
7个葡萄砧木根系的抗寒性研究
2016-05-12 09:25:10钟海霞艾尔买克才卡斯木张付春潘明启
新疆农业科学 2016年3期
钟海霞,艾尔买克·才卡斯木,张付春,潘明启,
韩守安,张 雯,谢 辉,陈 锐,伍新宇
(新疆农业科学院园艺作物研究所/农业部新疆地区果树科学观测试验站,乌鲁木齐 830091)
7个葡萄砧木根系的抗寒性研究
钟海霞,艾尔买克·才卡斯木,张付春,潘明启,
韩守安,张 雯,谢 辉,陈 锐,伍新宇
(新疆农业科学院园艺作物研究所/农业部新疆地区果树科学观测试验站,乌鲁木齐830091)
摘要:【目的】筛选出抗寒性较强的葡萄砧木品种,为新疆引进优良抗寒砧木及推广栽培提供科学依据。【方法】以新疆7个引种葡萄砧木的1年生扦插苗根系为材料,对7个低温处理下(-2、-4、-6、-8、-10和-12℃)根系的相对电导率、可溶性糖、游离脯氨酸、MDA含量和可溶性蛋白等理化指标进行测定,采用隶属函数法分析各理化指标,进行各品种抗寒性比较7个品种根系的抗寒性强弱。【结果】随着胁迫温度的逐渐降低,各品种根系的相对电导率、可溶性糖、游离脯氨酸和MDA含量逐渐升高,可溶性蛋白含量逐渐增加,但升高速度及增加幅度略有不同。隶属函数分析综合得出其抗寒性为:SO4<5BB<188-08<101-14<110R<5C<脱毒贝达。【结论】7个引种砧木中根系抗寒性最强的为脱毒贝达,能抵御的最低临界温度为-12℃;110R、5C、101-14、188-08和5BB抗寒能力中等,能抵御-10℃低温;根系抗寒性最弱的为SO4,能抵御临界最低温度为-8℃。
关键词:葡萄砧木;根系;抗寒性
0引 言
【研究意义】葡萄是新疆的主要果树树种之一,但新疆冬季寒冷气候大大制约了葡萄的大规模生产。葡萄根系冻害在南、北疆均有发生,其发生不利于树体生长,严重时可导致树体死亡,影响葡萄的产量和品质,同时影响了果农的收入和产业的发展[1]。利用抗寒砧木不但能提高葡萄的抗寒性,而且对提高产量和改善果实品质具有重要作用。根系作为葡萄抗寒能力最弱的器官,其抗寒性强弱决定了葡萄是否能安全越冬,研究葡萄砧木根系的抗寒性,并进行测定评价,对新疆葡萄栽培选择砧木有实际意义。【前人研究进展】张倩等[2]对5个种群的葡萄根系抗寒性研究发现,美洲种葡萄抗寒性最强,山欧杂种、法美杂种和欧美杂种抗寒性居中,欧亚种葡萄抗寒性最弱。其中美洲种的‘康可’抗寒能力最强,欧亚种的‘红地球’、‘无核白鸡心’抗寒性较弱;周香云等[3]对8种葡萄砧木根系进行了抗寒性研究发现随低温胁迫程度的加重,根段浸提液相对电导率呈“S”形曲线上升,根据求算出的低温胁迫半致死温度,初步判定其根系的抗寒能力大小为:140R<110R<520A<5BB 1材料与方法 1.1材 料 供试材料为7个从河北引种的葡萄砧木贝达(经过脱毒)、110R、5C、188-08、SO4、101-14、5BB的1a生扦插苗根系,在园艺所葡萄基地核心示范园种植,葡萄埋土前(2013年11月5日)采集。 1.2方 法 各砧木各选15株,取粗度直径2~3mm的根系,用蒸馏水洗净,去掉须根。放入高低温冰箱中处理,试验处理温度设-2℃(对照)、-4、-6、-8、-10和-12℃6个温度梯度处理,降温速度设4℃/h,至各低温处理温度后持续12h,再缓慢升温至0℃放置8h,室温放置8h,然后放置-2℃冰箱测定相关指标。每处理3次重复。 电解质渗出率采用电导仪(DDS-12A型)测定;游离脯氨酸含量采用酸性茚三酮显色法测定[8];可溶性糖采用蒽酮显色法测定;可溶性蛋白采用G-250考马斯亮蓝法测定;丙二醛(MDA)含量用硫代巴比妥酸法测定[9]。 采用Fuzzy数学中的隶属函数法[10]对各项抗寒理化指标进行综合评价,与抗寒性呈正相关的游离脯氨酸、可溶性糖、可溶性蛋白用以下公式:Uij=(Xij-Xjmin)/(Xjmax-Xjmin);与抗寒性呈负相关的相对电导率、MDA含量用以下公式:Uij=1-(Xij-Xjmin)/(Xjmax-Xjmin)。Uij为i砧木j指标的隶属函数值;Xij为i砧木j指标的测定值;Xjmax和Xjmin分别为7种砧木6个温度处理中j指标的最大值和最小值。对各指标隶属函数值的平均值比较,综合排名即各砧木抗寒性的强弱。 1.3数据统计 采用Excel 2010和SPSS 17.0软件进行数据统计及分析。 2结果与分析 2.1不同低温处理对根系相对电导率的影响 研究表明,各砧木品种的根系相对电导率呈现出明显的“S”型曲线,均随温度的逐渐降低而上升,对根系细胞膜的伤害程度也不断加重,表明相对电导率与砧木抗寒性之间存在密切的关系。在-2(CK)~-4℃时,相对电导率略有下降趋势;当温度继续下降(-4~-10℃)时,相对电导率急剧上升;低温处理至-6℃时7个砧木品种根系的相对电导率差异不显著(P<0.05),但较-4℃略有增加,这说明根系的细胞膜有自我调节的作用。温度从-6℃往下降时,相对电导率开始升高,-8℃时,除SO4的相对电导率达53.83%,超过了50%之外,其余品种均处于50%以下,说明此时已对SO4根系造成伤害。在-6~-8℃时SO4、101-14的相对电导率变化幅度较大,分别增加了27.37%、19.11%。当温度下降至-10℃时,除脱毒贝达的相对电导率为48.62%,不到50%外,其余品种的相对电导率均超过50%,其中SO4最大,为78.18%,这说明此时除脱毒贝达之外,其余6个砧木品种根系均受到了不同程度的伤害。在-8~-10℃时110R、5BB、5C、188-08的相对电导率变化幅度较大,分别增加了26%、25.67%、23.04%、22.02%;-10~-12℃所有品种相对电导率均超过了50%,其中SO4最大,为89.47%,比相对电导率最小的脱毒贝达高了28.5%。在-10~-12℃时脱毒贝达的相对电导率变化幅度最大,增加了21%,-12℃之后各品种缓慢上升逐渐趋于平衡,这说明-12℃已对砧木根系的细胞膜造成了不可逆的伤害,严重的甚至丧失了细胞膜透性,从而导致细胞死亡,大部分组织和器官遭受冻害。图1 图1低温胁迫下葡萄砧木根系相对电导率变化 2.2不同低温处理下根系游离脯氨酸含量变化 研究表明,各砧木品种根系的游离脯氨酸含量随温度的逐渐降低呈现先缓慢上升后迅速升高。在-2℃(CK)~-10℃时,游离脯氨酸含量缓慢上升,各砧木品种根系的相对电导率差异不显著(P<0.05)。当温度继续下降-12℃时,游离脯氨酸含量急剧上升。在-10~-12℃时游离脯氨酸含量变化幅度最大的是SO4,增加了42.64%;其次是101-14、188-08、5C、脱毒贝达、110R,分别增加了23.6%、23.06%、22.18%、21.61%、19.59%。变化幅度最小的是5BB,增加了11.72%,这说明各砧木品种对低温产生了积极响应。图2 2.3不同低温处理对根系丙二醛(MDA)含量的影响 在低温逆境下,果树发生膜脂过氧化作用后会产生丙二醛(MDA)积累物质,即为低温胁迫下抗寒性的鉴定指标之一。研究表明,7个砧木品种根系的MDA含量呈现出明显的 “S”型曲线,均随温度的逐渐下降呈现出先降后升的趋势,可以看出MDA含量与砧木根系的抗寒性存在着密切的关系。从-2℃(CK)~-4℃时,各品种根系的MDA含量有下降趋势,但其下降幅度存在差异。SO4和5BB下降幅度最大,分别下降了14%和13.6%。110R下降幅度最小,为3.9%。在-4~-10℃时,各品种根系的MDA含量迅速上升,其中上升趋势最明显的是SO4,最不明显的是脱毒贝达。当温度持续降至-12℃时,各品种的MDA含量缓慢上升,且趋于平衡状态。总体看来,整个低温胁迫过程中,SO4根系的MDA含量始终处于较高水平,其次是5BB、188~08和101~14等,脱毒贝达根系的MDA含量始终处于较低水平,这说明脱毒贝达根系细胞膜遭受损害的程度最小,膜透性也最小,而受伤害最大的为SO4,这说明SO4根系的抗寒性最弱。图3 图2低温胁迫下葡萄砧木根系游离脯氨酸含量变化 图3低温胁迫下葡萄砧木根系MDA含量变化 2.4不同低温处理对根系可溶性糖含量的影响 低温逆境中果树体内会积累大量的可溶性糖,它与抗寒性呈正相关,即抗寒性强的树体,可溶性糖积累量较大。研究表明,不同品种砧木的根系抗寒能力不同。随着温度的逐渐降低,7个葡萄砧木根系的可溶性糖含量呈逐渐升高趋势,即温度越低,可溶性糖含量越大。-2℃(CK)~-4℃时,各品种间可溶性糖含量无显著或极显著性差异。-4~-8℃时,各品种根系的可溶性糖含量迅速上升,各品种间存在显著差异(P﹥0.05),其中5C、脱毒贝达、101-14上升幅度较大,分别上升了2.38%、2.24%、2.10%。-8~-10℃时,各品种的可溶性糖含量略有增加,至-12℃时又迅速增加,这说明各品种根系已遭受到不同程度的伤害,此时110R、5C和脱毒贝达增加幅度较大,分别较-10℃时的可溶性糖含量增加了4.68%、3.97%和3.03%。从温度下降的整个过程来看,各品种中脱毒贝达根系的可溶性糖含量一直处于较高水平,其次是5C、110R、101-14和188-08,而5BB和SO4的可溶性糖含量在整个低温胁迫中处于较低水平。图4 图4低温胁迫下葡萄砧木根系可溶性糖变化 2.5不同低温处理对根系可溶性蛋白含量的影响 在低温逆境中树体可溶性蛋白含量会有所积累,对果树的抗寒性有一定的指示作用。低温胁迫对葡萄砧木根系可溶性蛋白的影响可以看出,不同低温处理下,各品种根系的可溶性蛋白含量有所不同。在-2~-4℃时,脱毒贝达、101-14、5C和110R呈下降状态,其中脱毒贝达下降幅度最大,下降了1.01%;188-08和5BB上升幅度最大,上升了0.32%。在-4~-12℃时,各品种根系的可溶性蛋白含量逐渐上升,品种之间存在显著差异。在整个低温下降过程,各品种中SO4始终处于最低状态,脱毒贝达始终处于最高状态。图5 图5低温胁迫下葡萄砧木根系可溶性蛋白变化 2.67个葡萄砧木抗寒性的综合评判 采用隶属函数法得出各理化指标的平均隶属度,平均隶属度的值越大,抗寒能力越强,反之抗寒能力越弱,抗寒位次数字越小表示抗寒性越强。研究表明,脱毒贝达的隶属函数值为最大(0.531),抗寒位次为1,说明脱毒贝达的抗寒性最强。5C、101-14、5BB、110R和188-08的隶属函数值位于中间,SO4的隶属函数值是0.466,为最小,抗寒位次最大(7),这说明7个砧木中SO4抗寒能力最弱。根据平均隶属度及抗寒位次来看, 7个引种砧木的抗寒性强弱顺序是:SO4< 5BB<188-08<101-14<110R<5C<脱毒贝达。表1 表1低温胁迫下7种葡萄砧木抗寒指标的隶属度值Table 1 Subordination value of cold resistance indicator of seven grape rootstock under cold stress 品种Varieties相对电导率Relativeelectiveconductivity游离脯氨酸Freeproline丙二醛MDA可溶性糖Solublesugar可溶性蛋白Solubleprotein平均隶属度AverageofSF抗寒位次Rateofcoldresistance脱毒贝达0.6780.3050.6120.4040.6820.5361101-140.5980.3090.5540.3700.5810.4834188-080.6290.4160.5480.3580.4520.4805SO40.5810.2600.5140.3430.6290.46675BB0.6100.4470.5350.4220.3770.47865C0.6530.3710.6520.3420.5930.5222110R0.6130.3190.6050.3200.6090.4933 注:表中抗寒位次数字小表示抗寒性强 Note: The smaller number of rate of cold resistance represents the stronger cold resistance 3讨 论 逆境胁迫下果树体内各种渗透调节物质大量积累,赋予多种树体渗透调节的能力。在低温下细胞溶质的主动积累产生的细胞渗透势的下降是渗透调节的关键[11]。树体细胞膜的抗性强弱取决于可溶性糖、可溶性蛋白质、游离脯氨酸等物质的积累,并通过其代谢调节,维持胞内的平衡。葡萄根系的抗寒能力是葡萄最弱的器官,低温冻害时根系最先遭到伤害。植物受低温胁迫后其电导率会有所增大,电解质出现大量外渗现象。低温达到一定程度后,生物膜的膜脂由液晶态转变为凝胶态;细胞膜的透性会增大,膜内和膜外物质发生自由扩散现象,即电解质外渗,其渗透量反映了细胞膜遭受伤害的程度。可用电导度的增加值来反应细胞质膜受伤害的程度。因细胞外渗物质的量与相对电导率成正比,相对电导率越高,质膜遭受冻害的程度则越大,树体遭受低温伤害则越严重。因此可用相对电导率的大小来反应抗寒性的强弱。抗寒能力强的膜透性变化幅度较小,反之亦然。实验中,不同低温逆境中贝达砧木的抗寒性最强,其电解质渗出率变化幅度最小;SO4的抗寒性较弱,电解质渗出率的变化幅度最大,其它5个品种居中。 游离脯氨酸可增强蛋白质的水合作用,对细胞结构、细胞运输和调节渗透压等具有维持平衡作用,避免水分流失而使细胞遭受伤害,使植物具有一定抗逆性。游离脯氨酸含量的大小与果树的抗寒性之间有密切关系。实验研究表明随处理温度的降低各品种根系的游离脯氨酸含量也逐渐积累和升高,对低温做出了积极的响应。这与王燕凌等[12]研究认为3个葡萄在低温环境中游离脯氨酸含量随温度的降低而逐渐升高的结果相一致。在温度下降过程中,游离脯氨酸含量变化幅度最大的是SO4,其次是101-14、5C和脱毒贝达等,变化幅度最小的是5BB。 果树在低温逆境中体内产生的自由基不断积累对细胞会造成不同程度的伤害。细胞膜系统遭受伤害,发生膜脂过氧化会不同程度地损害细胞膜的结构、功能,不利于细胞的存活,膜脂过氧化的产物是丙二醛(MDA),其积累和扩散不利于树体内反应。低温下MDA含量的变化可反映出细胞膜系统遭受的伤害,从而说明树体抗寒性强弱。研究结果表明7个砧木品种根系的MDA含量随温度的逐渐降低呈现出先降后升的趋势。低温胁迫过程中,根系MDA含量最高的是SO4,其次是5BB、188-08和101-14等,含量最低的是脱毒贝达根系,反映出根系细胞膜遭受损害程度最大的为SO4,而细胞膜透性及膜遭受损害最小的是脱毒贝达,即SO4根系的抗寒性最弱,5BB、188-08和101-14根系的抗寒性中等,贝达根系的抗寒性最强,这与高登涛等[13]对引入石河子地区的5个葡萄砧木抗寒性比较得出砧木‘贝达’和‘5BB’抗寒性最强结果基本一致。 可溶性糖是冷害或冻害下植物体内重要的渗透调节物质。糖分积累可使细胞渗透压增强,提高细胞内溶质的浓度,降低溶液冰点,缓冲胞质过度的脱水,避免胞质胶体遇冷后凝固,降低对细胞的伤害,从而增强其抗寒能力[14]。抗寒性强的品种可溶性糖含量高于抗寒性差的品种[15]。研究发现整个低温处理后,脱毒贝达根系的可溶性糖含量最高,抗寒性最强;5BB和SO4的可溶性糖含量处于较低水平,其抗寒性相对最弱,这与鲁金星等[16]研究结果相一致。可溶性蛋白可增强细胞持水力,对细胞具有保护的作用,可增强低温对植物抵御冻害的能力,其含量的增加与树体的抗寒能力存在正相关关系[17]。研究结果表明在整个低温下降过程,7个砧木品种中SO4根系的可溶性蛋白含量为最小,脱毒贝达根系的可溶性蛋白含量为最大。 研究利用隶属函数法对新疆7个引种葡萄砧木根系的各抗寒生理生化指标进行了综合评判,结果表明抗寒能力最强的砧木品种为脱毒贝达,根系能抵御-12℃低温;抗寒能力较强的品种为5C、110R、101-14、188-08和5BB,其根系能抵御-10℃低温;抗寒能力最弱的品种为SO4其根系能抵御-8℃低温。实验结果得出的7个引种品种中脱毒贝达抗寒能力虽最强,但生产中利用其进行砧木嫁接易出现黄化现象影响了葡萄生长及产业发展。110R、101-14、5BB等根系抗寒性虽比脱毒贝达弱,但在新疆不易出现黄化问题,且生长适宜性较好,对新疆砧木的推广及利用具有重要作用。 4结 论 对引进新疆的7个葡萄砧木品种(脱毒贝达、110R、5C、188-08、SO4、101-14和5BB)根系的抗寒性进行理化指标检测,并结合隶属函数法综合评价各砧木品种根系的抗寒性强弱,新疆7个引种砧木中根系抗寒性最强的为脱毒贝达;能抵御的最低临界温度为-12℃;5C、110R、101-14、188-08、5BB的根系抗寒能力中等,能抵御-10℃低温;根系抗寒性最弱的为SO4,能抵御最低临界温度为-8℃。 参考文献(References) [1]樊丁宇,卢春生,闫鹏,等. 南疆冬季不同条件下果园微环境的温度差异[J]. 果树学报,2013,30(6): 989-993. FAN Ding-yu,LU Chun-sheng,YAN Peng,et al. (2013). Preliminary study on temperature difference of orchard micro-environment in winter under different conditions in Southern Xinjiang [J].JournalofFruitScience,30(6): 989-993. (in Chinese) [2] 张倩.葡萄根系抗寒性研究[D].洛阳:河南科技大学硕士论文,2013. ZHANG Qian. (2013).Studyoncoldresistanceofgraperoot[D]. Master Dissertation. Henan University of Science and Technology, Louyang. (in Chinese) [3] 周香云, 李爽, 李连国, 等. 低温胁迫对 8 种葡萄砧木根系细胞膜透性影响的研究[J]. 内蒙古农业大学学报(自然科学版), 2012, 33(4):51-53. ZHOU Xiang-yun, LI Shuang, LI Lian-guo, et al. (2012). Study on the low temperatures stress on memebrane permeability of roots of eight grapevine rootstocks [J].JournalofInnerMongoliaAgricultureUniversity(NaturalScienceEd.), 33(4): 51-53. (in Chinese) [4] 袁军伟,郭紫娟,刘长江,等. 11个葡萄品种根系抗寒性的综合评价[J]. 中外葡萄与葡萄酒, 2015,(2):21-25. YUAN Jun-wei, GUO Zi-juan, LIU Chang-jiang, et al. (2015). Comprehensive assessment of the cold resistance of roots from eleven grape cultivars [J].Sino-OverseasGrapevine&Wine, (2):21-25. (in Chinese) [5] 焦奎宝. 葡萄根系抗寒性的研究[D].长春:东北农业大学硕士论文,2010. JIAO Kui-bao. (2010).Studyoncoldhardinessofgaperoot[D]. Master Dissertation. Northeast Agricultural University, Changchun. (in Chinese) [6] 曹建东,陈佰鸿,王利军,等. 葡萄抗寒性生理指标筛选及其评价[J]. 西北植物学报,2010,30(11):2 232-2 239. CAO Jian-dong, CHEN Bai-hong, WANG Li-jun, et al. (2010). Cold resistance indexes identification and comprehensive Evaluation of Grape Varieties [J].ActaBotanicaBoreali-OccidentaliaSinica, 30(11):2,232-2,239. (in Chinese) [7] 付晓伟,张倩,刘崇怀,等. 评价葡萄根系抗寒性指标的确定[J]. 果树学报,2014,31(1):52-59. FU Xiao-wei, ZHANG Qian ,LIU Chong-huai, et al. (2014). Index for the evaluation of grape root cold resistance [J].JournalofFruitScience, 31(1): 52-59. (in Chinese) [8] 许宏,王孝娣,邹英宁,等. 葡萄砧木及酿酒品种抗寒性比较[J]. 中外葡萄与葡萄酒, 2005,(6):20-23. XU Hong, WANG Xiao-di, ZOU Ying-ning, et al. (2005). Study on the cold resistance of grape rootstocks and wine grape cultivars [J].Sino-OverseasGrapevine&Wine, (6):20-23. (in Chinese) [9] 邹琦. 植物生理学实验指导[M] .北京:中国农业出版社,2000:131-135. ZOU Qi. (2000).Plantphysiologyexperimentinstruction[M]. Beijing:China Agriculture Press:131-135. (in Chinese) [10] 黎燕琼,刘兴良,郑绍伟,等. 岷江上游干旱河谷四种灌木的抗旱生理动态变化[J]. 生态学报,2007,27(3):870-878. LI Yan-qiong, LIU Xing-liang, ZHENG Shao-wei, et al. (2007). Drought resistant physiological characteristics of 4 shrub species in arid valley of Minjiang River [J].ActaEcologicaSinica, 27(3): 870-878. (in Chinese) [11] Morgan, J. M. (1984). Osmoregulation and water stress in higher plants.AnnualReviewofPlantPhysiology, 35(1):299-319. [12] 王燕凌,廖康,刘君,等. 越冬前低温锻炼期间不同品种葡萄枝条中渗透性物质和保护酶活性的变化[J]. 果树学报,2006,23(3):375-378. WANG Yan-ling, LIAO Kang, LIU Jun, et al. (2006). Changes of contents of osmosis substances and antioxidant enzyme activity in grape vines during cold exercise period before over-wintering [J].JournalofFruitScience, 23(3): 375-378. (in Chinese) [13] 高登涛,白茹,鲁晓燕,等. 引入石河子地区的5个葡萄砧木抗寒性比较[J]. 果树学报,2015,(2):232-237. GAO Deng-tao,BAI Ru,LU Xiao-yan, et al. (2015). Study on the cold resistance of five grapevine rootstocks introduced to Shihezi [J].JournalofFruitScience, (2):232-237. (in Chinese) [14]李鹏程,郭绍杰,李铭,等. 4种材料覆盖越冬对葡萄根系抗寒生理指标的影响[J]. 新疆农业科学,2013,50(12):2 230-2 235. LI Peng-cheng, GUO Shao-jie, LI Ming, et al. (2013). The effect of cold resistance indexes of the winter grape roots when covered with four different insulation materials [J].XinjiangAgriculturalSciences, 50(12): 2,230-2,235. (in Chinese) [15] Purvis, A. C., & Yelenosky, G. (1982). Sugar and proline accumulation in grapefruit flavedo and leaves during cold hardening of young trees.JournalAmericanSocietyforHorticulturalScience. (2):222- 226. [16] 鲁金星,姜寒玉,李唯. 低温胁迫对砧木及酿酒葡萄枝条抗寒性的影响[J]. 果树学报 ,2012,29(6):1 040-1 046. LU Jin-xing, JIANG Han-yu, LI Wei. (2012). Effects of low temperature stress on the cold resistance of rootstock and branch of wine grapes [J].JournalofFruitScience, 29(6):1,040-1,046. (in Chinese) [17] Bornman, C. H., & Jansson, E. (1980). Nicotiana tabacum callus studies. X. ABA increases resistance to cold damage.PhysiologiaPlantarum, 48(4):491-493. Research of Cold Resistance of Seven Grape Rootstock Roots Once Introduced in Xinjiang ZHONG Hai-xia, Ermek Caikasimu, ZHANG Fu-chun, PAN Ming-qi, HAN Shou-an,ZHANG Wen, XIE Hui, CHEN Rui, WU Xin-yu (ResearchInstituteofHorticulturalCrops,XinjiangAcademyofAgriculturalSciences/ScientificObservingandExperimentalStationofPomology(Xinjiang),MinistryofAgriculture,Urumqi830091,China) Abstract:【Objective】 In order to provide a scientific basis for introducing excellent cold resistance rootstock and promoting the rootstock cultivation of Xinjiang, this paper screened strong cold resistance grape varieties of the root stock.【Method】Taking one year root system of Xinjiang seven introduction grape rootstock as the experimental material, and detecting root relative electrical conductivity, soluble sugar, free proline, MDA content and soluble protein determination such as physical and chemical indexes under seven low temperature treatments(-2,-4,-6,-8,-10,-12℃). Using subordinate function method to compare and analyze the physical and chemical indicators of various varieties of hardiness comprehensively compared cold resistance of the 7 varieties root system. 【Result】With the stress temperature gradually decreases, the relative electrical conductivity, soluble sugar, free proline, MDA content and soluble protein content of every varieties roots increased gradually, but the higher speed and increase amplitude were slightly different. The subordinate function synthetically analyzed the hardiness was that SO4<5BB<188-08<101-14<110R<5C Key words:grape rootstock; root; old resistance 中图分类号:S663.1 文献标识码:A 文章编号:1001-4330(2016)03-0429-08 作者简介:钟海霞(1988-),女,助理研究员,硕士,研究方向为葡萄栽培与生理,(E-mail)zhonghaixia1@sina.cn通讯作者:艾尔买克·才卡斯木(1969-),男,副研究员,研究方向为果树栽培与贮藏保鲜,(E-mail) amanjol.ok@163.com 基金项目:自治区公益性科研院所基本科研业务经费资助项目(KY2014032);新疆维吾尔自治区青年科学基金项目(2015211B021) 收稿日期:2015-08-19 doi:10.6048/j.issn.1001-4330.2016.03.006 Fund project:Supported by the Basic Science and Technology Research Support Funds of Non-profit Research Institutions of Xinjiang Uygur Autonomous Region (KY2014032)and Youth Science and Technology Research Projects of Xinjiang Uygur Autonomous Region (2015211B021).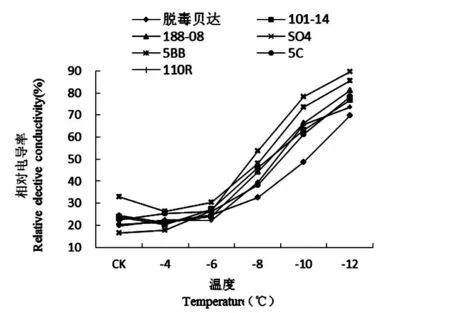
Fig.1The effect of low temperature stress on relative elective conductivity of roots of grape rootstocks
Fig.2 The effect of low temperature stress on free proline content of roots of grape rootstocks
Fig.3 The effect of low temperature stress on MDA content of roots of grape rootstocks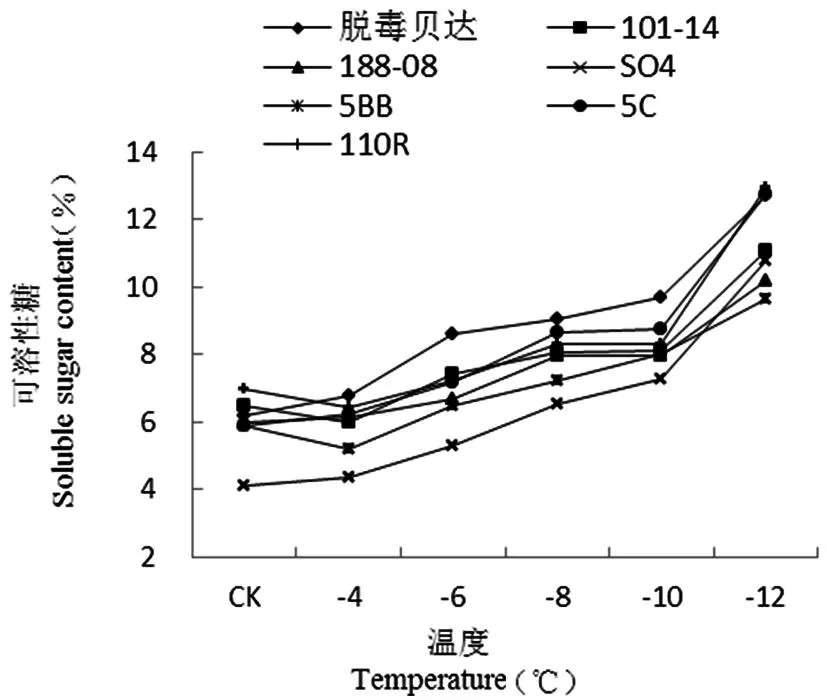
Fig.4 The effect of low temperature stress on soluble sugar content of roots of grape rootstocks
Fig.5 The effect of low temperature stress on soluble protein content of roots of grape rootstocks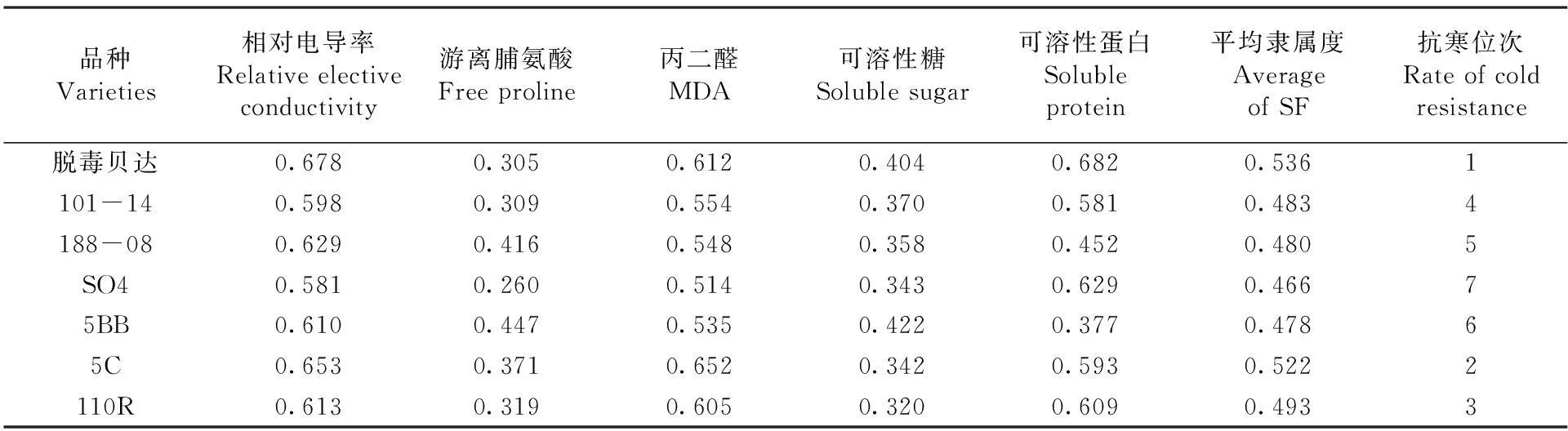 猜你喜欢
现代园艺(2022年7期)2022-11-19 10:18:42热带作物学报(2022年5期)2022-06-01 00:18:44四川劳动保障(2021年3期)2021-06-09 07:08:56现代园艺(2017年13期)2018-01-19 02:28:05新疆农垦科技(2016年2期)2016-08-21 13:50:26西南农业学报(2016年5期)2016-05-17 05:42:36中国农业文摘-农业工程(2016年5期)2016-04-12 05:38:09中国果菜(2016年9期)2016-03-01 01:28:40中国马铃薯(2015年5期)2016-01-09 06:11:11园艺与种苗(2015年10期)2015-02-27 11:27:51
猜你喜欢
现代园艺(2022年7期)2022-11-19 10:18:42热带作物学报(2022年5期)2022-06-01 00:18:44四川劳动保障(2021年3期)2021-06-09 07:08:56现代园艺(2017年13期)2018-01-19 02:28:05新疆农垦科技(2016年2期)2016-08-21 13:50:26西南农业学报(2016年5期)2016-05-17 05:42:36中国农业文摘-农业工程(2016年5期)2016-04-12 05:38:09中国果菜(2016年9期)2016-03-01 01:28:40中国马铃薯(2015年5期)2016-01-09 06:11:11园艺与种苗(2015年10期)2015-02-27 11:27:51
