
黄芩悬浮细胞培养过程中黄芩苷含量与POD活性的相关性分析
2016-05-14 08:41张东向焦战战张令昂李升林孟庆伟刘丽洁
湖北农业科学 2016年7期
张东向 焦战战 张令昂 李升林 孟庆伟 刘丽洁


摘要:以黄芩悬浮细胞为材料,研究其生长过程中过氧化物酶(POD)活性变化规律及其与黄芩苷次生合成之间的关系。结果表明,在黄芩悬浮细胞生长周期(21 d)内,POD活性与黄芩苷的积累呈正相关。在黄芩悬浮细胞生长周期的前6 d,悬浮细胞处于适应期,POD低水平表达,黄芩苷合成缓慢;6~15 d,悬浮细胞处于增长期,POD高水平表达,并到达最大值,期间黄芩苷迅速合成;15~21 d,悬浮细胞进入饱和期,此时POD活性降低,黄芩苷合成几乎停止。采用L9(3)正交表优化培养基中6-BA,碳源、氮源、pH的配比,筛选POD活性表达最优的培养条件为:6-BA 3 mg/L、碳源15g/L蔗糖+15 g/L葡萄糖、氮源(NH4+)30 mmol/L、pH 5.6。
关键词:黄芩(Scutellaria baicalensis Georgi);悬浮细胞;黄芩苷;过氧化物酶活性;正交设计
中药黄芩为唇形科植物黄芩的干燥根,具有清热燥湿、泻火解毒、止血安胎等功效。其主要成分黄芩苷(Baicalin)具有抗菌、抗病毒、抗炎、抗变态反应、抗氧化、清除氧自由基、抗癌、抗肿瘤、抗凝、抗血栓形成和保护肝脏、心脑血管、神经元等作用。黄芩苷是黄芩细胞次生代谢的一种,它本身就可以代表细胞次生代谢的旺盛程度。植物次生代谢途径研究方面也已取得了不少进展,但是关于次生代谢产物产生机理的研究并不多。次生代谢过程是各种酶促反应的过程,受关键酶与限速酶的调控,关键酶的表达决定代谢途径的启动及相关特定物质的合成。黄芩体内过氧化物酶(POD)能够催化黄芩有效成分黄芩苷与过氧化氢反应生成黄芩素,还可以清除脂类氢过氧化物,减轻有机氢过氧化物对机体的损伤,防御活性氧等自由基对细胞膜的损伤,有一定的抗衰老作用。任振兴等研究表明,黄芩愈伤组织中多酚氧化酶活性与黄芩苷合成有相关性。本试验研究了黄芩悬浮细胞中黄芩苷与过氧化物酶活性相关性,通过对黄芩悬浮细胞增长量、黄芩苷的含量、过氧化物酶活性的测定及分析结果,讨论三者之间的相关性。
1.材料与方法
1.1试验材料
黄芩悬浮细胞,来源于本试验室诱导的黄芩愈伤组织,培养基为液体SM培养基(去除琼脂MS培养基)。
试验仪器有YPL-1全温摇瓶柜、T6新世纪型紫外可见分光光度计、GTR16-2型高速台式冷冻离心机。
1.2试验方法
1.2.1黄芩悬浮细胞鲜重、干重增长曲线和黄芩苷、过氧化物酶活性变化曲线将2 g黄芩愈伤组织研磨成细胞状接种于80 mL/250 mL三角瓶液体培养基中,置于(25±1)℃、转速110-130 r/min的摇瓶柜中暗培养,培养周期21 d。从第3天起,每隔3 d取样,测定黄芩悬浮细胞鲜重和干重的增长量以及黄芩苷的含量和过氧化物酶活性(各测6组样品,平均值为最终结果)。4条变化趋势曲线横坐标均为生长时间,纵坐标分别为黄芩悬浮细胞鲜重和干重的增长量以及黄芩苷的含量和过氧化物酶活性。
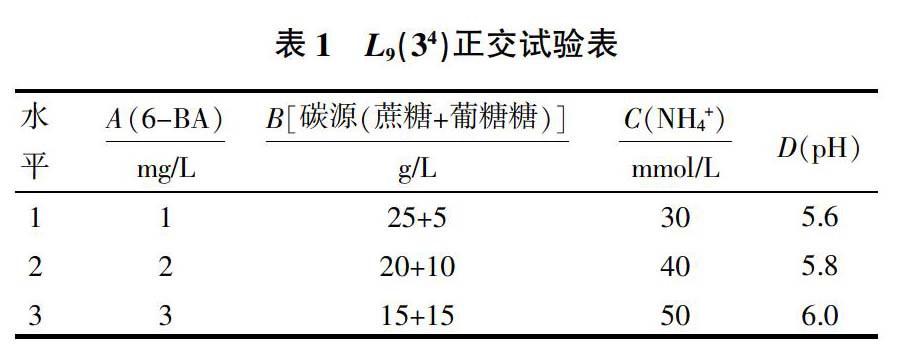
1.2.2正交试验设计 为了研究在黄芩悬浮细胞生长周期中过氧化物酶活性的表达情况,试验采用L(3)正交表设计培养基中6-BA、碳源、氮源、pH的配比(表1)。在不同培养条件下,黄芩悬浮细胞中过氧化物酶的表达情况不同,通过筛选可以得出过氧化物酶表达的最佳培养条件。
1.3项目测定
1.3.1黄芩悬浮细胞黄芩苷含量测定 采用紫外分光光度法。培养悬浮细胞中总黄芩苷含量=培养液中黄芩苷含量+细胞中的黄芩苷含量。量取2 mL的悬浮细胞液,通过抽滤将培养细胞与培养液分离。培养液中黄芩苷含量的测定:将培养液置于60℃烘箱内烘干,加入50%的乙醇10 mL,置于50℃水浴箱内温浴5~6 h,冷却后用50%的乙醇定容。用紫外可见分光光度计测定278 nm处的OD值。细胞中黄芩苷含量的测定:将细胞培养物置于60℃烘箱内烘至恒重,研成粉末,用50目筛网过筛,称取适量粉末,加入50%的乙醇10 mL,置于50℃水浴箱内温浴5~6 h,冷却后用50%的乙醇定容,用蒸馏水稀释、摇匀,作为待检样。用紫外可见分光光度计测定278 nm处的OD值。依据标准曲线方程(图1)计算黄芩苷浓度C。
2.结果与分析
2.1黄芩悬浮细胞的增长曲线和黄芩苷的变化曲线
图2和图3显示,黄芩悬浮细胞的增长曲线呈典型的“s”形,0-6 d为潜伏期,6-18 d为快速合成期,18-21 d为稳定期,黄芩悬浮细胞增长变缓,鲜重最大值达到15.430 6 g,干重最大值达到0.843 7 g。由图4可以看出,黄芩苷在前6d合成比较缓慢,但从培养6 d后合成速率增加,9-15 d为黄芩苷的快速合成期,到18 d时,黄芩苷的积累量达到最大值73.496 2 mg/g。由图2、图3、图4不难发现,黄芩悬浮细胞的增长和黄芩苷合成积累基本上同步,即边增长边合成。
2.2黄芩悬浮细胞中过氧化物酶活性的变化规律
图5显示,在黄芩悬浮细胞生长周期的前6 d,POD低水平表达,活性在低于0.327 U/(g·min)的范围;6-15 d高水平表达,12 d左右POD活性达到最高值1.346 U/(g·min),15-21 d POD低水平表达。
结合图2、图3、图4可以看出,活性变化规律与黄芩悬浮细胞增长和黄芩苷积累量的关系为:在黄芩悬浮细胞生长周期的前6 d,悬浮细胞主要是适应从固体培养到液体培养的环境,这一时期细胞的增长量、黄芩苷合成量和POD活性都不明显:6~15 d,随着细胞适应液体培养的环境,细胞的增长量、黄芩苷合成量和POD活性都迅速增加:15-21 d,由于液体培养基成分的逐渐消耗,所以这一时期细胞的增长量和黄芩苷的合成量开始变得不明显,达到一种稳定的状态,而这时期的POD活性也低水平表达。
通过SPSS Statistics对POD活性与黄芩苷含量的相关性进行分析,得出在黄芩悬浮细胞生长周期前12 d,POD活性与黄岑苷含量的相关系数为0.997,P=0.03<0.01,极显著相关。由此可见,在黄芩愈伤组织生长周期的前12 d,POD活性与黄芩苷的积累相关。
在黄芩悬浮细胞生长周期前12 d,POD的活性与黄芩苷的积累呈正相关性。以POD活性为自变量,黄芩苷含量为因变量,做回归分析。回归方程为y=0.442 1 x+0.868 9,回归系数R2=0.994 3。对回归方程进行方差分析,显著性检测结果见表2。从表2可以看出,概率值P=0.002 832<0.01,说明方程回归极显著。
2.3正交试验结果
从表3可知,最优的培养条件为:MS附加6-BA3 mg/L,碳源15 g/L蔗糖+15 g/L葡萄糖,氮源(NH4+)30 mmol/L,pH 5.6。极差分析结果显示4种不同因素对POD表达都有不同程度的影响,而6-BA在黄芩悬浮细胞培养过程中对黄芩悬浮细胞POD表达有较显著的影响。
3.小结与讨论
植物体中含有大量POD,是活性较高的一种酶。它与呼吸作用、光合作用及生长素的氧化等都有关系,在植物生长发育过程中它的活性不断发生变化。次生代谢过程是各种酶促反应的过程,受酶的调控,酶的表达决定代谢途径的启动及相关特定物质的合成。黄芩苷是细胞次生代谢的产物,它本身可以表征次生代谢的旺盛程度。在本试验中,黄芩悬浮细胞生长周期前6 d为缓冲期,黄芩苷的含量较低,同时POD低水平表达;6-15 d,POD有高水平表达,并达到最高值,而此时也是黄芩苷快速合成期:15~21 d,POD低水平表达,黄芩苷含量的积累变得缓慢,达到一个稳定值。由此可以看出,在黄芩悬浮细胞生长周期(21 d)内,POD活性高低与黄芩苷的积累呈正相关。
结合图4和图5可以看出,POD活性高低与黄芩苷合成的快慢在黄芩悬浮细胞生长周期表现出了同步性,黄芩苷合成速率最快时,POD表达水平也最高。同时,POD活性与细胞的衰老也密不可分,POD活性越强,细胞衰老的速度也就越快,即POD活性也是细胞生长周期长短的一个制约因素。次生代谢产物的积累主要在植物生命的后期。可以看出POD是影响黄芩悬浮细胞成长和黄芩苷积累的一个重要因素,因此POD的活性对黄芩悬浮细胞生长和黄芩苷积累有着重要意义。影响植物次生代谢产物积累的因素很多,既有内因又有外因。通过对影响POD积累因子的研究,初步掌握了一些影响植物次生代谢产物积累的规律,对生产实践有一定的指导作用,但这些规律又因具体植物的不同而有差异。因此,如何将这些规律进行推广还需要深入研究。
猜你喜欢
黑龙江大学自然科学学报(2022年4期)2022-11-17
河北科技师范学院学报(2022年2期)2022-08-26
现代畜牧科技(2021年8期)2021-10-13
今日农业(2020年16期)2020-12-14
基层中医药(2020年7期)2020-09-11
现代园艺(2020年9期)2020-05-24
现代园艺(2019年1期)2019-02-18
南方农业学报(2018年12期)2018-09-10
中成药(2017年12期)2018-01-19
安徽农学通报(2017年12期)2017-07-13
