酸代表放线菌[19]。
数据处理:采用SPSS13.0软件进行描述统计和正态检验。运用成对样本T检验,检验土壤理化性质和土壤微生物数量在施肥后变化的显著性。
3结果与分析
3.1施肥对云杉林土壤理化性质的影响


表1 施肥与对照处理的云杉林土壤化学特征
注:表中数值为“平均数±标准误”;同列不同字母表示同一土层内差异达到显著水平(P<0.05)。
3.2施氮肥对土壤微生物总生物量的影响
施肥导致土壤表层和亚表层的土壤微生物总量均显著低于对照(P<0.05)(见表2)。表层土壤微生物的PLFA标记物量由1 479.0 nmol·g-1降低到1 057.4 nmol·g-1,降幅为28.5%;而在亚表层中则由836.5 nmol·g-1降低到739.9 nmol·g-1,降幅为11.5%。此外,无论施肥与否,表层土壤微生物总量均显著高于亚表层。
3.3施氮肥对土壤细菌、真菌、放线菌生物量的影响
由表2可知,施肥导致云杉林土壤细菌数量显著降低(P<0.05)。在表层土中,施肥使土壤细菌的PLFA标记物量由1 267.2 nmol·g-1下降到885.9 nmol·g-1,降幅为30.1%;在亚表层土则由739.9 nmol·g-1下降到621.0 nmol·g-1,降幅为16.1%。细菌中G+细菌的PLFA标记物含量在表层降低显著(P<0.05),由224.3 nmol·g-1下降到145.3 nmol·g-1,降幅为41.0%;而亚表层土的G+细菌标记物含量下降不显著(P>0.05)。与G+细菌不同,G-细菌的PLFA标记物含量因施肥而显著增加(P<0.05),在表层和亚表层分别增加了51.8%和31.0%。因此,表层和亚表层土壤细菌的G+/G-分别由1.42和1.21降低至0.56和0.85。
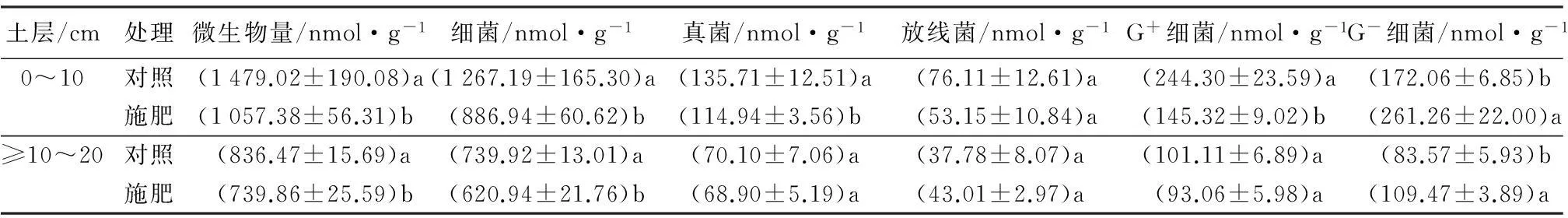
表2 施肥与对照处理的云杉林各类群微生物PLFA标记物含量
注:表中数值为“平均数±标准误”;同列不同字母表示同一土层内差异达到显著水平(P<0.05)。
施肥对真菌生物量也有显著影响(P<0.05)。在表层土壤中,真菌PLFA标记物的数量由135.7 nmol·g-1显著降低到115.0 nmol·g-1,降幅为15.3%;而亚表层土的真菌生物量的变化并不显著(P>0.05)。施肥对土壤细菌和真菌均产生了显著的影响,但并没有显著影响土壤真菌/细菌的比例。此外,施肥对放线菌的影响不显著(P>0.05)。
4讨论与结论
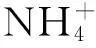
施肥不但对土壤养分产生影响,而且对云杉林土壤的微生物生物量也产生重要影响。磷脂脂肪酸法(PLFA)的测定结果显示,施N肥在短期内直接导致了土壤各层的PLFA表征的微生物总生物量的显著下降,说明通过施肥引起的土壤N有效性增加,会抑制土壤微生物数量的增长,该结果与以往的大部分研究一致[9]。Demoling et al[24]对瑞典挪威云杉林的研究发现,施N肥使挪威云杉(Piceaabies)林土壤微生物数量下降了40%,而微生物活性也下降了30%;Deforest et al[25]在美国密西根北部的北方阔叶林的研究显示,施N肥样地土壤微生物生物量比对照减少了68%;此外,贾淑霞等[26]对帽儿山地区落叶松和水曲柳人工林的研究也得到类似的结果。造成土壤微生物数量下降的原因可能有以下几方面:首先,N添加可以导致土壤pH的下降,而pH的下降在以往的研究中也被证明可以抑制微生物数量和生物量的增长[9];其次,N添加可以改变土壤酶活性,导致土壤酶生产力和分解效率的改变,进而影响土壤微生物数量和生物量的变化。Carreiro et al[27]研究发现,沼生栎(Quercuspalustris)凋落物分解速率和氧化酶活性随N添加而下降,导致氧化酶对底物的利用效率降低,随之带来土壤微生物生物量的减少。此外,本研究发现施肥之后,微生物生物量降低可能还与细菌和真菌生物量同时降低有密切关系,但与放线菌关系较弱,因为施N肥对放线菌生物量影响并不显著。
施N肥改变了土壤微生物群落结构,如改变了土壤真菌/细菌比率。一般认为,N添加会降低土壤细菌和真菌的生物量,且对真菌的影响大于细菌[28-29],但本研究中真菌/细菌的比例并未有显著变化,可能是由于施肥时间短或施肥总量不足以显著改变二者的关系;施肥同时也导致细菌群落内部结构的改变,如G+细菌生物量降低,G-细菌生物量增加,从而降低了G+/G-比,这与其他施肥研究的结果具有一致性[13]。这可能因为G+细菌能够适应条件较差的土壤环境,且在底物资源有限的情况下更擅长竞争资源[30];而G-细菌的生存更依赖于土壤中易分解的新鲜有机物[31]。向土壤中施加N肥,增加了土壤有效N的质量分数,缓解了土壤营养条件对微生物的限制[18],土壤的G-在表层和亚表层均显著增加,而G+减少,G+/G-比显著降低。G+细菌和G-细菌是土壤细菌中重要的组成部分,对陆地生态系统循环,尤其是对枯落物的分解起重要的作用[32],因此,施肥对云杉人工林凋落物分解过程可能具有重要的影响,还需要进一步的研究验证。一些研究发现N添加有助于土壤放线菌数量的增加,原因是N添加解除了土壤N对放线菌的限制,增加了土壤养分的活性,利于放线菌的生长[33-34]。本研究中施肥并未导致土壤放线菌发生显著变化,原因可能是N添加导致云杉林地土壤的酸化(pH下降)和部分阳离子的流失及土壤养分失衡,这些均对放线菌产生了负面的影响,但需要其他试验进一步验证。
参考文献
[1]BOYLE S A, ROCKIE R Y, PETER J. Bacterial and fungal contributions to soil nitrogen cycling under Douglas fir and red alder at two sites in Oregon[J]. Soil Biology and Biochemistry,2008,40(2):443-451.
[2]HOBBIE S E, VITOUSEK P M. Nutrient limitation of decomposition in Hawaiian forest[J]. Ecology,2000,81(7):1867-1877.
[3]EISENLORD S D, FREEDMAN Z, ZAK D R, et al. Microbial mechanisms mediating increased soil C storage under elevated atmospheric N deposition[J]. Applied and Environmental Microbiology,2013,79(4):1191-1199.
[4]巨天珍,任海峰,孟凡涛,等.土壤微生物生物量的研究进展[J].广东农业科学,2011,38(16):45-47.
[5]赵超,彭赛,阮宏华,等.氮沉降对土壤微生物影响的研究进展[J].南京林业大学学报(自然科学版),2015,39(3):149-155.
[6]ZAK D R, GRIGAL D F, GLEESON S, et al. Carbon and nitrogen cycling during old-field succession: constraints on plant and microbial biomass[J]. Biogeochemistry,1990,11(2):111-129.
[7]PIAO H C, HONG Y T, YUAN Z Y. Seasonal changes of microbial biomass carbon related to climatic factors in soils from karst areas of southwest China[J]. Biology and Fertility of Soils,2000,30(4):294-297.
[8]刘爽,王传宽.五种温带森林土壤微生物生物量碳氮的时空格局[J].生态学报,2010,30(12):3135-3143.
[9]COMPTON J E, WATRUDA L S, PORTEOUSA L A, et al. Response of soil microbial biomass and community composition to chronic nitrogen additions at Harvard forest[J]. Forest Ecology and Management,2004,196(1):143-158.
[10]HE Yating, QI Yuchun, DONG Yunshe, et al. Effects of nitrogen fertilization on soil microbial biomass and community functional diversity in temperate grassland in inner mongolia, China[J]. Clean-Soil Air, Water,2013,41(12):1216-1221.
[11]TRESEDER K K. Nitrogen additions and microbial biomass: a meta-analysis of ecosystem studies[J]. Ecology Letters,2008,11(10):1111-1120.
[12]VITOUSEK P M, ABER J D, HOWARTH R W, et al. Human alteration of the global nitrogen cycle: sources and consequences[J]. Ecological Applications,1997,7(3):737-750.
[13]毕捷,张乃莉,梁宇,等.氮水添加对半干旱地区草原土壤微生物群落结构的交互影响[J].广东农业科学,2012,39(19):166-170.
[14]INSELSBACHER E, ÖHLUND J, JMTÅRD S, et al. The potential of microdialysis to monitor organic and inorganic nitrogen compounds in soil[J]. Soil Biology and Biochemistry,2011,43(6):1321-1332.
[15]杜亚琴,郑丽行,樊小林.三种控释肥在赤红壤中的氧化亚氮排放[J].应用生态学报,2011,22(9):2370-2376.
[16]王庆贵,邢亚娟,周晓峰,等.黑龙江省东部山区谷地红皮云杉生态学与生物学特性[J].东北林业大学学报,2007,35(3):4-6.
[17]国家林业局.中华人民共和国林业行业标准:森林土壤分析方法[M].北京.中国标准出版社,2000.
[18]FROSTEGÅRD A, TUNLID A, BÅÅTH E. Microbial biomass measured as total lipid phosphate in soils of different organic content[J]. Journal of Microbiological Method,1991,14(3):151-163.
[19]颜慧,蔡祖聪,钟文辉.磷脂脂肪酸分析方法及其在土壤微生物多样性研究中的应用[J].土壤学报,2006,43(5):851-859.
[20]李学垣.土壤化学[M].北京:高等教育出版社,2001:233-243.
[21]INGWERSEN J, BUTTERBACH-BAHL K, GASCHE R, et al. Barometric process separation: New method for quantifying nitrification, denitrification, and nitrous oxide sources in soils[J]. Soil Science Society of America,1999,63(1):117-128.
[22]KRONZUCKER H J, GLASS A D M, SIDDIQI M Y. Inhibition of nitrate uptake by ammonium in barely. Analysis of component flues[J]. Plant Physiology,1999,120(1):283-292.
[23]BERGKVIST B O, FOLKESON L. Soil acidification and element fluxes of a Fargus sylvatica forest as influenced by simulated nitrogen deposition[J]. Water, Air, and Soil Pollution,1992,65(1/2):111-113.
[24]DEMOLING F, NILSSON L O, BÅÅTH E. Bacterial and fungal response to nitrogen fertilization in three coniferous forest soils[J]. Soil Biology and Biochemistry, 2008, 40(2):370-379.
[25]DEFOREST J L, ZAK D R, PREGITZER K S, et al. Atmospheric nitrate deposition and the microbial degradation of cellobiose and vanillin in a northern hardwood forest[J]. Soil Biology and Biochemistry, 2004, 36(6):965-971.
[26]贾淑霞,赵妍丽,孙玥,等.施肥对落叶松和水曲柳人工林土壤微生物生物量碳和氮季节变化的影响[J].应用生态学报,2009,20(9):2063-2071.
[27]CARREIRO M M, SINSABAUGH R L, REPERT D A, et al. Microbial enzyme shifts explain litter decay responses to simulated nitrogen deposition[J]. Ecology,2000,81(9):2359-2365.
[28]FREY S D, KNOOR M, PARRENT J L, et al. Chronic nitrogen enrichment affects the structure and function of the soil microbial community in temperate hardwood and pine forests[J]. Forest Ecology and Management,2004,196(1):159-171.
[29]刘彩霞,焦如珍,董玉红,等.应用PLFA方法分析氮沉降对土壤微生物群落结构的影响[J].林业科学,2015,51(6):155-162.
[31]GRIFFITHS B S, RITZ K, EBBLEWHITE N, et al. Soil microbial community structure: effects of substrate loading rates[J]. Soil Biology and Biochemistry,1999,31(1):145-153.
[32]张圣喜,陈法霖,郑华,等.土壤微生物群落结构对中亚热带三种典型阔叶树种凋落物分解过程的响应[J].生态学报,2011,31(11):3020-3026.
[33]ZHANG Q C, WANG G H, YAO H Y. Phospholipid fatty acid patterns of microbial communities in paddy soil under different fertilizer treatments[J]. Journal of Environmental Sciences,2007,19(1):55-59.
[34]FREEDMAN Z, EISENLORD S D, ZAK D R, et al. Towards a molecular understanding of N cycling in northern hardwood forests under future rates of N deposition[J]. Soil Biology and Biochemistry,2013,66(11):130-138.
收稿日期:2015年9月26日。
第一作者简介:尉建埔,男,1990年5月生,东北林业大学林学院,硕士研究生。E-mail:254487956@qq.com。通信作者:谷加存,东北林业大学林学院,副教授。E-mail:gjcnefu@163.com。
1)国家林业科技支撑计划课题(2012BAD21B0202)。
责任编辑:王广建。
猜你喜欢
今日农业(2020年19期)2020-12-14
江苏农业科学(2016年8期)2017-02-15
江苏农业科学(2016年8期)2017-02-15
热带农业科学(2016年11期)2017-01-21
热带农业科学(2016年11期)2017-01-21
中国科技信息(2015年2期)2015-11-16
河北渔业(2015年7期)2015-07-21
河北渔业(2015年6期)2015-06-24
植物营养与肥料学报(2014年1期)2014-03-11
中国烟草学报(2012年3期)2012-04-10
