
转Bt基因油菜的抗虫性分析
2016-05-14 06:42万丽丽王转茸辛强董发明洪登峰杨光圣
湖北农业科学 2016年9期
万丽丽 王转茸 辛强 董发明 洪登峰 杨光圣

摘要:利用农杆菌介导的遗传转化分别将Cry1C和Cry2A的2个单价Bt基因转入油菜(Brasica napusL.),以两个纯合的转基因抗虫油菜为亲本,通过有性杂交的方法将不同Bt基因聚合,培育双价Bt抗虫油菜,并对其抗虫性和种子品质性状进行评价。结果表明,Cry1C、Cry2A在聚合后均能稳定表达,和单价Cry1C转基因植株相比,双价株系中蛋白质含量明显降低,以Cry2A为母本的聚合株系蛋白质含量降低更多,Cry1C在遗传上存在母本效应。室内接种小菜蛾二龄幼虫结果显示,转化单价和双价聚合Bt基因的抗虫性增强,其中转化单价Cry1C的抗虫性优于双价聚合Bt基因和单价Cry2A基因的转基因植株。玻璃温室栽培转基因植株,单价Bt和双价聚合Bt基因的转基因植株能生存而非转基因植株受到严重虫害而死亡。对抗性优良的单价和双价聚合的转基因植株种子品质测定发现,与未受到虫害的受体材料双低优良恢复系7-5的含油量和硫苷含量差异不显著,从而达到抗性改良的目的。
关键词:油菜(Brassica napus L.);苏云金芽孢杆菌;Cry1C;Cry2A;双价聚合;农杆菌介导遗传转化;品质性状
中图分类号:Q785 文献标识码:A 文章编号:0439-8114(2016)09-2386-06
油菜(Brassica napus L.)在生长发育过程中容易受到虫害的侵袭,有10多种害虫在油菜生产上能大面积发生并造成严重减产。近年来,气候变化使得虫害日趋严重。对农业生产造成损失。尽管使用化学农药能够取得良好的防控效果,但是农药残留对环境和人体的健康产生的危害已引起广泛的关注。利用转基因技术将抗虫基因转入油菜中,可以一定程度上改良油菜的抗虫性。转基因抗虫植物最重要的基因来源于微生物的抗虫基因,即苏云金芽孢杆菌杀虫基因(Bt)。Bt杀虫晶体被昆虫摄入后,经历了晶体蛋白溶解、原毒索酶解活化、与中肠受体结合、不可逆插入膜内、形成离子通道五个阶段最后实现杀虫效果。根据杀虫晶体蛋白的杀虫特异性以及氨基酸序列的同源性标准将发现的42种杀虫蛋白基因分为7大类。其中类型Ⅰ(CryⅠ)抗鳞翅目(Lepidoptera)昆虫,类型Ⅱ(CryⅡ)抗鳞翅目和双翅目(Diptera)昆虫,类型Ⅲ(CryⅢ)抗鞘翅目(Coleoptera)昆虫,类型Ⅳ(CryⅣ)抗双翅目昆虫,CryⅤ对鳞翅目和鞘翅目以及线虫特异,CryⅥ对线虫特异,这六类统称为晶体蛋白基因家族(Crvstalprotein gene,cry)。CrtA对双翅目特异,与Cry基因完全不同,属于细胞外毒索(Cvtolvtic protein,Cyt)。Bt产生的晶体蛋白对鳞翅目昆虫具有特异的毒性,作为一种生物杀虫剂在农业害虫防治中广泛应用。通过人工修饰Bt基因,如位点特异性突变的Crv3Bbl毒蛋白在玉米中表达可提高其对根虫的抗性。1990年Monsanto公司首次在转Bt抗虫棉中获得人工修饰的CryIA(6)和CryIA(c)基因,这2个基因在植株中蛋白含量占总可溶性蛋白的0.05%-0.10%。Cry基因与植物基因密码子上的差异导致转入植物体内转录过程形成二级结构,mRNA的稳定性下降以及翻译效率降低,来自苏云金芽孢杆菌的5个δ-内毒素Cry1Ab、Cry1Ac、Cry1C、Cry2A和Cry9C能够与蛀茎虫三化螟和二化螟中肠不同受体位点结合,利用毒性蛋白与刷状缘膜囊结合能力竞争性试验分析得出不同Bt基因抗虫效果的差异。华中农业大学水稻课题组人工合成Bt基因Cry1Ac、Cry2A、Cry1C和Cry9C转入优良水稻恢复系明恢63中,获得对两种主要水稻钻蛀害虫三化螟和二化螟抗性优良的转基因家系。对苏云金芽孢杆菌的Cry1Cal进行截短,依据单子叶植物偏好来提高Bt基因的GC含量转化到粳稻品种秀水11中能提高转基因水稻对斜纹夜蛾和钻蛀虫的抗性。
随着Bt作物的普遍商业化种植给目标昆虫群体造成了极大的选择压,产生了昆虫克服抗性的巨大风险。小菜蛾(Plutella xylostella)是迁飞害虫,主要危害十字花科植物,对几乎所有类型的化学杀虫剂都产生过抗性。在1990年田间发现了对Bt蛋白产生抗性的小菜蛾,是首次在自然条件下发现对Bt杀虫蛋白产生抗性的昆虫,是长期使用Bt制剂的产物,可以利用不同Bt基因聚合延缓昆虫抗性。Tu等2000年利用水稻ActinⅠ启动子驱动两个Bt基因cry1A(6)和cry1A(c)基因融合表达载体转化明恢63恢复系和杂交种汕优63。Bt汕优63在自然条件以及室内接种水稻钻蛀害虫三化螟表现出很好的抗性,并且不会带来产量损失。
本研究采用农杆菌介导法,以Bar基因作为抗性筛选标记,将华中农业大学水稻课题组人工合成的Bt基因Cry2A和Cry1C转入到甘蓝型油菜Pol-CMS优良恢复系材料7-5中,获得抗虫性改良的转基因家系。另外。将分别转化2个基因的植株杂交聚合,对杂交后纯合转基因家系进行抗虫性评价,以期为转Bt基因油菜的抗虫性研究提供参考。
1 材料与方法
1.1 材料
供试的甘蓝型油菜为华中农业大学油菜室所提供的优良PolCM双低恢复系7-5。选取干净饱满油菜种子,75%乙醇消毒1min,0.1%HgCl2溶液表面灭菌15min,无菌水冲洗3次,置于MS固体培养基上发芽。取生长6d的无菌苗下胚轴,用于试验材料。植物表达载体pCAMBIA-Ubiquitin promoter-Cry1C和oCAMBIA-Ubiquitin promoter-Cry2A为华中农业大学水稻转化课题组林拥军教授提供。大肠杆菌Top10和根癌农杆菌GV3101由华中农业大学作物遗传改良国家重点实验室油菜课题组保存。DNA聚合酶购自Promega公司,DNA marker购自Takara公司。田间喷施Basta购自Bayer公司。
1.2 方法
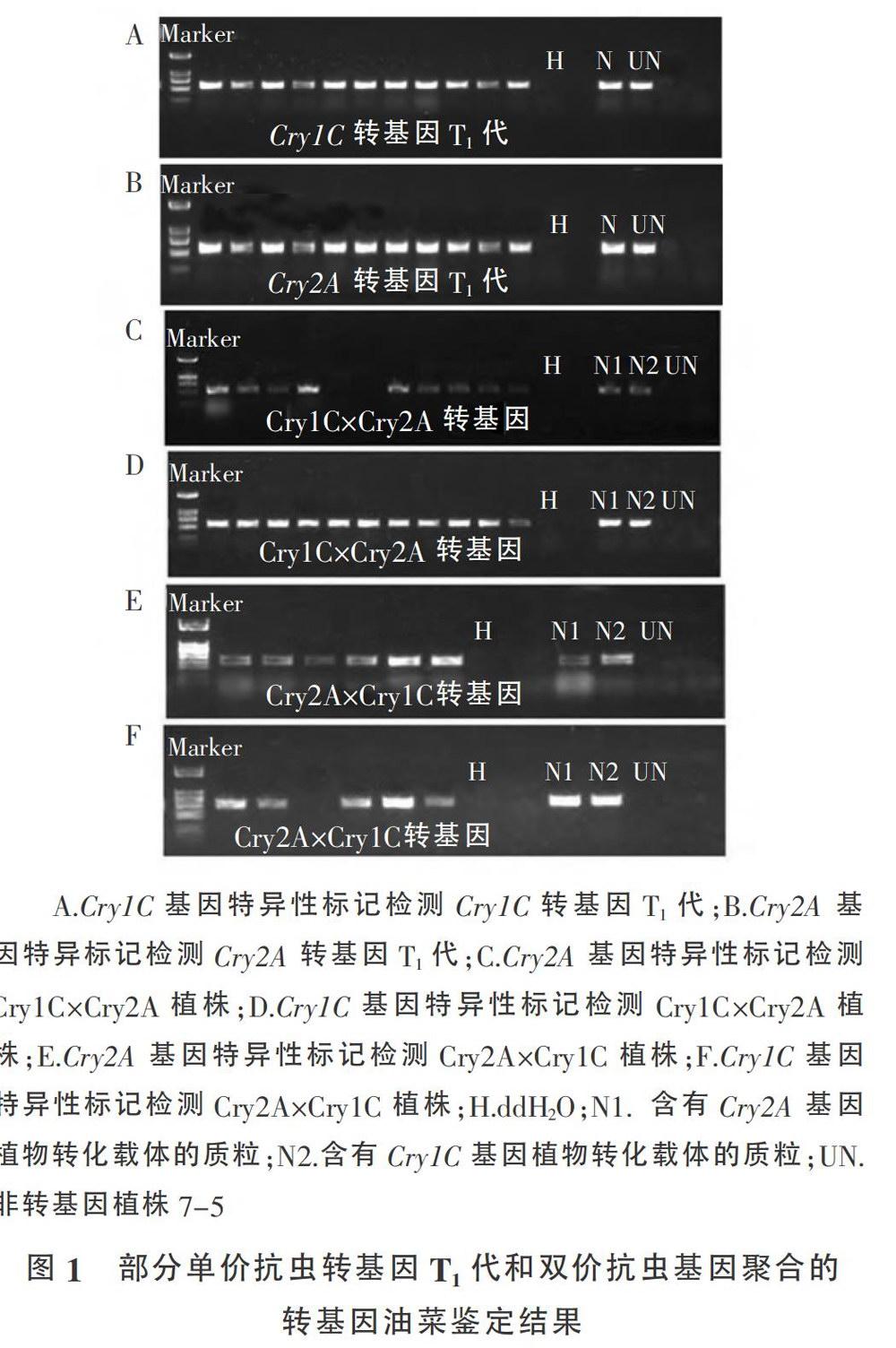
1.2.1 转基因植株鉴定 采用农杆菌介导的遗传转化方法,以改造的pCAMBIA质粒作为载体,抗除草剂基因膦丝菌素乙酰转移酶(Bar)基因作为选择标记基因,以Cry1C和Cry2A分别转化优良的油菜恢复系7-5。农杆菌菌株为GV3101,转基因植物检测的引物Crv2A-L(CGTGTCAATGCTGACCTGAT)、Cry2A-R(GATGCCGGACAGGATGTAGT)、Cry1C-L(TTCTACTGGGGAGGACGTCG)、Cry1C-R(CGGTATCTTTGGGTGATTGG)、Bar/L(GCTCAACACATGAGCGAAAC)、Bar-R(CGCACAATCCCACTATCCTF)。转基因植株田间喷施除草剂Basta的浓度为500mg/L,苗期每7d喷施1次,连续21d。 1.2.2 转基因植株Bt蛋白含量的测定 转基因植株叶片中Bt蛋白浓度测定采用EnvironLogix公司(EnvironLogix。Portland,USA)的Bt蛋白ELISA检测试剂盒,CryIC蛋白的测定使用了EnvironLogix公司ELISA检测试剂盒QualiPlateTMKit fo rCry1C,具体操作参考试剂盒说明。
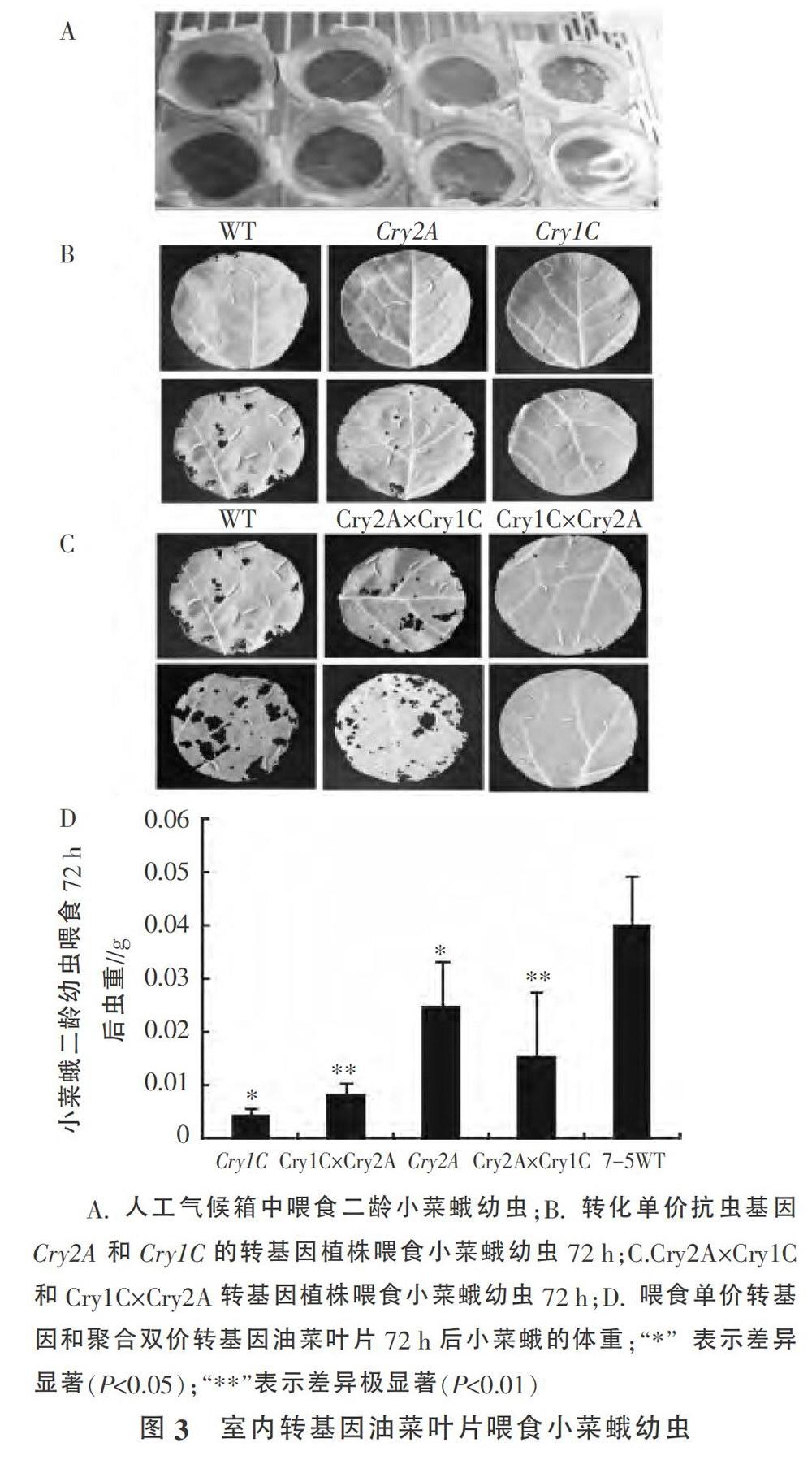
1.2.3 小菜蛾二龄幼虫室内和人工玻璃温室中的抗性测定 待田间油菜生长至5-6叶期时,采集新鲜的叶片,剪成直径为6(3m的圆盘。将圆盘放置于铺有卫生纸的培养皿上,将10头二龄幼虫接种到每片叶子上,将接种材料置于人工气候箱中。培养温度25℃,湿度80%,光照3000lx,光周期16h光照/8h黑暗。检测转基因植株抗虫效果,每个处理重复3次。记录幼虫取食72h后的平均重量、致死率和生长状况。转基因植株种植在人工玻璃温室中,苗期观察植株被害虫取食的情况。
2 结果与分析
2.1 转基因植株的获得和鉴定
通过转基因技术分别获得了转化Cry1C和Cry2A的植株,根据基因特异序列设计引物检测单价抗虫转基因T。代植株,确定阳性转基因植株。在田间将转化不同单价抗虫基因的转基因植株正反交,其中获得Cry1C×Cry2A(以Cry1C转基因植株为母本,Cry2A转基因植株为父本的杂交后代)和Cry2A×Cry1C(以Cry2A为母本。Cry1C为父本的杂交后代)植株,对其进行抗虫基因的特异性标记分析(图1)。将所获得的转基因植株种植在大田,用Basta喷施鉴定后发现阳性的转基因植株都具有除草剂抗性(图2)。
2.2 转基因株系室内抗虫性评价
选取单价抗虫转基因和聚合双价抗虫转基因油菜植株的叶片喂食二龄小菜蛾幼虫(图3A、B、C),72h后统计喂食幼虫的虫重,结果表明,与野生型7-5相比较,转化单价Cry2A基因的转基因油菜植株喂食后虫重下降,差异显著。而转化单价Cry1C基因以及双价抗虫基因聚合后的转基因油菜植株叶片喂食后,小菜蛾幼虫虫重出现了显著或者极显著下降,进一步证明了转化抗虫基因能够有效地提高油菜抗小菜蛾幼虫能力(图3D、表1)。
由于转化Cry1C基因的转基因植株的抗虫性表现极显著优于对照,选取单价转化Cry1C的T1代转基因3个家系共21个植株的叶片和双价聚合转基因植株6个家系共38个植株的叶片,测定其Cry1C蛋白的含量,结果显示在转化Cry1C基因的转基因植株中Cry1C蛋白平均含量高于聚合Cry1C和Cry2A基因的转基因植株中的蛋白含量(图4)。
2.3 转基因株系在玻璃温室中的抗虫性
将转化单价抗虫基因和双价抗虫基因的转基因油菜植株于4月底种植在人工玻璃温室中,观察虫害对转基因幼苗的影响(图5),可见转化单价Cry1C基因的转基因植株抗虫性表现最优。
2.4 转基因株系种子品质评价
抗虫转基因油菜遇到虫害时自身会启动一些防御反应,同时植物会消耗自身的能量从而导致产量和品质的下降。转基因受体材料7-5是优良双低恢复系油菜,抗虫基因Cry1C和Cry2A在泛素启动子作用下在7-5材料中组成型表达,一定程度上会使植株产生代谢负荷,油菜子的品质受到影响。因此,对单价和双价聚合的转基因植株种子品质进行测定(表2),与对照7-5相比较,Cry1C×Cry2A聚合转基因家系油酸C18:1含量显著增加,二十碳烯酸C20:1含量极显著下降。Cry1C转基因T1代两个家系二十碳烯酸C20:1含量极显著高于7-5。Cry2A转基因T1代家系二十碳烯酸C20:1含量显著高于7-5。对种子中含油量和硫苷进行分析得出,所有的转基因家系与7-5对照相比较差异不显著(图6)。
3 讨论
3.1 Bt基因在转基因后代中的遗传
为了获得优良抗虫表型的转基因家系,对抗虫表现不一致的T0代转基因植株的选择是育种过程中的必要步骤。本试验所获得的T1代都来源于转基因插入位点为一个拷贝并且抗虫表现优于转基因受体对照7-5的T0代植株。对T1代进一步的选择按照以下原则:①单价Bt基因稳定遗传的纯合株系:②抗虫表型稳定有效:③与7-5相比没有明显的表型变异,产量和品质不受影响。将纯合的单价抗虫Cry1C和Cry2A转基因植株杂交获得双价转基因油菜,其中以Cry1C为母本获得的聚合油菜抗虫性优于以Cry2A为母本获得的聚合油菜,这一结果与单价抗虫Cry1C基因转化所得的抗性高于转化Cry2A基因植株的抗虫性相一致,证明了Cry1C转基因植株作为母本传递核物质有着母体效应。
3.2 Bt基因聚合的重要性以及有性杂交聚合的优势
本试验在甘蓝型油菜7-5中导入两个Bt基因,若2种毒素能够识别有差异的受体,那么昆虫对他们同时产生抗性的几率会降低。因为2种与昆虫类受体结合的毒蛋白同时失效的可能性不大,双价Bt抗虫转基因植株能够有效地延迟抗性失效的时间。在大田环境下,油菜容易遭受多种虫害危害,聚合多个Bt基因能够产生很好的防控作用,同时可以有效延缓抗性品种的应用时间。在美国和澳大利亚等国家主要采用高剂量和庇护所同时种植,即大田环境下种植Bt作物的同时需要种植一些非转基因的植株来维持一定数量的敏感昆虫个体。那么双价Bt作物所需要的庇护所比例较小,可以将经济损失降到很低。但是中国的种植制度、农户规模以及靶标害虫等社会和生态环境不同,这种高剂量庇护所策略在中国推广实施较困难,因而通过双价或者多价基因聚合更符合国情需求。
在本试验中通过有性杂交的方式进行基因聚合,首先分别获得纯合的单价Bt转基因家系,将它们杂交后再次选择纯合株系。这个过程中需要对两个随机插入到基因组的Bt基因同时进行标记筛选,经历的时间长工作量大。但这种方法在实际育种材料的改良应用中存在着一定优势,首先将现有的多个育种材料作为受体进行单价Bt基因转化,获得变异小抗性和农艺性状稳定优良的家系,之后可以根据不同育种需求将含有单价Bt基因的材料自由组配,实现父母本家系的优势互补,改良产量、品质等性状,从而提高获得理想材料的机会,这样可以避免对每个组合进行双价或者多价基因的遗传转化。
3.3 双价Bt抗虫转基因叶片中Cry1C蛋白含量与单价Cry1C蛋白的差异
本试验中单价抗虫基因Cry1C和Cry2A都是在来源于玉米的组成型启动子Ubiquitin promoter作用下实现表达的。尽管这两个Bt基因在序列上没有极高的相似性,但是二者聚合后由于启动子的完全同源会引起基因沉默导致抗虫基因表大量下降。对Cry1C蛋白含量检测发现,双价转基因家系中蛋白浓度普遍低于单价Cry1C转基因植株。在实际应用中采用不同的启动子,如CaMV35S和Actin作用聚合不同的Bt基因表达可以降低聚合后基因表达量下降以及基因沉默失效的机率。另外,组成型表达的启动子在植物的各个器官表达势必造成能量的消耗而产生代谢损耗,并且种子中Bt蛋白的含量高会引发消费者对转基因食品安全的质疑。因此,利用组织特异启动子,如水稻rbcS(1,5-二磷酸羧化酶或者加氧酶的小亚基)启动子驱动Bt抗虫基因在叶片、茎秆组织中特异表达可以提高抗虫的有效性。Bt基因持久高效的表达会影响植株的其余性状的变异,如株高、育性等。增加对田间昆虫的选择压。加速昆虫的抗性变异,最终导致植物抗性丧失。因此。选择农艺和品质性状变异少。抗虫性优良而Bt蛋白含量较低的转基因家系应用于大田可以有效地避免上述问题,
猜你喜欢
中国棉花(2023年4期)2023-08-08
小学生学习指导(低年级)(2021年3期)2021-07-21
数学小灵通·3-4年级(2021年6期)2021-07-16
林业科技(2020年3期)2021-01-21
小学生学习指导(低年级)(2019年6期)2019-07-22
中国蔬菜(2016年8期)2017-01-15
茶叶通讯(2014年4期)2014-02-27
