
拉萨河谷山地灌丛草地植物多样性监测方法的比较研究
2016-04-27 03:36罗黎鸣武建双余成群潘影苗彦军武俊喜明升平郭应杰
草业学报 2016年3期
罗黎鸣,武建双,余成群,潘影,苗彦军,武俊喜*,明升平,郭应杰
(1.中国科学院地理科学与资源研究所,生态系统网络观测与模拟重点实验室,北京 100101;2.西藏农牧科学院农业研究所,
西藏 拉萨 850002;3.西藏大学农牧学院,西藏 林芝 860000;4.西藏大学理学院,西藏 拉萨 850001)
拉萨河谷山地灌丛草地植物多样性监测方法的比较研究
罗黎鸣1,2,武建双1,余成群1,潘影1,苗彦军3,武俊喜1*,明升平4,郭应杰4
(1.中国科学院地理科学与资源研究所,生态系统网络观测与模拟重点实验室,北京 100101;2.西藏农牧科学院农业研究所,
西藏 拉萨 850002;3.西藏大学农牧学院,西藏 林芝 860000;4.西藏大学理学院,西藏 拉萨 850001)
摘要:生物多样性是西藏生态安全监测与评估的重要内容之一。在气候变暖和放牧干扰背景下沿海拔梯度探讨西藏拉萨河谷山地灌丛草地植物群落物种多样性的变化特征,对于该地区草地的保护与可持续利用具有重要意义。以拉萨市林周县白朗村沟域的山地灌丛草地为研究对象,采用常规样方法、重复样线法和Modified-Whittaker样地法3种方法进行系统调查,研究草地群落物种组成及其沿海拔梯度的变化特征。结果表明,1) 3种不同取样方法α多样性的物种丰富度均随海拔的升高呈先增加后降低的趋势,其中50 m样线法效果最好。2) 表征β多样性的Sorensen相似性指数和Bray-Curtis指数均随样地间海拔高差的增大而减小。3) 种-面积关系的截距c、斜率z随海拔的升高呈先增加后降低的趋势,与α、β多样性随海拔梯度的变化特征吻合一致。4) 依据种-面积关系研究结果,山地灌丛草地设置100 m2的样地可覆盖80%左右的植物物种。山地灌丛草地的α和β多样性指数沿海拔梯度均呈先增后减的变化特征,未来研究应注重植物多样性分布格局对山体气候梯度以及放牧干扰变化的响应和适应机制。
关键词:西藏;海拔梯度;物种多样性;监测方法;山地灌丛草地
物种多样性通常从不同层次、尺度分α多样性、β多样性、γ多样性进行度量[1]。其中,β多样性联系群落局部α多样性和区域γ多样性,反映区域内的物种组成随时间、空间的变化,对于理解生态系统功能、加强生物多样性保护和优化生态系统管理具有重要意义[1-4]。物种多样性野外调查方法很多,如样方法和样线法,各有优缺点。样方法[5]有覆盖式的正方形镶嵌式取样方法,如巢式取样法;多尺度长方形镶嵌式取样方法,如Whittaker样地法和Modified-Whittaker样地法。巢式样方因其简便实用在实际调查中经常应用,但其并不足以反映整个地段上的群落特征[6]。而Modified-Whittaker样地法特别适合空间异质性较大的生态系统,能更好地分析物种丰富度[5],广泛应用于全球不同类型生态系统的物种调查中[7-8],也适合研究天然草地物种多样性的空间异质性[9]。样线法是美国监测草地物种丰富度的主要方法[10-11]。样线的长度,重复数多少,间隔距离多少,都会影响样线法监测物种丰富度的精度。在物种多样性研究中取样方法和面积大小在很大程度上会影响研究结果[12],种-面积关系是常用来确定特定群落取样最小的取样面积[13]。国内已有不少研究[14-16]探讨了不同区域、不同草地类型的最优取样法和最小取样面积。研究方法的一致性和可比性是保证数据质量的关键环节[15],同一地区采用不同调查方法取得的物种多样性各指标存在差异,故此有必要比较多种调查监测方法以确定最优取样法和最小取样面积。
青藏高原因其独特的高寒气候和复杂多样的自然条件,造就了丰富多样、独具特色的草地生态系统,是我国宝贵的高原生物基因库和可贵的景观资源[17-18]。高原地区的生物多样性是生态安全监测与评估的重要内容之一[18]。孟凡栋等[19]已对西藏高原3种不同类型的高寒草地植物多样性和地上生物量监测进行了比较研究。董世魁等[20]对青藏高原不同状况(健康和退化)的高寒草地植物多样性的最小样地面积进行研究。而针对青藏高原河谷地区山地灌丛草地植物多样性监测方法的比较研究甚少。
本文采用常规样方法、重复样线法和Modified-Whittaker样地法3种方法对拉萨山地灌丛草地生物多样性进行系统调查。研究植物群落α、β多样性随海拔升高的变化特征,验证种-面积关系参数同α、β多样性的关系,探究山地灌丛草地群落植物多样性的最优取样法和取样面积,旨在揭示西藏“一江两河”地区山地灌丛草地群落物种组成及其沿海拔梯度的变化特征,为该地区天然草地的保护与可持续利用提供科学依据和理论基础。
1材料与方法
1.1研究区域概况
“一江两河”地区属高原温带季风半干旱气候区,年平均气温在4.7~8.3℃,年降水量251.7~580.0 mm之间[21]。土地面积6.65×106hm2,占西藏总面积的5.52%;天然草地4.75×106hm2,占该地区土地面积的71.44%。白朗村沟域位于 “一江两河”的拉萨河流域支流澎波藏曲,地处于念青唐古拉山脉中段南翼,行政隶属于西藏自治区拉萨市林周县卡孜乡白朗村。林周县域海拔3900~3950 m,年日照时数>3000 h,年均温7.5~7.6℃,年均降水量440 mm左右且主要集中在6-9月。
试验地位于白朗村沟域的阴坡上,土壤类型由坡脚至山顶分别发育是亚高山草甸草原土、亚高山灌丛草甸土、高山草甸土;对应的植被类型由下至上分别是亚高山草甸草原、亚高山灌丛草甸、高山草甸。其中,灌丛层片以雪层杜鹃(Rhododendronnivale)、微毛樱草杜鹃(Rhododendronprimuliflorum)、金露梅(Potentillafruticosa)为主。草本层建群种主要为高山嵩草(Kobresiapygmaea)。在整个山体调查中,共记录植物种121个,分属32科,70属。其中灌木植物4种,草本植物117种。样地基本信息见表1。
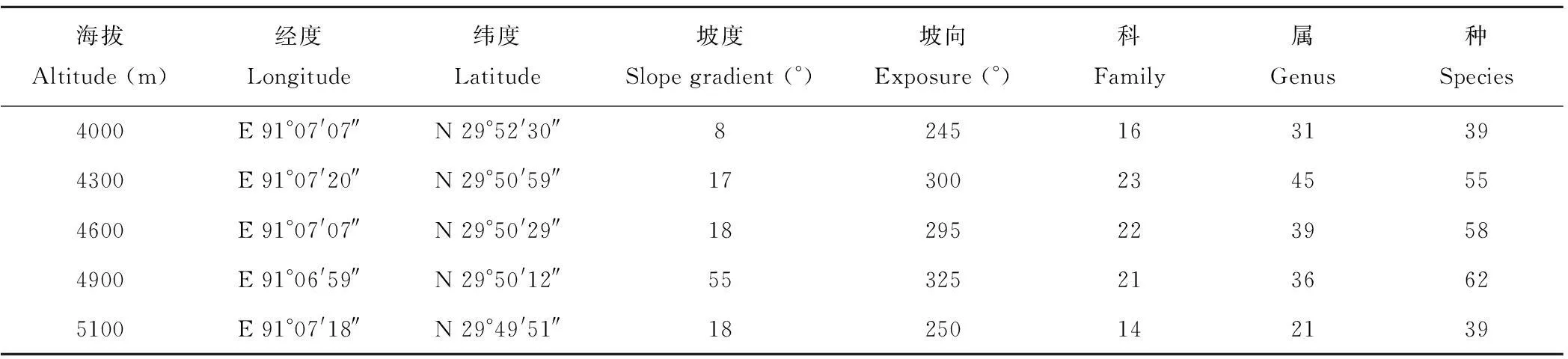
表1 样地基本信息
1.2试验方法

图1 Modified-Whittaker取样法示意图Fig.1 The sketch map of Modified-Whittaker sample
于2014年8-9月,对林周县白朗村沟域的山地灌丛草地沿海拔梯度进行常规样方法、重复样线法和Modified-Whittaker样地法系统调查。从山脚4000 m至山顶5100 m沿坡面按植被类型差异,每隔300 m设置一个样地,共设5个样地,样地大小约50 m×50 m;在每样地随机选取6个面积为0.5 m×0.5 m的样方,做常规草地样方调查,记录每个样方各物种的高度和盖度;在每个样地间隔20 m处设置3条50 m样线,样线方向垂直于样地海拔等高线,每隔5 cm针刺记录所出现的物种和次数,用以测定各物种频度和物种丰富度;在每样地采用Modified-Whittaker样地法[22]进行种-面积关系调查及最小面积的确定,在20 m×50 m的大样方里分别取如下小样方:10个0.5 m×1 m、2个2 m×5 m、1个5 m×20 m、1个20 m×50 m(图1)。
1.3数据处理
1.3.1各物种重要值[23]物种重要值(I)=(Cr+Hr+Fr)/3,其中物种相对盖度(relative coverage,Cr)和相对高度(relation height,Hr)为常规样方法所得,物种相对频度(relative frequency,Fr)为重复样线法统计所得。
1.3.2物种多样性本研究中物种多样性分α多样性和β多样性。α多样性的测定采用物种丰富度指数S。该指数是表示物种多样性最客观、最真实的指标[24]。β多样性的测定[25]采用Sorensen指数和Bray-Curtis指数。Sorensen相似性指数表示不同样地间共有物种占每个样地群落物种总数的比例,能体现群落物种组成出现的重叠效应。Bray-Curtis指数是数量化Sorensen指数,更能体现群落物种组成的变化[26]。Sorensen指数,Cs=2j/(a+b),式中,j为两个群落或样地共有物种数;a和b分别为样地A和样地B的物种数;Bray-Curtis指数,CI=2jI/(aI+bI),式中,jI为样地A(jIa)和样地B(jIb)共有种物种重要值较小者之和,jI=∑min(jIa+jIb),式中,aI、bI分别为样地A和B物种重要值。本研究中α多样性和β多样性Sorensen指数的测算分别采用常规样方法、重复样线法和Modified-Whittaker样地法中1000 m2样地中的物种。Bray-Curtis指数仅采用常规样方法获取的相关数据。
1.3.3种-面积关系本研究中,种-面积关系的公式拟合采用目前公认最简单且最有效的幂函数的对数形式[27-28],即logS=logc+zlogA;公式描述的种-面积关系在双对数坐标轴上是一条直线。一般认为,其c值可以用于表征单位面积的物种多样性;z值则可表征随采样面积增大物种数的积累速率,是物种多样性面积效应的直接体现。因此,种-面积曲线中的c值与z值可以近似理解为α多样性和β多样性[29]。
1.3.4最小面积的确定本研究中,选用非饱和曲线S=b+alnA作为种-面积曲线的拟合模型[30]。要得到群落总物种数一定比例ρ(0<ρ<1)的物种所需的最小面积(或临界抽样面积)[14]为:Amin=exp[(ρSt-b)/a],式中,A为样地面积,S为面积A中出现的物种数,a、b都是模型中待估参数,St是调查群落物种总数。
1.4数据分析
将各海拔梯度上所采的样方数据计算均值,通过SPSS 19.0软件,利用单因子方差分析(One-way ANOVA)的最小显著差数法(LSD)进行分析,作图采用Origin 8.0软件。
2结果与分析
2.1随海拔升高山地灌丛草地植物群落α多样性比较
在3种不同取样方法下,山地灌丛草地植物群落α多样性(物种丰富度)均随海拔的升高呈“单峰曲线”变化特征(图2)。 常规样方法(图2a)在海拔4300 m 处物种数最多(34), 在5100 m处最少(25); 重复样线法(图2b)
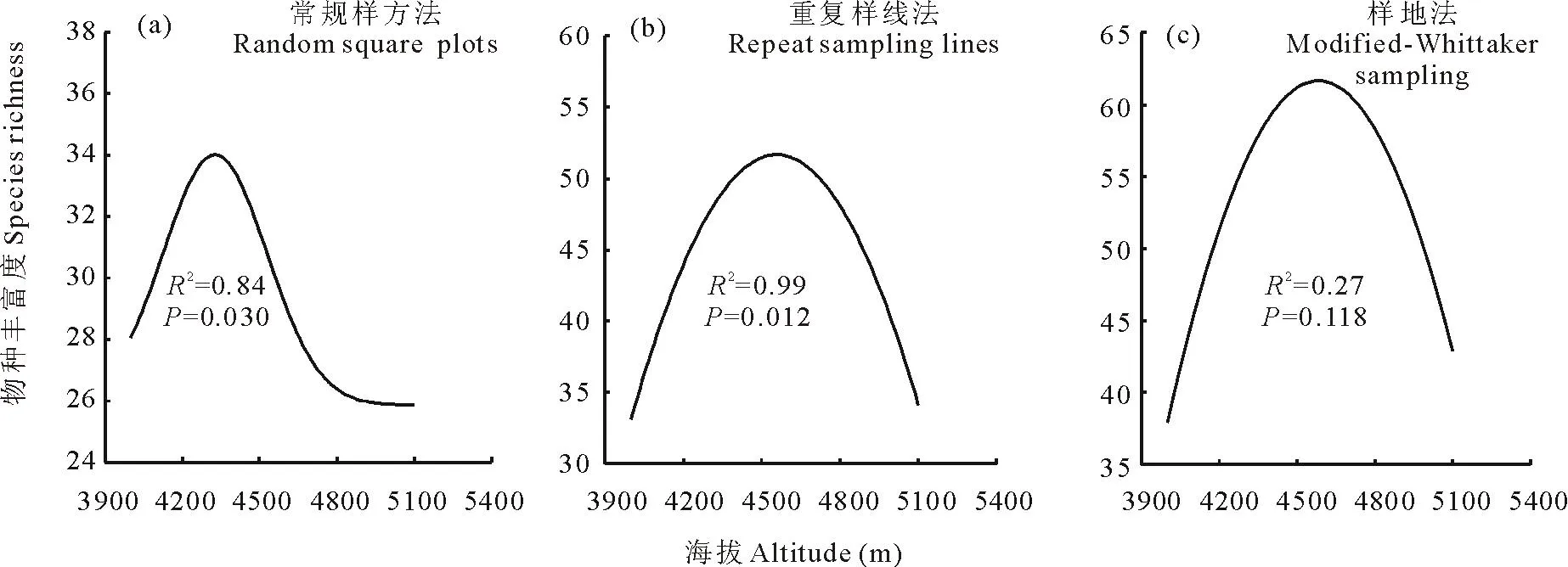
图2 3种取样方法下α多样性随海拔升高的变化Fig.2 Variations of α diversity along an altitude gradient with three sampling methods
在海拔4600 m 处物种数最多(54),在4000 m处最少(34);Modified-Whittaker样地法(图2c)在海拔4900 m 处物种数最多(62),在4000 和5100 m处最少(39)。从物种丰富度最大值来看,Modified-Whittaker样地法>重复样线法>常规样方法。从拟合效果(R2)来看,重复样线法>常规样方法>Modified-Whittaker样地法。综合考虑,本研究推荐50 m样线重复3次可较好地反映这一地区山地灌丛草地物种的α多样性。
2.2随海拔升高山地灌丛草地植物群落β多样性比较
从3种不同取样方法下Sorensen指数测度结果矩阵(表2)来看,β多样性整体上呈与α多样性相同的变化趋势。即Sorensen指数均随样地间海拔高差的增大而减小,群落物种组成差异变大。常规样方法Sorensen指数在海拔4600和4900 m之间最大(0.7143),在海拔4000和5100 m之间最小(0.0755);重复样线法Sorensen指数在海拔4900和5100 m之间最大(0.7436),在海拔4000和5100 m之间最小(0.0909);Modified-Whittaker样地法Sorensen指数在海拔4600和4900 m之间最大(0.7167),在海拔4000和4900 m之间最小(0.1584)。
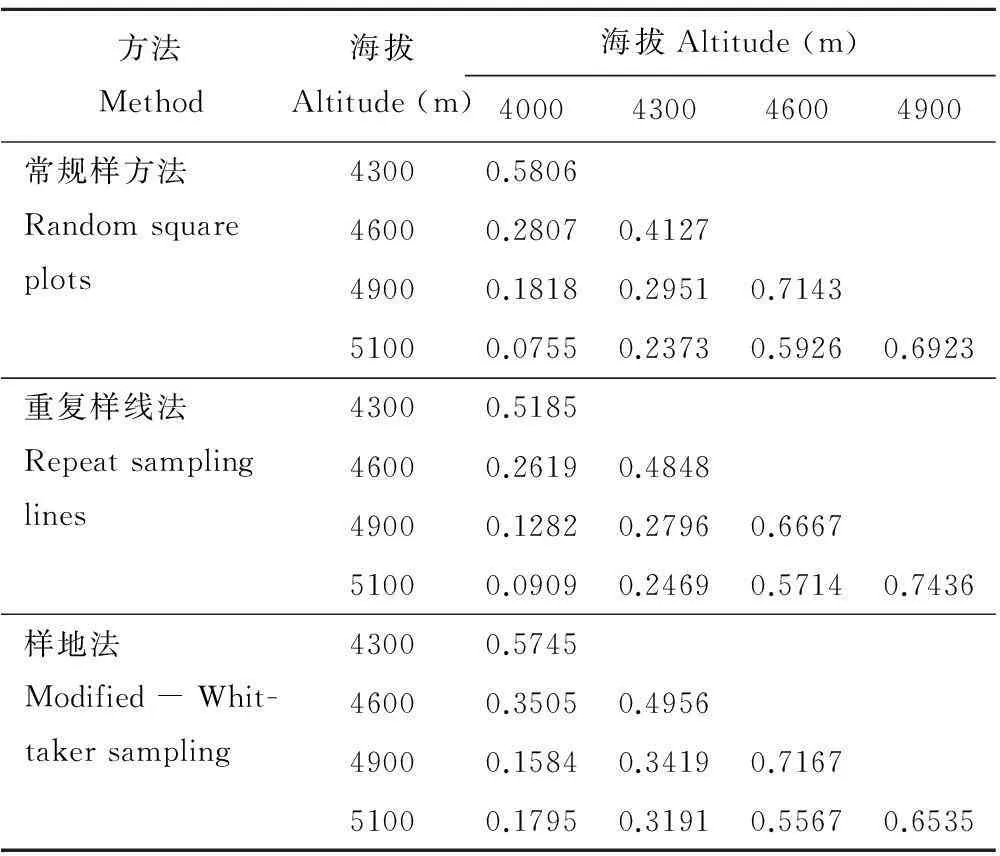
表2 各海拔样地β多样性Sorensen指数测度结果矩阵
从整体上看,β多样性Bray-Curtis指数测度结果矩阵(表3)整体趋势和各Sorensen指数(表2)随海拔的变化趋势基本一致。这说明二者均可用于群落物种组成差异性测度且无明显差异。本研究中Bray-Curtis指数随样地间海拔高差的增大而减小,表明随海拔梯度差的增大,物种组成差异越大。在海拔4600和4900 m之间最大(2.7729),在海拔4000和5100 m之间最小(1.3630)。从Bray-Curtis指数在各海拔梯度之间变化速率来看,中海拔较低海拔、高海拔变化慢,说明中海拔生境差异性较低,物种组成差异较小。
2.3随海拔升高山地灌丛草地植物群落种-面积关系的变化
在同一海拔,随着取样面积的增大物种数呈显著增加。在取样面积相同的情况下,物种数随海拔的升高呈先增加后减少的趋势。在海拔5100 m处,取样面积为1 m2时物种数最少(15.9±2.8);在海拔4900 m处,取样面积为1000 m2时物种数最多(62)(表4)。
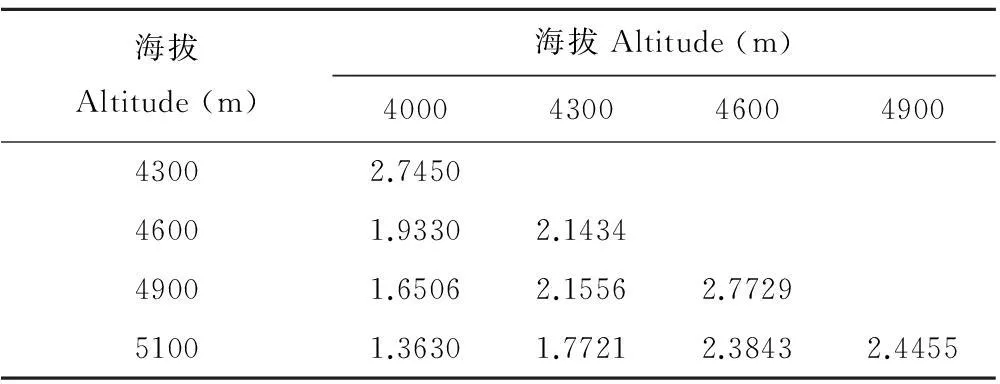
表3 各海拔样地β多样性Bray-Curtis指数测度结果矩阵
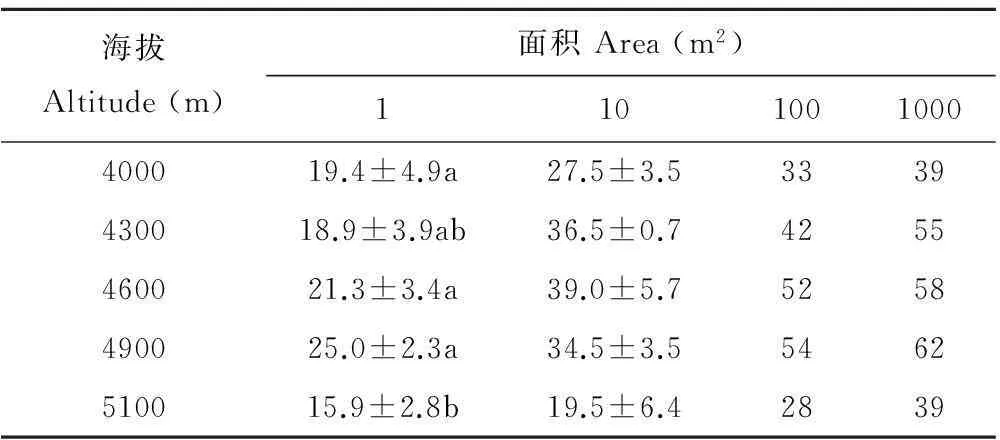
表4 种-面积关系随海拔升高的变化
注:同列不同小写字母表示差异显著(P<0.05)。
Note: Different small letters in the same column indicate significant differences atP<0.05.
种-面积关系截距c随海拔的升高呈先升高后降低的趋势,海拔5100 m处c值最小(15.601),海拔4900 m处c值最大(25.401)。这表明截距c表征的α多样性亦呈“单峰曲线”分布特征,与物种丰富度的分布特征相吻合。随海拔的升高种-面积关系斜率z亦呈先增加后降低的趋势,表明β多样性随海拔的升高呈先增加后降低的趋势。海拔4000 m处z值最小(0.103),海拔4600 m处z值最大(0.157),表明斜率z表征的β多样性在低海拔处最小,中海拔处最大。这可能是因为低海拔处生境差异性较高,物种组成差异较大;中海拔处生境较为均一,物种组成差异较小(图3)。

图3 不同海拔种-面积关系比较Fig.3 Comparison of species-area relationships between different altitude gradients
2.4随海拔升高山地灌丛草地植物群落最小取样面积的确定
拉萨河谷山地灌丛草地种-面积关系拟合结果如表5所示。模型均通过F检验(P<0.0001),但各海拔样地存在明显差异,拟合曲线R2整体随海拔的升高呈先增加后降低的趋势,说明种-面积关系受海拔影响明显。模型参数斜率(a)、截距(b)随海拔的升高呈先增加后降低的趋势。同一海拔最小样地面积随精度ρ的增加而增大。同一精度ρ最小样地面积随海拔的升高呈先增加后降低再升高的趋势。为满足研究精度ρ=60%的最低研究,最小面积至少约为10 m2;为满足研究精度ρ=80%的精确研究,最小面积则至少约为100 m2。
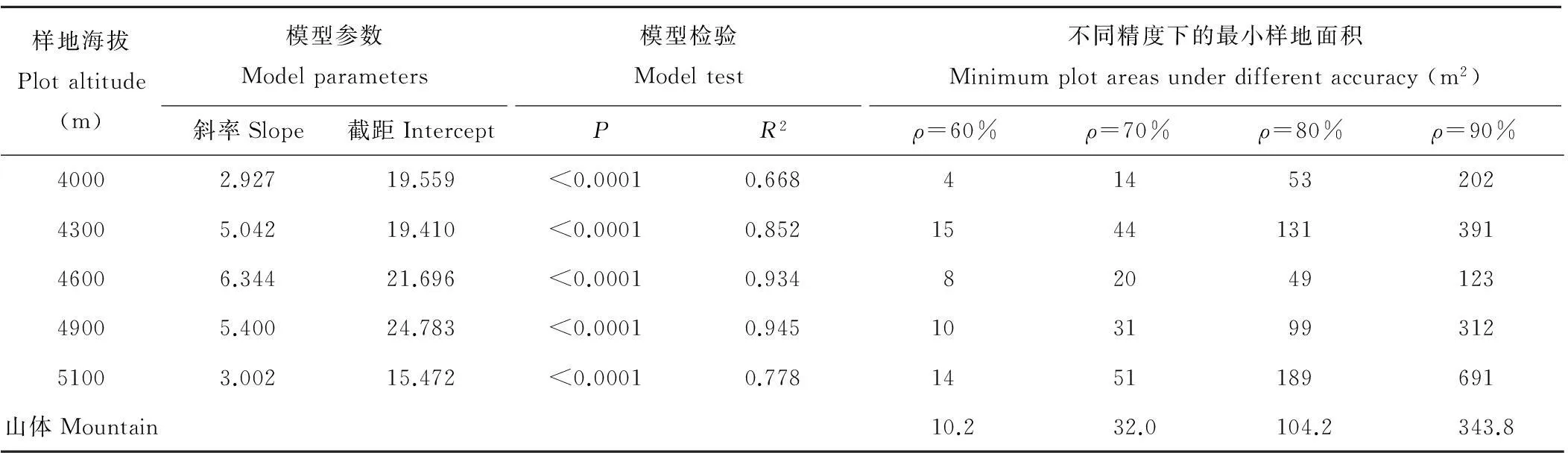
表5 拉萨河谷山地灌丛草地不同海拔样地群落最小面积
3讨论
3.1α多样性随海拔升高的变化特征
α多样性指某一确定采样面积内的物种多样性,反映群落物种组成和结构的多样化程度,是物种多样性研究的基础[31]。玉龙雪山的研究[24]表明木本、草本、全部物种的α多样性(物种丰富度)随海拔的升高均呈“单峰曲线”。青藏高原草甸区的研究[32-33]表明,α多样性随海拔的升高呈“单峰曲线”,即物种丰富度、Shannon-Wiener多样性指数、Pielous均匀度指数随海拔的升高均呈先增加后降低的趋势。拉萨河谷地区山地灌丛草地的已有研究[34]也表明,物种α多样性随海拔的升高呈“单峰曲线”。本研究结果验证了拉萨河谷地区山地灌丛α多样性随海拔的升高呈“单峰曲线”。该地区山地灌丛草地群落α多样性的这种变化特征可能是低海拔受水分制约,高海拔受温度制约,中海拔常有云雾覆盖、水热条件相对均衡等气候差异造成。通过常规样方法、重复样线法和Modified-Whittaker样地法3种取样方法的比较,我们认为常规样方法受草地空间异质性的影响太大,人为选择样地会对结果造成较大偏差,研究中应尽量避免样地选择的主观性;Modified-Whittaker样地法样地面积大,覆盖的物种更多,主要是涵盖了少量稀有种,但稳定性相对差些。重复样线法在评估样地物种丰富度时最全面、最可靠,这可能是因为样线法覆盖了更多的微小生境,且增加样线重复次数,即可较为全面地代表当地物种库。这与孟凡栋等[19]在西藏高寒草地的研究结果不一致,也可能是区域草地类型和数据处理方法差异的原因。
3.2β多样性随海拔升高的变化特征
β多样性指沿着环境梯度不同生境群落之间物种组成的相异性或物种沿着环境梯度的更替速率,反映了生境间的多样性[35]。秦岭太白山群落间的β多样性因坡向而有差异,北坡2800 m以下变化不大,2800 m以上随海拔的升高而降低,南坡整体随海拔的升高而降低[36],说明不同植被类型及地形都会影响生境间物种的多样性。玉龙雪山的木本和灌木的β多样性随海拔的升高呈线性下降趋势,而草本的β多样性随海拔的升高总体呈“单峰曲线”[24],这可能是木本、灌木和草本植物对山地生境的适应策略差异造成的。青藏高原东部甘肃尕海湖草地群落β多样性随海拔的升高而降低[37],这可能是人类活动强烈影响的结果。在本研究中,拉萨河谷山地灌丛草地随海拔的升高呈“单峰曲线”,说明该地区群落生境间在中海拔处相似性最大(差异最小),物种组成在中海拔处差异较小,而在低海拔和高海拔处差异较大。这一特征与物种丰富度的海拔分布特征一致[34]。可能原因是中海拔处气候条件相比高海拔处稳定,且人类活动干扰相比低海拔处弱。
3.3种-面积关系截距c斜率z
国外诸多研究[27-29]表明种-面积关系中截距c、斜率z表征的生态学意义明确,截距c表征了单位面积的物种多样性;斜率z表征随采样面积增大物种积累的速率。乔秀娟等[38]在天山的研究表明草甸和草原的截距c显著高于荒漠,草甸和草原的斜率z显著低于荒漠,说明草甸和草原的α多样性高于荒漠,草甸和草原的β多样性低于荒漠。本文研究中,种-面积关系截距c、斜率z随海拔的升高均呈先增加后降低的趋势,与α、β多样性随海拔升高的变化特征吻合一致。说明种-面积关系中截距c、斜率z也可用来表征拉萨河谷山地灌丛草地生物多样性的变化特征。
3.4群落最小面积
种-面积关系法是确定最小样地面积测定中最常用的方法[20]。关于“面积”的取样方法有很多,国内大多采用常规的巢式取样法[15,39]、改进的巢式样方[40]或以巢式为基础的组合取样法[16,41]。青藏高原针对不同类型[19]和不同健康状况[20]高寒草地物种多样性的研究亦采用巢式样方。巢式样方是经典种-面积曲线的取样法,适用性广,其尺度可从群落到区域乃至世界各大洲[42],其取样严格从总体样本抽取小样本,将大样方套小样方,区域布点均匀,取样方便;但巢式样方只代表了群落的某一局部地段,其确定的最小面积并不足以反映整个地段上的群落特征。巢式样方在一定程度上受到物种集群分布和环境异质性的影响[30]。而Modified-Whittaker样地法特别对空间异质性较大的生态系统具有较高的实用价值,该方法有助于降低空间尺度对物种丰富度的影响,使之更为有效地量度和预测不同生态系统的物种丰富度[5]。为满足山地灌丛草地研究精度ρ=80%的精确研究,本研究建议最小面积至少约为100 m2。这与董世魁等[20]研究的高寒草甸最小面积为108 m2相近,也符合高山嵩草草原草甸所采用的100 m2经验值[43]。
4结论
拉萨河谷山地灌丛草地α多样性随海拔的升高先增加后降低,β多样性随海拔的升高亦呈“单峰曲线”,种-面积关系的参数截距c、斜率z也印证了这种变化特征。说明这是该地区物种多样性呈现的一般规律,是植物对山地气候因子沿海拔梯度变化的适应特征。未来研究应注重植物多样性分布格局对山体气候梯度以及放牧干扰的响应和适应机制。
监测方法是影响野外调查的重要手段。本文中利用50 m样线3次重复能较好地反映拉萨河谷山地灌丛草地的α多样性。Modified-Whittaker样地法的结果很好地反映了拉萨河谷山地灌丛草地的β多样性。为满足研究精度ρ=60%的最低研究,群落最小面积至少约为10 m2;为满足研究精度ρ=80%的精确研究,最小面积至少约为100 m2。我们认为应探究Modified-Whittaker样地法在青藏高原其他草地类型的实用性和可靠性。
致谢:植物鉴定方面得到西藏大学农牧学院郑维列教授和罗建副教授的大力帮助,在此深表感谢!
References:
[1]Jacquemyn H, Honnay O, Pailler T. Range size variation, nestedness and species turnover of orchid species along an altitudinal gradient on Re’union Island: Implications for conservation. Biological Conservation, 2007, 136: 388-397.
[2]Legendre P, Borcard D, Peresneto P R. Analyzing or explaining beta diversity? Comment. Ecology, 2008, 89(11): 3238-3244.
[3]Chen S B, Ou-Yang Z Y, Xu W H,etal. A review of beta diversity studies. Biodiversity Science, 2010, 18(4): 323-335.
[4]Jost L, Devries P, Walla T,etal. Partitioning diversity for conservation analyses. Diversity and Distributions, 2010, 16(1): 65-76.
[5]Liang J Y, Yu C T, Yang X H. Modified Whittaker: a multi scale nested vegetation sampling method. Research of Soil and Water Conservation, 2007, 14: 226-230.
[6]Zhang D Y, Wang G, Zhao S L. Study on the number of plant community succession in the abandoned farmland of the alpine meadow in South Gansu Ⅰ. The characteristic analysis of the succession of pioneer community. Chinese Grassland, 1988, 6: 14-18.
[7]Bashkin M, Stohlgren T J, Otsuki Y. Soil characteristics and plant exotic species invasions in the Grand Staircase-Escalante National Monument, Utah, USA. Applied Soil Ecology, 2002, 22: 67-77.
[8]Adler P B. Neutral models fail to reproduce observed species-time and species-area relationships in Kansas grasslands. Ecology, 2004, 85: 1265-1272.
[9]Wang L, Wang D L, Bai Y G,etal. Spatial distributions of multiple plant species affect herbivore foraging selectivity. Oikos, 2010, 119: 401-408.
[10]Stohlgren T J, Bull K A, Otsuki Y. Comparison of rangeland vegetation sampling techniques in the Central Grasslands. Journal of Range Manage, 1998, (51): 164-172.
[11]Wang S P, Li Y H, Wang Y F,etal. Influence of different stocking rates on plant diversity ofArtemisiafrigidacommunity in Inner Mongolia Steppe. Acta Biotanica Sinica, 2001, 43(1): 89-96.
[12]Drakare S, Lennon J J, Hillebrand H. The imprint of geographical, evolutionary and ecological context on species-area relationships. Ecology Letters, 2006, 9(2): 215-227.
[13]Tang Z Y, Qiao X J, Fang J Y. Species-area relationship in biological communities. Biodiversity Science, 2009, 17(6): 549-559.
[14]Liu C R, Ma K P, Yu S L,etal. Plant community diversity in Donglingshan Mountain, Beijing, China: Ⅶ. The determination of critical sampling areas for several types of plant communities. Acta Ecology Sinica, 1998, 18(1): 15-23.
[15]Wang G P, Wang S P. Comparative study on the sampling methods ofLeymuschinensisgrassland of Inner Mongolia Autonomous Region. Acta Agrestia Sinica, 2003, 11(4): 283-288.
[16]Zhang R, Chen J Q, Hou Y C,etal. Relationships between plant species number and sampling area for sub-alpine meadow plant communities. Chinese Journal of Ecology, 2013, 32(9): 2268-2274.
[17]Zhao H X. Degradation status, causes and improving measures of Tibet grassland. Tibet’s Science and Technology, 2007, 2: 48-51.
[18]Sun H L, Zheng D, Yao T D,etal. Protection and construction of the national ecological security shelter zone on Tibetan Plateau. Acta Geographica Sinica, 2012, 67(1): 3-12.
[19]Meng F D, Wang C S, Zhang Z H,etal. Comparative study of monitoring methods about plant diversity and above-ground biomass of alpine grasslands in the Tibetan Plateau. Acta Botanica Boreali-Occidentalia Sinica, 2013, 33(9): 1923-1929.
[20]Dong S K, Tang L, Wang X X,etal. Minimum plot size for estimating plant biodiversity of the alpine grasslands on the Qinghai-Tibetan Plateau. Biodiversity Science, 2013, 21(6): 651-657.
[21]Zhang G L, Ou-Yang H, Zhou C P,etal. Response of agricultural thermal resources to climate change in the region of the brahmaputra river and its two tributaries in Tibet during past 50 years. Resources Science, 2010, 32(10): 1943-1954.
[22]Stohlgren T J, Falkner M B, Schell L D. A Modified-Whittaker nested vegetation sampling method. Vegetation, 1995, 117: 113-121.
[23]Wu J S, Li X J, Shen Z X,etal. Species diversity distribution pattern of alpine grasslands communities along a precipitation gradient across Northern Tibetan Plateau. Acta Prataculturae Sinica, 2012, 21(3): 17-25.
[24]Feng J M, Wang X P, Xu C D,etal. Altitudinal patterns of plant species diversity and community structure on Yulong Mountains, Yunnan, China. Journal of Mountain Science, 2006, 24(1): 110-116.
[25]Gao X M, Ma K P, Huang J H,etal. Studies on plant community diversity in Donglingshan Mountain, Beijing, China: Ⅺ. The β diversity of mountain meadow. Acta Ecology Sinica, 1998, 18(1): 24-32.
[26]Zhang L M, Gao X, Dong K,etal. Study on community β diversity quantification structures and its measurements. Journal of Yunnan Agricultural University, 2014, 29(4): 578-585.
[27]Tjørve E. Shapes and functions of species-area curves: a review of possible models. Journal of Biogeography, 2003, 30(6): 827-835.
[28]Scheiner S M. Six types of species-area curves. Global Ecology and Biogeography, 2003, 12(6): 441-447.
[29]Connor E F, McCoy E D. Statistics and biology of the species-area relationship. American Naturalist, 1979, 113: 791-833.
[30]Gleason H A. Species and area. Ecology, 1925, 6: 66-74.
[31]Huston M A. Biological Diversity: The Coexistence of Species on Changing[M]. Cambridge: Cambridge University Press, 1994: 142-143.
[32]Wang C T, Wang Q J, Long R J,etal. Changes in plant species diversity and productivity along an elevation gradient in an alpine meadow. Acta Phytoecologica Sinica, 2004, 28(2): 240-245.
[33]Duan M J, Gao Q Z, Guo Y Q,etal. Species diversity distribution pattern of alpine grassland communities along an altitudinal gradient in the Northern Tibet. Pratacultural Science, 2011, 28(10): 1845-1850.
[34]Luo L M, Miao Y J, Wu J S,etal. Variation in biodiversity of montane shrub grassland communities along an altitudinal gradient in a Lhasa River basin valley. Acta Prataculturae Sinica, 2014, 23(6): 320-326.
[35]Ma K M, Ye W H, Sang W G,etal. Study on plant community diversity in Donglingshan Moutain, Beijing, China: Ⅹ. β diversity and fractal analysis on transect on different scales. Acta Ecology Sinica, 1997, 7(6): 626-634.
[36]Tang Z Y, Fang J Y, Zhang L. Patterns of woody plant species diversity along environmental gradients on Mt. Taibai, Qinling Mountains. Biodiversity Science, 2004, 12(1): 115-122.
[37]Xu P B, Deng J M, Zhao C M,etal. Study on grassland community characteristics and species diversity along altitudinal gradients in the Gahai wetland, Gansu province. Acta Prataculturae Sinica, 2012, 21(2): 219-226.
[38]Qiao X J, Tang Z Y, Mmat A N W E,etal. Study on species-area relationships in the southern and northern slopes of the Tianshan Mountains. Arid Zone Research, 2011, 28(1): 54-59.
[39]Jiang J, Zhang C Y, Zhao X H. Plant species-area relationship in a 42-hm2research plot of coniferous and board-leaved mixed forest in Jiaohe, Jilin Province, China. Chinese Journal of Plant Ecology, 2012, 36(1): 30-38.
[40]Bai Y F, Xu Z X, Li D X. Study on α diversity of fourStipacommunities in Inner Mongolia Plateau. Biodiversity Science, 2000, 8(4): 353-360.
[41]Chen H, Li Y Q, Zheng S W,etal. Determination of species-area relationships and minimum sampling area for the shrub communities in arid valley in the upper reach of the Minjiang River. Acta Ecology Sinica, 2007, 27(5): 1818-1825.
[42]Weiher E. The combined effects of scale and productivity on species richness. Journal of Ecology, 1999, 87: 1005-1011.
[43]Wang W Y, Wang Q J, Wang H C. The effect of land management on plant community composition, species diversity, and productivity of alpineKobersiasteppe meadow. Ecological Research, 2006, 21(2): 181-187.
参考文献:
[3]陈圣宾, 欧阳志云, 徐卫华, 等. Beta多样性研究进展. 生物多样性, 2010, 18(4): 323-335.
[5]梁继业, 于春堂, 杨晓晖. 改进的Modified Whittaker:一种多尺度的嵌套式植物多样性取样方法. 水土保持研究, 2007, 14: 226-230.
[6]张大勇, 王刚, 赵松岭. 甘南亚高山草甸弃耕地植物群落演替的数量研究Ⅰ.演替先锋群落的特征分析. 中国草地, 1988, 6: 14-18.
[11]汪诗平, 李永宏, 王艳芬, 等. 不同放牧率对内蒙古冷蒿草原植物多样性的影响. 植物学报, 2001, 43(1): 89-96.
[13]唐志尧, 乔秀娟, 方精云. 生物群落种-面积关系. 生物多样性, 2009, 17(6): 549-559.
[14]刘灿然, 马克平, 于顺利, 等. 北京东灵山地区植物群落多样性研究Ⅶ.几种类型植物群落临界抽样面积的确定. 生态学报, 1998, 18(1): 15-23.
[15]王国平, 汪诗平. 羊草草原植被监测方法的比较研究. 草地学报, 2003, 11(4): 283-288.
[16]张蕊, 陈军强, 侯尧宸, 等. 亚高山草甸植物物种数与取样面积的关系. 生态学杂志, 2013, 32(9): 2268-2274.
[17]赵好信. 西藏草地退化现状、成因及改良对策. 西藏科技, 2007, 2: 48-51.
[18]孙鸿烈, 郑度, 姚檀栋, 等. 青藏高原国家生态安全屏障保护与建设. 地理学报, 2012, 67(1): 3-12.
[19]孟凡栋, 王常顺, 张振华, 等. 西藏高原高寒草地群落植物多样性和地上生物量监测方法的比较研究. 西北植物学报, 2013, 33(9): 1923-1929.
[20]董世魁, 汤琳, 王学霞, 等. 青藏高原高寒草地植物多样性测定的最小样地面积. 生物多样性, 2013, 21(6): 651-657.
[21]张戈丽, 欧阳华, 周才平, 等. 近50年来气候变化对西藏“一江两河”地区农业气候热量资源的影响. 资源科学, 2010, 32(10): 1943-1954.
[23]武建双, 李晓佳, 沈振西, 等. 藏北高寒草地样带物种多样性沿降水梯度的分布格局. 草业学报, 2012, 21(3): 17-25.
[24]冯建孟, 王襄平, 徐成东, 等. 玉龙雪山植物物种多样性和群落结构沿海拔梯度的分布格局. 山地学报, 2006, 24(1): 110-116.
[25]高贤明, 马克平, 黄建辉, 等. 北京东灵山地区植物群落多样性的研究Ⅺ.山地草甸U多样性. 生态学报, 1998, 18(1): 24-32.
[26]张立敏, 高鑫, 董坤, 等. 生物群落β多样性量化水平及其评价方法. 云南农业大学学报, 2014, 29(4): 578-585.
[32]王长庭, 王启基, 龙瑞军, 等. 高寒草甸群落植物多样性和初级生产力沿海拔梯度变化的研究. 植物生态学报, 2004, 28(2): 240-245.
[33]段敏杰, 高清竹, 郭亚奇, 等. 藏北高寒草地植物群落物种多样性沿海拔梯度的分布格局. 草业科学, 2011, 28(10): 1845-1850.
[34]罗黎鸣, 苗彦军, 武建双, 等. 拉萨河谷山地灌丛草地物种多样性随海拔升高的变化特征. 草业学报, 2014, 23(6): 320-326.
[35]马克明, 叶万辉, 桑卫国, 等. 北京东灵山地区植物群落多样性研究Ⅹ.不同尺度下群落样带的β多样性及分形分析. 生态学报, 1997, 7(6): 626-634.
[36]唐志尧, 方精云, 张玲. 秦岭太白山木本植物物种多样性的梯度格局及环境解释. 生物多样性, 2004, 12(1): 115-122.
[37]徐鹏彬, 邓建明, 赵长明, 等. 甘肃尕海湿地不同海拔草地群落组分及物种多样性研究. 草业学报, 2012, 21(2): 219-226.
[38]乔秀娟, 唐志尧, 安尼瓦尔·买买提, 等. 天山南北坡植物种-面积关系. 干旱区研究, 2011, 28(1): 54-59.
[39]姜俊, 张春雨, 赵秀海. 吉林蛟河42 hm2针阔混交林样地植物种-面积关系. 植物生态学报, 2012, 36(1): 30-38.
[40]白永飞, 许志信, 李德新. 内蒙古高原针茅草原群落α多样性研究. 生物多样性, 2000, 8(4): 353-360.
[41]陈泓, 黎燕琼, 郑绍伟, 等. 岷江上游干旱河谷灌丛群落种-面积曲线的拟合及最小面积确定. 生态学报, 2007, 27(5): 1818-1825.
A comparative study of methods for monitoring plant diversity in montane shrub grassland in the Lhasa River Basin
LUO Li-Ming1,2, WU Jian-Shuang1, YU Cheng-Qun1, PAN Ying1, MIAO Yan-Jun3, WU Jun-Xi1*, MING Sheng-Ping4, GUO Ying-Jie4
1.KeyLaboratoryofEcosystemNetworkObservationandModeling,InstituteofGeographicScienceandNaturalResourcesResearch,ChineseAcademyofSciences,Beijing100101,China; 2.AgriculturalResearchInstitute,TibetAcademyofAgriculturalandAnimalSciences,Lhasa850002,China; 3.AgriculturalandAnimalHusbandryCollegeofTibetUniversity,Linzhi860000,China; 4.CollegeofScience,TibetUniversity,Lhasa850001,China
Abstract:Biodiversity is an important consideration in the monitoring and assessment of the ecological stability of rangelands in Tibet. Climate warming and grazing disturbance both threaten the stability of the Tibetan rangelands; hence, research into species succession, transition patterns and species diversity is important for the protection and utilization of grasslands in the region. A field survey was carried out to measure the community species composition and the species change along an altitudinal gradient in the Bailang village valley, Linzhou County in the Lhasa River Basin. Three conventional sampling methods, random square quadrats, repeat sampling lines and the Modified-Whittaker plots, were compared in this study. Key results were: (1) All three sampling methods indicated that α-diversity initially increased and then decreased with increasing altitude, though the line transect method using three 50 m transects detected more species than the other two methods. (2) The Sorensen similarity index and the Bray-Curtis index of β-diversity decreased with increasing altitude. (3) The intercept c and slope z of the relationship between species and sampling area showed unimodal patterns with increasing altitude as also did species richness. (4) In order to cover 80% of local plant species, the minimum sampling area should be up to 100 m2. Future research should focus on the factors determining biodiversity distribution and community assembly in response to climate change and grazing disturbance.
Key words:Tibet; altitudinal gradient; species diversity; monitoring methods; montane shrub grassland
*通信作者
Corresponding author. E-mail: wujx@igsnrr.ac.cn
作者简介:罗黎鸣(1989-),男,湖南邵阳人,硕士。E-mail:luolm1989tibet@126.com
基金项目:国家自然青年基金项目(41401070),国家科技支撑计划项目(2011BAC09B03,2011BAD17B05)和“西部之光”人才培养计划项目(20100309)资助。
收稿日期:2015-04-16;改回日期:2015-09-30
DOI:10.11686/cyxb2015199
http://cyxb.lzu.edu.cn
罗黎鸣,武建双,余成群,潘影,苗彦军,武俊喜,明升平,郭应杰. 拉萨河谷山地灌丛草地植物多样性监测方法的比较研究. 草业学报, 2016, 25(3): 22-31.
LUO Li-Ming, WU Jian-Shuang, YU Cheng-Qun, PAN Ying, MIAO Yan-Jun, WU Jun-Xi, MING Sheng-Ping, GUO Ying-Jie. A comparative study of methods for monitoring plant diversity in montane shrub grassland in the Lhasa River Basin. Acta Prataculturae Sinica, 2016, 25(3): 22-31.
猜你喜欢
学与玩(2022年7期)2022-10-31
小猕猴智力画刊(2017年12期)2017-12-27
海峡姐妹(2017年4期)2017-05-04
科技与创新(2016年21期)2017-02-14
中国高新技术企业(2016年35期)2017-02-10
中国高新技术企业(2016年35期)2017-02-10
绿色科技(2016年21期)2016-12-27
现代农业科技(2016年20期)2016-12-20
科技创新导报(2016年21期)2016-12-17
剑南文学(2016年11期)2016-08-22
