
海南热带次生林恢复过程中凋落物及土壤水源涵养功能变化*
2016-05-07 06:07林灯陈壮黄荣俊王旭丁昱董翔宇龙文兴2黄瑾2方精李时兴
西部林业科学 2016年2期
关键词:土壤
林灯,陈壮,黄荣俊,王旭,丁昱,董翔宇,龙文兴2,,黄瑾2,,方精,李时兴
(1.海南大学 环境与植物保护学院,海南 海口570228;2.热带作物种质资源保护与利用教育部重点实验室,海南 海口570228;
3.海南大学 园林园艺学院,海南 海口570228;4.海南省黎母山省级自然保护区管理局,海南 琼中572929)
海南热带次生林恢复过程中凋落物及土壤水源涵养功能变化*
林灯1,2,陈壮3,黄荣俊3,王旭1,丁昱3,董翔宇3,龙文兴2,3,黄瑾2,3,方精4,李时兴4
(1.海南大学 环境与植物保护学院,海南海口570228;2.热带作物种质资源保护与利用教育部重点实验室,海南海口570228;
3.海南大学 园林园艺学院,海南海口570228;4.海南省黎母山省级自然保护区管理局,海南琼中572929)
摘要:以海南岛霸王岭刀耕火种和吊罗山择伐恢复的次生林为研究对象,设置幼龄林、中龄林和老龄林样地,研究凋落物及不同土层的最大持水率和自然含水率随干扰类型及恢复阶段的变化规律。结果显示,(1)刀耕火种凋落物和土壤的最大持水率及自然持水率都小于择伐;(2)两种干扰类型的凋落物最大持水率随着森林恢复而减小,自然含水率在3个恢复阶段间无显著差异。刀耕火种土壤的最大持水率随着森林恢复而增大。择伐0~40cm土层最大持水率先减小后增大,40~60cm土壤最大持水率在3个恢复阶段间无显著差异;(3)两种干扰类型的凋落物最大持水率和自然含水率都显著大于土壤;总体上,浅层土壤的最大持水率大于深层土壤,但不同土层间自然含水率无显著差异。(4)两种干扰类型中,凋落物与不同土层间的最大持水率显著负相关,二者间的自然持水率相关不显著。刀耕火种不同深度土壤间的最大持水率显著正相关;择伐不同深度土壤间的最大持水率及自然持水率都显著正相关。研究结果将为中国热带次生林生态恢复提供参考。
关键词:最大持水率;自然含水率;凋落物;土壤;刀耕火种;择伐;热带次生林恢复
森林水源涵养功能是以植被、凋落物以及土壤来阻滞降水、涵蓄水源,从而起到调节地表径流,保持水土的功能[1~3]。其中,凋落物与土壤层间相接、联系紧密[4],共同作为大气降水的主要调节器和蓄存库,在森林水循环中起着重要的作用。一方面凋落物可以抑制土壤水分蒸发,防止雨滴侵蚀并截留部分降雨以涵养水源保护水土[5~7];另一方面土壤的涵养水源的功能间接影响凋落物的组分和数量[8]。而凋落物和土壤的水源涵养功能通常用最大持水率或自然含水率等参数度量[9~10]。
人为干扰后的次生林,其凋落物和土壤的涵养水源功能的影响因素是多方面的,如干扰类型、恢复林龄和土层深度等都对其有直接或间接的影响。干扰类型的不同,水源涵养功能常常会有所差异[11]。林龄对于凋落物和土壤水源涵养功能的影响并非完全一致,如兴安落叶松(Larixgmelinii)林的凋落物最大持水率随林龄增大而递增,但土壤却随林龄增大而减小[10]。凋落物和浅层土壤由于受自身理化特性、地被物和根系的影响,对外界的干扰反应尤为敏感[4]。浅层土壤涵养水源功能往往大于深层土壤[12]。
热带雨林由于高温高湿的气候特性,其水源涵养功能在不同地区乃至全球的生态水循环扮演着重要角色。然而,热带森林受到人为干扰的威胁,不仅原始林面积不断减小,而且由于遭受大规模的刀耕火种和采伐,部分林地出现了水土流失的现象,凋落物和土壤水源涵养功能受到破坏[11~13]。不同干扰类型的热带次生林凋落物和土壤的水源涵养功能是否可以随着林龄的增长而得到恢复,受到国内外生态、气象和水文等方面研究者的广泛关注[12,14~15]。海南岛热带雨林位于亚洲雨林北缘,是印度-马来群系的重要组成部分。由于当地少数民族长期的刀耕火种和大规模的商业性砍伐(如择伐),导致原始林分布破碎化并形成大面积的热带次生林。本文采用空间代替时间的方法[16],选择海南岛刀耕火种和择伐后“幼龄林、中龄林和老龄林”3个恢复阶段的热带次生林的凋落物和土壤为对象,以最大持水率和自然含水率为指标,分析水源涵养功能在热带次生林恢复过程中的变化规律。研究凋落物和土壤水源涵养功能在不同干扰类型、恢复阶段、土层深度的变化规律及相关关系。
1研究区域与方法
1.1样地概况
研究地点主要设置在海南岛霸王岭自然保护区和吊罗山自然保护区。霸王岭自然保护区地理坐标18°52′~19°12′N,108°53′~109°20′E,海拔100~1 654m,面积约500km2,热带季风气候,年均温22.5℃,年均降雨量1 676~2 553mm。吊罗山自然保护区地理坐标18°40′~18°49′N,109°45′~109°57′E,海拔50~1 499m,面积约380km2,年均温24.4℃,年均降雨量1 870~2 760mm。两地均属热带季风气候,降雨量年分布不均,有湿季和旱季区分,土壤类型为山地红壤和山地黄壤,植被沿海拔从低到高依次为热带低地雨林、热带山地雨林、热带山地常绿林和热带山顶矮林。
依据卫星影像资料和实地踏查,我们在霸王岭自然保护区建立8个面积为0.25hm2(50m×50m)的刀耕火种恢复样地(19°06′47″~19°08′56″N,109°07′06″~109°11′20″E),样地海拔范围为405~594m,坡度12°~21°;在吊罗山自然保护区建立9
个0.25hm2(50m×50m)的择伐恢复样地(18°40′18″~18°46′12″N,109°50′20″~109°56′51″E),样地海拔范围为245~750m,坡度7°~27°。所有样地划分为3个恢复阶段,每个恢复阶段至少包含2个重复:幼龄林(林龄约30年)、中龄林(林龄约60年)和老龄林(林龄大于120年)。每块样地的土地利用历史和林龄根据当地林业局的造林资料确定。
刀耕火种恢复样地的3个林龄阶段地面主要由乔灌木的凋落物覆盖,其草本层仅由零星分布的割鸡芒(Hypolytrumnemorum)、草豆蔻(Alpiniahainanensis)等组成,多个样地盖度不足10%;而乔灌木层在幼龄林至中龄林阶段主要由较为耐旱瘠的落叶树种乌墨(Syzygiumcumini)、枫香树(Liquidambarformosana)等乔木和银柴(Aporosadioica)、黄牛木(Cratoxylumcochinchinense)等灌木组成,其郁闭度为67%~81%;在老龄林阶段主要由常绿树种青梅(Vaticamangachapoi)、托盘青冈(Cyclobalanopsispatelliformis)等乔木和粗毛野桐(Hanceahookeriana)、白茶树(Koilodepashainanense)等灌木组成,其郁闭度达到86%~88%。择伐恢复样地3个林龄阶段由于有竹亚科(Bambusoideae)植物侵入,地面凋落物的竹叶组分较高,其草本层由粽叶芦(Thysanolaenalatifolia),芒萁(Dicranopterispedata)和一些蕨类(Pteridiaceae)植物组成,种类相对丰富,盖度达到8%~25%;而乔灌木层在3个林龄阶段均以常绿树种为优势种,主要由青梅、托盘青冈、蝴蝶树(Heritieraparvifolia)等乔木和粗毛野桐、九节(Psychotriaasiatica)、狗骨柴(Diplosporadubia)等灌木组成,郁闭度83%~87%。
1.2数据采集
数据采集在2015年的6月和7月进行。采样期间避开连续降雨或风暴天气,以减少对实验的影响。
1.2.1凋落物持水特性
按五点梅花桩取样法在每一块50m×50m的监测样地四角和中央位置,以1m2(1.0m×1.0m)的水平面积收集凋落物样品并记录其湿重及80℃烘干后的干重,然后计算凋落物自然含水率(WNC0,质量比/%)。采用浸水法测定凋落物的饱和含水率[17~18]。将烘干称重后的凋落物原样放入30cm×30cm的尼龙网袋中,然后将装有凋落物的尼龙网完全浸入盛有清水的容器中,水面略高于尼龙网袋上沿。浸泡24h后将凋落物连同尼龙网一并取出,静置5min左右至凋落物不滴水时测定其湿重,此时的凋落物的持水量为最大持水率,换算凋落物最大持水率(MWH0,质量比/%)。
1.2.2土壤持水特性
在取凋落物样品正下方挖掘土壤剖面,记录土壤剖面发生层次后从上至下将土层分为4层:第1层,0~10cm;第2层,10~20cm;第3层,20~40cm;第4层,40~60cm。用100cm3环刀分别在每层取2个重复土样。利用环刀法测定各土层最大持水率(MWH1~MWH4,质量比/%)、土壤自然含水率(WNC1~WNC4,质量比/%)[19]。
1.3数据分析
首先,分土层绘制MWH0~MWH4和WNC0~WNC4的蚕豆图,分别比较不同干扰类型和恢复阶段的水源涵养功能差异,结果用Randomization-based 双边检验。
然后,分干扰类型和恢复阶段绘制MWH0~MWH4和WNC0~WNC4的玫瑰图,分别比较凋落物与不同土层相互之间水源涵养功能的差异,结果用Randomization-based单边检验。
最后,两种干扰类型分别采用Spearman相关性分析其凋落物和土壤的MWH0~MWH4和WNC0~WNC4之间的相关性,得到相关性分析图。
本文所有的统计检验通过R 3.2.3编写程序完成。
2结果与分析
2.1不同干扰类型间凋落物与土壤水源涵养功能
差异
刀耕火种的MWH0在幼龄林阶段和老龄林阶段显著小于择伐(图1,表1);并且3个恢复阶段刀耕火种的WNC0都小于择伐。在幼龄林阶段和中龄林阶段,刀耕火种的MWH1、 MWH2、MWH3和MWH4都显著小于择伐(图1,表1);而老龄林阶段,不同深度土壤最大持水率在两干扰类型间无显著差异(图1,表1)。在3个恢复阶段,刀耕火种的WNC1、WNC2、WNC3和WNC4基本上显著小于择伐(图1,表1)。
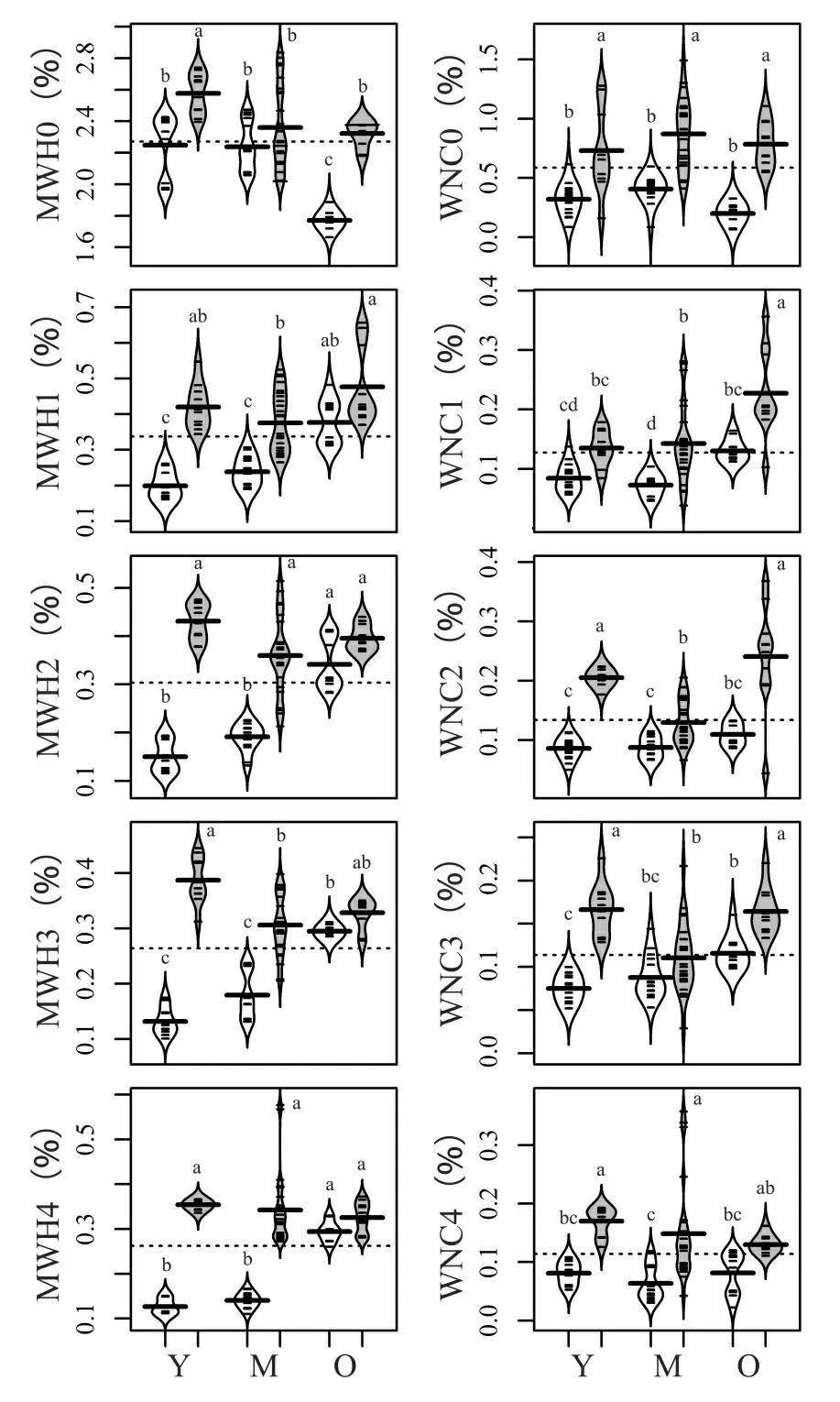
图1 刀耕火种和择伐不同恢复阶段的最大持水率和
2.2不同恢复阶段间凋落物与土壤水源涵养功能差异
刀耕火种和择伐的MWH0随群落恢复整体均呈现下降趋势,但两种干扰类型的WNC0在3个恢复阶段间无显著差异(图1,表1)。刀耕火种的MWH1、MWH2、MWH3和MWH4随群落恢复呈上升趋势,且老龄林显著大于幼龄林;择伐中龄林的MWH1、MWH2和MWH3小于其他两个阶段,而MWH43个阶段间无显著差异。刀耕火种老龄林的WNC1、WNC2和WNC3高于幼龄林和中龄林,而WNC4在3个阶段间无显著差异;择伐的WNC1、WNC2、WNC3和WNC4在3个恢复阶段间变化不一致:WNC1呈现递增趋势;WNC2和WNC3则先减后增,中龄林最小;WNC4在3个阶段间差异不显著。
2.3凋落物和不同深度土壤的水源涵养功能差异
相同干扰类型和相同恢复阶段,凋落物的MWH0和WNC0都显著大于各深度土壤的最大持水率和自然含水率(图2,p< 0.005,表2),表明凋落物比土壤的水源涵养功能高。
刀耕火种幼龄林的MWH1显著大于MWH2、MWH3和MWH4(图2,表3);中龄林的MWH1显著大于MWH4(图2,表3);老龄林的MWH1显著大于MWH3(图2,表3)。择伐幼龄林的MWH1显著大于MWH3(图2,表3);中龄林不同深度土壤间的最大持水率无显著差异(图2,表3);老龄林的MWH1 显著大于MWH3和MWH4(图2,表3)。
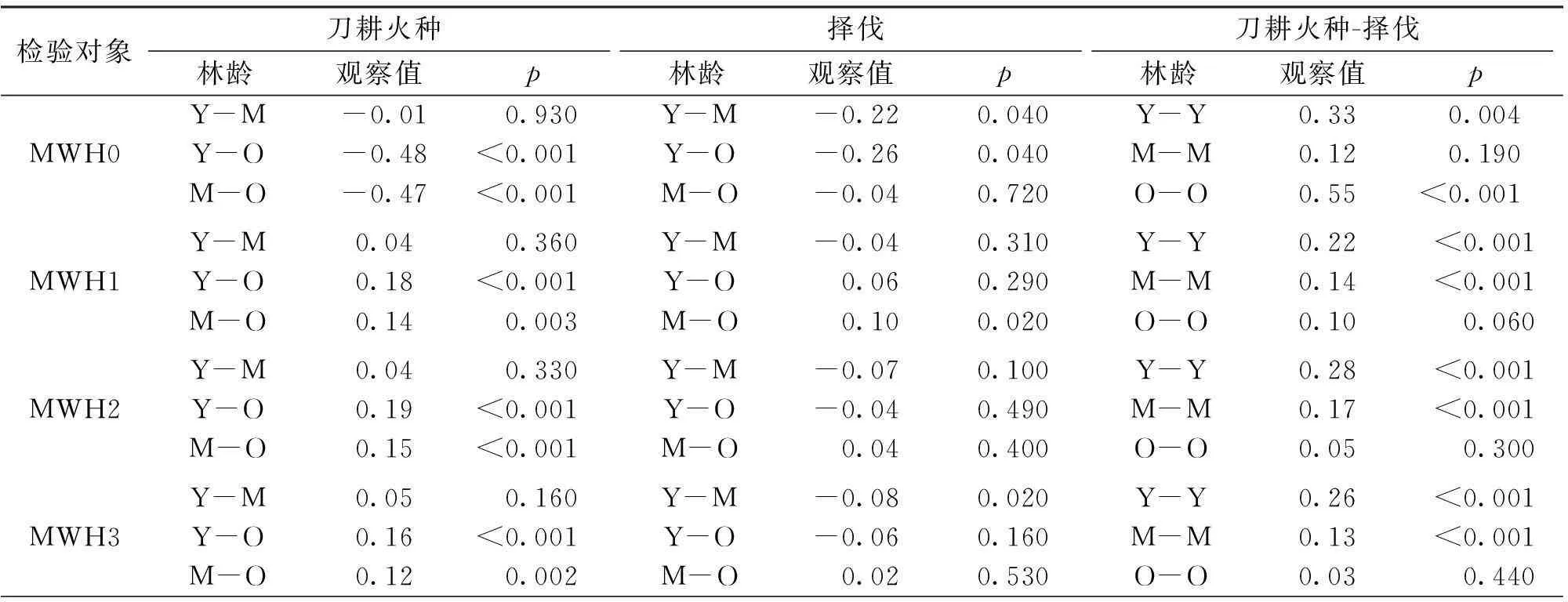
表1 刀耕火种和择伐凋落物和土壤的最大持水率和自然含水率在不同干扰类型和恢复阶段间的Randomization-based 双边检验
续表1
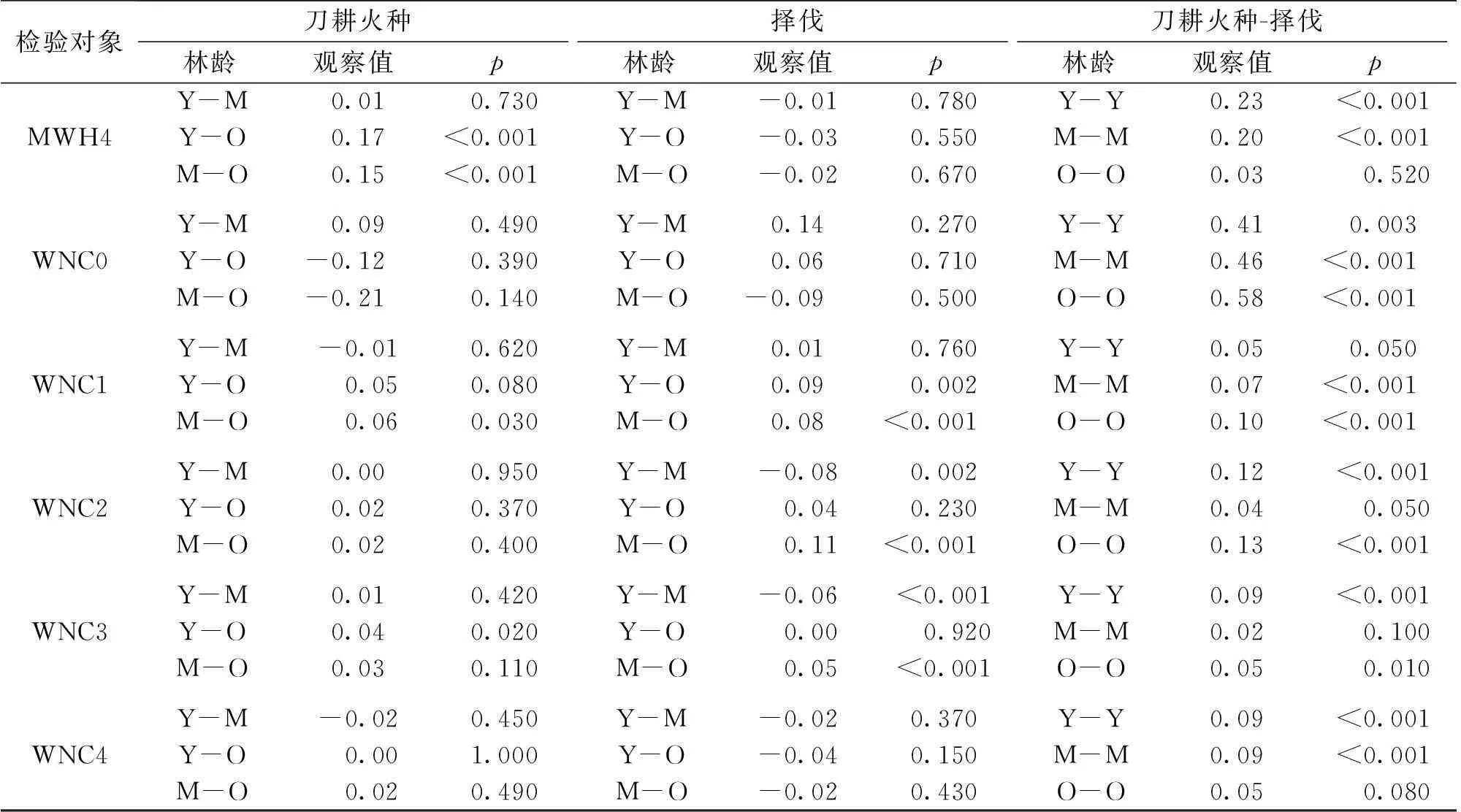
检验对象刀耕火种林龄观察值p择伐林龄观察值p刀耕火种-择伐林龄观察值pMWH4Y-M0.010.730Y-M-0.010.780Y-Y0.23<0.001Y-O0.17<0.001Y-O-0.030.550M-M0.20<0.001M-O0.15<0.001M-O-0.020.670O-O0.030.520WNC0Y-M0.090.490Y-M0.140.270Y-Y0.410.003Y-O-0.120.390Y-O0.060.710M-M0.46<0.001M-O-0.210.140M-O-0.090.500O-O0.58<0.001WNC1Y-M-0.010.620Y-M0.010.760Y-Y0.050.050Y-O0.050.080Y-O0.090.002M-M0.07<0.001M-O0.060.030M-O0.08<0.001O-O0.10<0.001WNC2Y-M0.000.950Y-M-0.080.002Y-Y0.12<0.001Y-O0.020.370Y-O0.040.230M-M0.040.050M-O0.020.400M-O0.11<0.001O-O0.13<0.001WNC3Y-M0.010.420Y-M-0.06<0.001Y-Y0.09<0.001Y-O0.040.020Y-O0.000.920M-M0.020.100M-O0.030.110M-O0.05<0.001O-O0.050.010WNC4Y-M-0.020.450Y-M-0.020.370Y-Y0.09<0.001Y-O0.001.000Y-O-0.040.150M-M0.09<0.001M-O0.020.490M-O-0.020.430O-O0.050.080
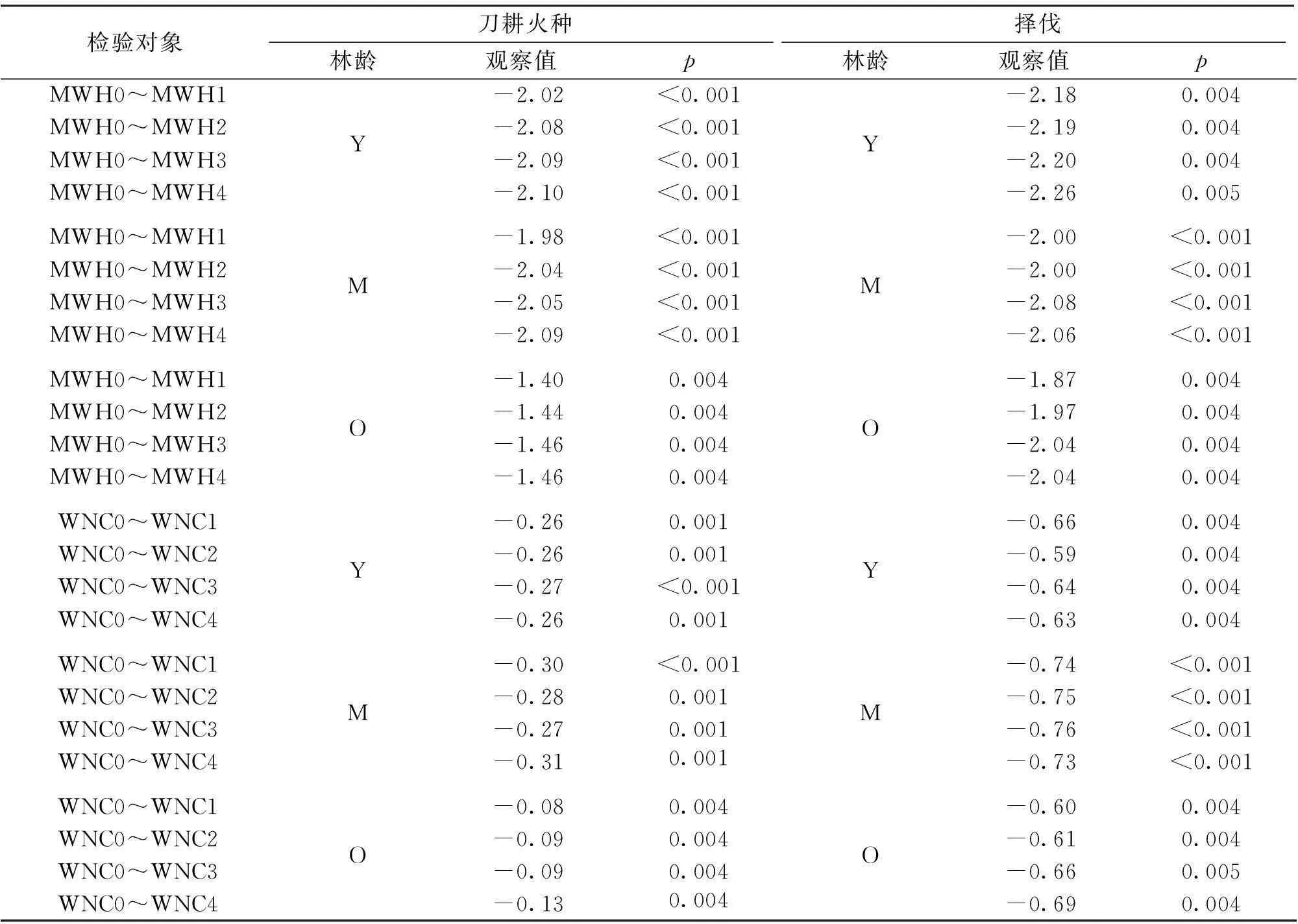
表2 凋落物与土壤之间的最大持水率和自然含水率的Randomization-based 单边检验
注:原假设为凋落物最大持水量<土壤最大持水量;凋落物自然含水量<土壤自然含水量。
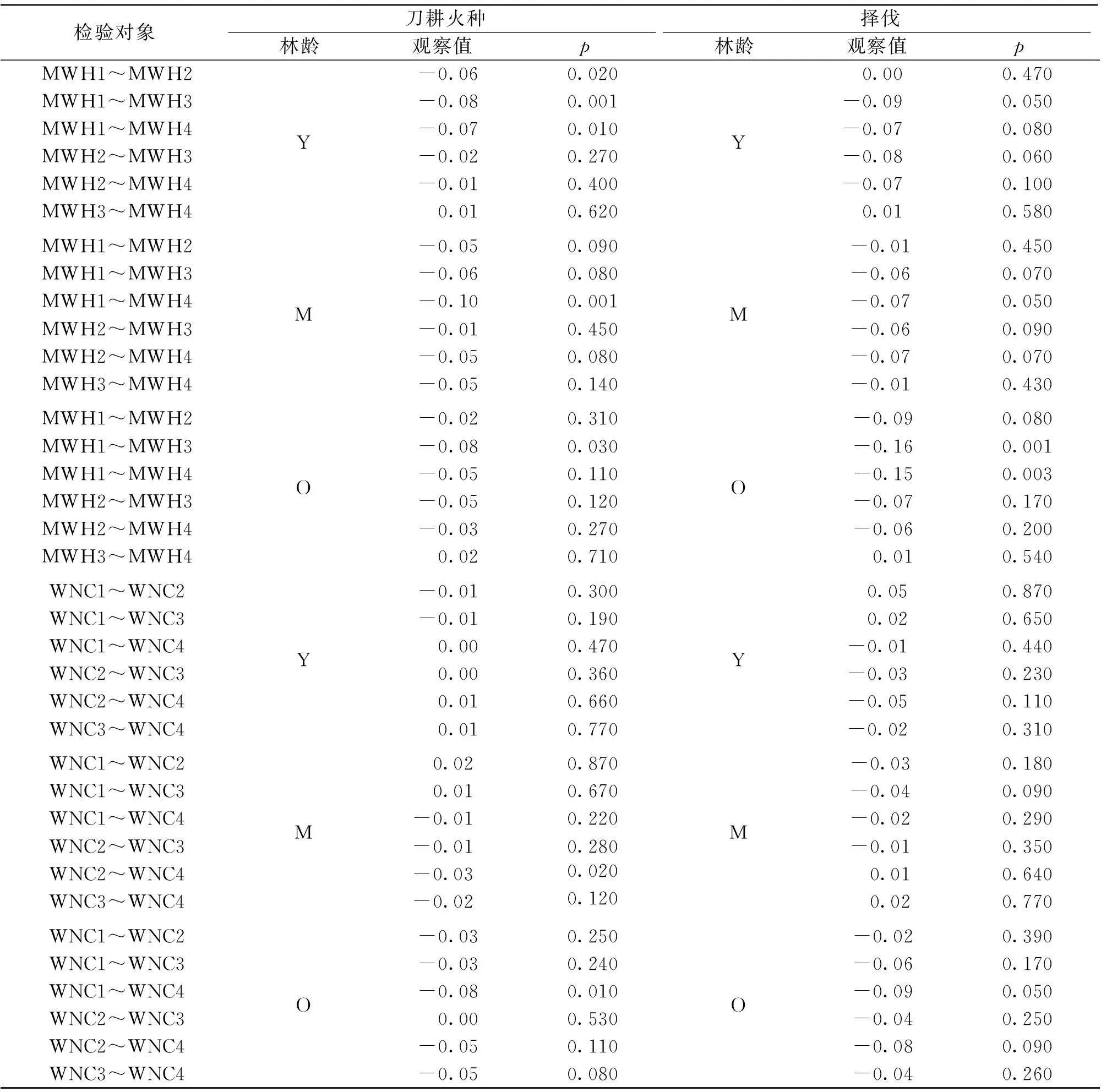
表3 不同深度土壤的最大持水率和自然含水率Randomization-based 单边检验
注:原假设为浅层土壤的最大持水量<深层土壤的最大持水量;浅层土壤的自然含水量<深层土壤的自然含水量。
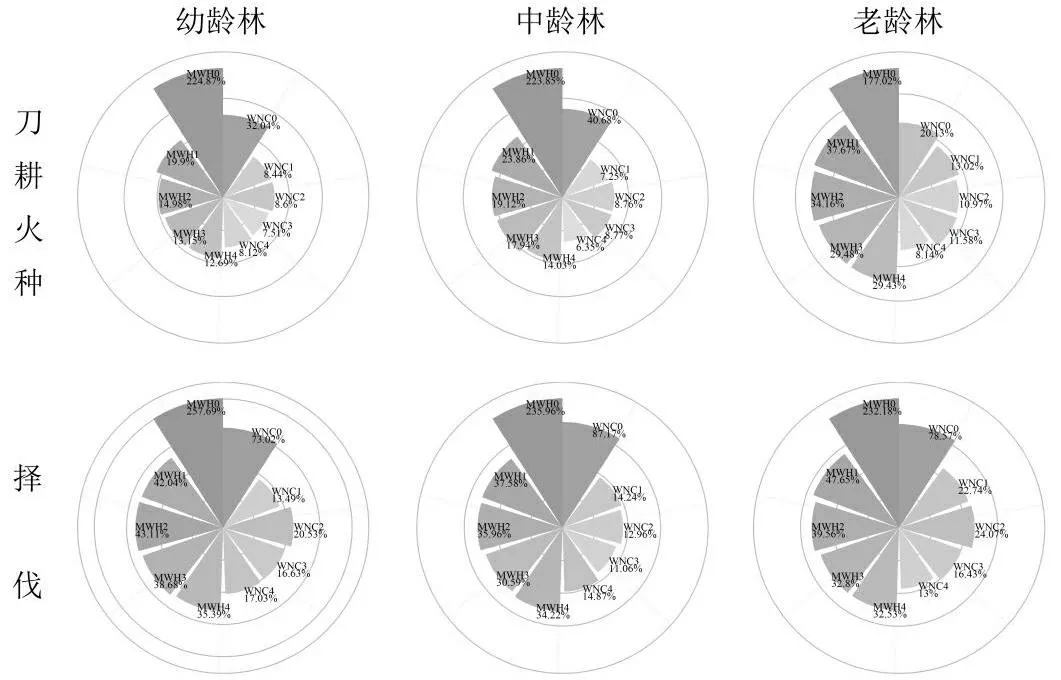
图2 凋落物与不同深度土壤的最大持水率和自然含水率
刀耕火种幼龄林不同深度土壤间自然含水率无显著差异,中龄林的WNC2显著大于WNC4,老龄林的WNC1显著大于WNC4。择伐仅老龄林WNC1显著大于WNC4(图2,表3)。土壤的比较结果表明,浅层的土壤多数情况比深层的土壤有着更高的最大持水率,但是自然含水率上多数情况差异并不显著。
2.4凋落物及土壤水源涵养功能相关关系
刀耕火种的MWH0与MWH1、MWH2、MWH3和MWH4显著负相关;WNC0与WNC1显著负相关,与WNC2、WNC3及WNC4相关不显著;MWH1、MWH2、MWH3及MWH4间显著正相关(图3)。
择伐的MWH0与MWH1显著负相关,而MWH0与MWH2、MWH3及MWH4相关不显著,WNC0与WNC1、WNC2、WNC3及WNC4相关不显著。不同深度土壤间最大持水率及自然持水率各自显著正相关(图3)。
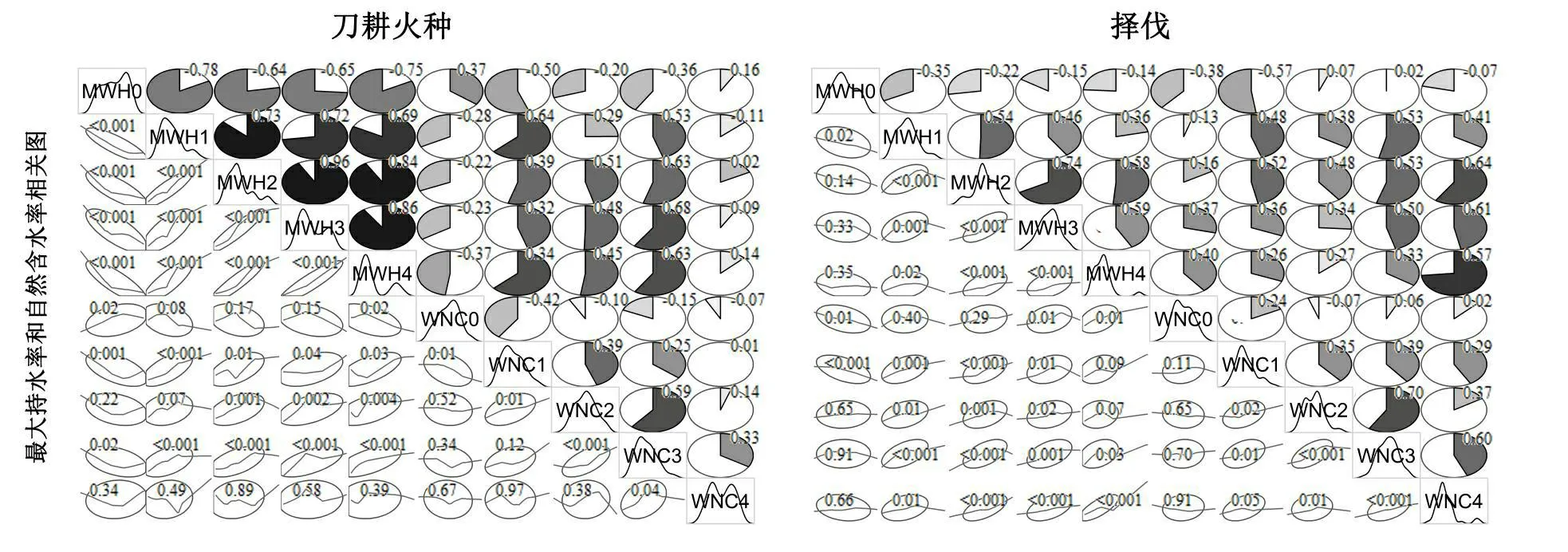
图3不同干扰类型最大持水率和自然含水率间的相关性
注:对角线:变量名及概率分布曲线;上三角:扇形图上标注相关系数,阴影顺时针表示正相关,逆时针表示负相关;椭球限制框表示68%数据集中的位置;loess平滑曲线上方标注相关性分析的p值。
Fig.3Correlation of the maximum water-holding ratio and natural water content in the different disturbance regimes
3讨论
3.1不同干扰类型下水源涵养功能的差异
刀耕火种的凋落物的最大持水率和自然含水率普遍低于择伐(图1,表1)。可能是由于刀耕火种的次生林相对择伐的次生林要干燥,导致前者凋落物降解的速度普遍弱于后者,而半分解的凋落物相对于未分解的凋落物有更高的持水功能[20~21],故在最大持水率上后者要强于前者。刀耕火种的土壤最大持水率在幼龄林和中龄林阶段都显著小于择伐,而老龄林阶段两种干扰类型没有显著差异。可能是由于演替早期,择伐的破坏没有刀耕火种严重,土壤最大持水能力相对恢复较快。但随着林龄增加,土壤最大持水能力的差距逐渐减小,至老龄林阶段基本持平;然而,由于刀耕火种次生林普遍比同恢复阶段的择伐次生林更干燥,导致各恢复阶段刀耕火种的凋落物和土壤自然含水率都小于择伐。
3.2不同恢复阶段水源涵养功能的差异
刀耕火种和择伐干扰类型的凋落物最大持水率随着林龄增长而下降,而其自然含水率在3个恢复阶段并未有显著差异(图1,表1)。凋落物的组分可能是引起其最大持水率变化的主要因素[22]。刀耕火种破坏了原有的植被类型,在幼龄林和中龄林阶段以耐旱耐贫瘠的落叶树种为优势种。这些物种的凋落物相比常绿阔叶树种对于水分的吸收能力更强[22]。但随着演替的进行,落叶树种却逐渐被常绿阔叶树种替代,故而导致凋落物水源涵养功能随林龄增长而逐渐降低;然而择伐破坏后的次生林保留了原有的植被类型,幼龄林、中龄林和老龄林阶段的冠层均以常绿阔叶树种为优势种,但幼龄林阶段因为形成了较大的林窗让生长相对迅速的竹亚科植物占据林下。而竹亚科植物相对常绿阔叶树种易产生更多的凋落物,其凋落物在湿润的环境中降解较快,幼龄林阶段的凋落物半分解比例增加使得其最大持水率相对中龄林和老龄林更高[20~21]。而凋落物的自然含水率,由于凋落物吸持水分的自然过程受区域气候、环境和天气的影响要大于自身组分的影响[5],故出现3个恢复阶段自然含水率变化不显著。
刀耕火种土壤最大持水率随林龄增长而上升(图2,表1),说明刀耕火种早期对于土壤破坏较为严重,土壤保水能力下降,但随着植被的恢复,土壤水源涵养功能也逐渐改善。择伐0~40cm土层最大持水率主要呈现出先减后增的情况,而40~60cm土层3个恢复阶段无显著差异(图1,表1)。表明择伐干扰一定程度上改变了0~40cm土层的物理结构与性质,而40~60cm土层可能由于择伐时破坏的土层深度较浅,并且择伐后保留的植被在一定程度上避免了水土流失,深层土壤的水源涵养功能较大程度得以维持。另外,在不同年代的择伐的破坏程度可能存在差异[23],土壤在最大持水率方面未体现连续性的增长。
不同深度土壤通常对外界干扰、植被和环境变化的敏感程度不同[24]。本研究中,刀耕火种和择伐不同土层自然含水率表现出3个共同规律:①0~10cm土层作为土壤系统与大气系统的交界,对气候和小环境变化最为敏感,故随着森林逐步恢复,郁闭度和空气湿度逐渐增加,表层土壤的自然含水率逐渐增加(图1,表1);②20~40cm土壤相对表层土壤要紧实,该层中的土壤受环境与植被影响较弱,土壤自然含水率主要由土壤物理性质决定。故研究结果表现为土壤自然含水率和土壤最大持水率的变化规律相一致;③40~60cm土层作为较深层次的土壤,外界干扰、植被和环境对其的影响相对弱于较浅土层,故在自然含水率上表现为无显著差异(图1,表1)。
3.3凋落物与不同土层水源涵养功能的差异
凋落物在最大持水率和自然含水率始终显著大于各个土层深度的土壤(图2,表2),与国内外大多数研究结果吻合[24~26]。与此同时,浅层土壤的最大持水率大于深层土壤(图2,表3)。这一现象要归功于凋落物的分解形成的土壤腐殖质,可以显著地由浅入深增加土壤有机质含量,改善土壤结构,提高土壤的涵养水源功能[27~29]。然而,不同土层的自然含水率无显著差异(图2,表3),有可能是因为人为干扰后的土壤相对疏松,透水性较强,浅层和深层的土壤自然含水率相对均衡。
3.4水源涵养功能相关关系
刀耕火种的凋落物与土壤的最大持水率之间呈现负相关(图3)。对于刀耕火种后干旱贫瘠土壤,植被受到胁迫与自然选择,其植物与凋落物组分具备了更强的汲水蓄水能力以抵御恶劣环境,但随着土壤涵养水源功能逐渐改善,植被逐渐重组,其凋落物水源涵养功能随之下降[13]。这种现象充分反映了植被应对环境的自适应过程;自然含水率相关关系不显著(图3)。刀耕火种不同土层的最大持水率主要呈现显著正相关,体现不同深度土壤水源涵养功能都随着林龄增长而逐渐恢复(图3);但自然含水率相关关系同样不显著。刀耕火种破坏以后生境较为干旱,土壤板结硬化,透水性不佳,凋落物和不同深度土壤的自然含水率都易受到环境的影响而产生分化,故相关关系较弱。
择伐的凋落物与土壤的最大持水率和自然含水率的相关性不显著(图3)。表明择伐的凋落物和土壤水源涵养功能没有显著的权衡协同关系。可以归结于,一方面择伐对于植被和土壤的破坏要弱于刀耕火种,在凋落物和土壤的水源涵养功能在演替早期可以较快恢复[11],凋落物和土壤涵养水源功能在幼龄林阶段就已经接近老龄林水平。另一方面由于择伐在不同年代,采伐的强度和目标种会有一定差别[23],导致破坏程度和恢复水平可能有所差异,削弱了植被与土壤的关联性,致使凋落物与土壤涵养水源功能之间无法呈现较为统一的相关关系;但是,单纯比较不同土层的水源涵养功能,由于择伐后土层整体结构未受到严重破坏,水土流失相对较轻,不同层级的协同关系较强,最大持水率和自然含水率均为显著正相关(图3)。
参考文献:
[1]陈光升,胡庭兴,黄立华,等.华西雨屏区人工竹林凋落物及表层土壤的水源涵养功能研究[J].水土保持学报,2008,22(1):159-162.
[2]Wang X C.Problems and relevant strategies on natural forest protection in Changbai Mountain forest area[J].Journal of Forestry Research,2003,14(3):259-262.
[3]Armenteras D,Rodríguez N,Retana J.Are conservation strategies effective in avoiding the deforestation of the Colombian Guyana Shield?[J].Biological Conservation,2009,142(7):1411-1419.
[4]Elliot W J,Dumroese D P,Robichaud P R.The effect of forest management on erosion and soil productivity[A].In:LalR.Soil Quality and Soil Erosion.New York:CRC Press,1999.
[5]Mohanty B P,Famiglietti J S,Skaggs T H.Evolution of soil moisture spatial structure in a mixed vegetation pixel during the Southern Great Plains 1997 (SGP97) Hydrology Experiment[J].Water Resources Research,2000,36:3675-3686.
[6]Prescott C E,Blevins L L,Staley C L.Effects of clear-cutting on decomposition rates of litter and forest floor in forests of British Columbia[J].Canadian Journal of Forest Research,2000,30(11):1751-1757.
[7]Zhang Y,Liu S,Ma J.Water-holding capacity of ground covers and soils in alpine and sub-alpine shrubs in western Sichuan,China[J].Acta Ecologica Sinica,2006,26(9):2775-2781.
[][]
[8]林波,刘庆,吴彦.森林凋落物研究进展[J].生态学杂志,2004,23(1):60-64.
[9]刘世荣,温远光,王兵,等.中国森林生态系统水文生态功能规律[M].北京:中国林业出版社,1996.
[10]王美莲,王飞,姚晓娟,等.不同林龄兴安落叶松枯落物及土壤水文效应研究[J].生态环境学报,2015,24(6):925-931.
[11]许炼烽,朱伍坤.热带次生林利用与土壤物理性质变化[J].生态学报,1996,16(6):652-659.
[12]时雷雷.海南尖峰岭热带山地雨林原始林土壤理化性质空间异质性研究[D].北京:中国林业科学研究院,2012.
[13]黄永涛.海南霸王岭热带低地雨林不同演替阶段土壤理化性质比较[D].重庆:西南大学,2013.
[15]燕东.海南尖峰岭热带雨林凋落物和土壤水文特性研究[D].北京:中国林业科学研究院,2011.
[16]Pickett S T A.Space-for-time substitution as an alternative to long-term studies[A].In:Likens G E.Long-term Studies in Ecology,Approaches and Alternatives.New York:Springer,1989.
[17]Kosugi K,Mori K,Yasuda H.An inverse modeling approach for the characterization of unsaturated water flow in an organic forest floor[J].Journal of Hydrology,2001,246(1):96-108.
[18]Yoshinobu S,Tomo’Omi K,Atsushi K,etal.Experimental analysis of moisture dynamics of litter layers-The effects of rainfall conditions and leaf shapes[J].Hydrological Processes,2004,18(16):3007-3018.
[19]国家林业局.LY/T 1215-1999 森林土壤水分-物理性质的测定[S].北京:中国标准出版社,1999.
[20]刘伦辉,刘文耀,郑征.滇中山地主要植物群落水土保持效益比较[J].水土保持学报,1990(1):36-43.
[21]朱丽晖,李冬,邢宝振.辽东山区天然次生林枯落物层的水文生态功能[J].辽宁林业科技,2001(1):35-37.
[22]王鹏程.三峡库区森林植被水源涵养功能研究[D].北京:中国林业科学研究院,2007.
[23]周新年,巫志龙,郑丽凤,等.森林择伐研究进展[J].山地学报,2007,25(5):629-636.
[24]刘畅.栽针保阔林分土壤水分动态及影响因子分析[D].哈尔滨:东北林业大学,2007.
[25]Kate C,Miguel C,John R.Ammonia volatilization from broiler litter:Effect of soil water content and humidity[J].Soil Science Society of America Journal,2015,79(2):543-550.
[26]逯军峰.不同林龄油松人工林凋落物及其对土壤理化性质的影响研究[D].兰州:甘肃农业大学,2007.
[27]Pritchett W L.Properties and management of forest soils[J].Soil Science,1980,129(6):389.
[28]朱丽琴,黄荣珍,李凤,等.红壤侵蚀地植被恢复后土壤水分特征及其凋落物碳归还模式[J].水土保持通报,2015,35(5):1-6.
[29]Dexter A R,Richard G,Arrouays D,etal.Complexed organic matter controls soil physical properties[J].Geoderma,2008,144(3):620-627.
Variation of Water-conservation Functions of Litter and Soil during Forest Restoration Process in Secondary Tropical Forest
LIN Deng1,2,CHEN Zhuang3,HUANG Rong-jun3,WANG Xu1,DING Yu3,Dong Xiang-yu3,
LONG Wen-xing2,3,HUANG Jin2,3,FANG Jing4,LI Shi-xing4
(1.College of Environment and Plant Protection,Hainan University,Haikou Hainan 570228,P.R.China;2.Key Laboratory of Protection and Development Utilization of Tropical Crop Germplasm Resource,Ministry of Education,Hainan University,Haikou Hainan 570228,P.R.China;3.College of Horticulture and Landscape Agriculture,Hainan University,Haikou Hainan 570228,P.R.China;4.Administration of Limushan Natural Reserve,Qiongzhong,Hainan 572929,P.R.China)
Abstract:To assess the variation in the maximum water-holding ratio and nature-water-content rate of litter and soil in different depths during forest restoration process after shifting cultivation and selective logging,young-aged,middle-aged and old-growth forest plots were established in the secondary forest of Bangwangling and Diaoluoshan of Hainan island.The results showed that,(1)the maximum water-holding ratio and nature-water-content rate of litter and soil after shifting cultivation were lower than those of selecting logging;(2)the maximum water-holding ratio of litter decreased after disturbance,but nature-water-content rate did not differ among the three restoration stages.The maximum water-holding ratio of soil increased.However,the maximum water-holding ratio of 0~40 cm soil decreased and then increased after selecting logging.But it did not differ at the depth of 40~60 cm among the threerestoration stages;(3)the maximum water-holding ratio and nature-water-content rate of litter were greater than those of soil.In general,the maximum water-holding ratio were higher than those of deeper soil,but the nature-water-content rate of soil in different depths did not differ;(4)for the two disturbance regimes,the maximum water-holding ratio of litter was negatively correlated with those of soil in different depths.The nature-water-content rate,however,did not have significant correlation.The maximum water-holding ratio of soil in different depths of shifting cultivation were significantly correlated;the maximum water-holding ratio and nature-water-content rate of soil in different depths were positively correlated with each others.
Key words:maximum water-holding ratio;nature-water-content rate;litter;soil;shifting cultivation; selective logging;restoration of secondary tropical forest
中图分类号:S 714
文献标识码:A
文章编号:1672-8246(2016)02-0007-09
通讯作者简介:龙文兴(1974-),男,副教授,主要从事森林生态学和群落生态学研究。E-mail:oklong@hainu.edu.cn
第一作者简介:林灯(1990-),男,硕士生,主要从事生态系统服务价值研究。E-mail:465798427@qq.com
基金项目:国家自然科学基金项目(31260109,31270474),海南自然科学基金创新研究团队项目(2016CXTD003),海南大学青年 (qnjj1210),海南省自然科学基金项目(312064),中科院战略先导专项项目(XDA05050206、XDA05050208和XDA05050302),海南省研究生创新科研课题(Hys2014-13)。
*收稿日期:2016-02-25
猜你喜欢
红领巾·探索(2021年2期)2021-08-26
今日农业(2020年13期)2020-08-24
学生天地(2020年34期)2020-06-09
军事文摘·科学少年(2020年2期)2020-03-19
石油化工建设(2019年6期)2020-01-16
当代陕西(2019年11期)2019-06-24
作文通讯·高中版(2017年12期)2017-02-06
中国卫生(2016年4期)2016-11-12
山东青年(2016年2期)2016-02-28
中国卫生(2014年4期)2014-12-06
