
两种土著食细菌线虫对五种细菌的取食偏好及相关机制①
2016-04-25 07:36闫小梅郑文波李辉信
土壤 2016年1期
闫小梅,虞 丽,李 根,郑文波,武 俊,胡 锋,李辉信
(南京农业大学资源与环境科学学院,南京 210095)
两种土著食细菌线虫对五种细菌的取食偏好及相关机制①
闫小梅,虞 丽,李 根,郑文波,武 俊,胡 锋,李辉信*
(南京农业大学资源与环境科学学院,南京 210095)
摘 要:食细菌线虫通过取食细菌改变土壤微生物的活性和群落结构从而影响土壤生态系统功能,而食细菌线虫取食细菌存在选择偏好。本试验通过设置“自助餐”式平板培养试验,研究了2种土著食细菌线虫(拟丽突属线虫和中杆属线虫)对5种细菌的活菌体、灭活菌体及菌体分泌物的取食偏好,以为进一步揭示土壤生态系统中食细菌线虫与细菌相互作用关系奠定基础。研究结果显示:拟丽突属和中杆属线虫对活菌体的取食偏好基本一致,均对G-菌具有偏好,其偏好顺序为:荧光假单胞菌>大肠杆菌>贪噬菌>解淀粉芽孢杆菌>巨大芽孢杆菌。拟丽突属线虫对5种细菌分泌物的辨识和取食不存在选择性偏好,从而使其对活菌体及灭活菌体的取食偏好顺序基本一致;而中杆属线虫对活菌体及灭活菌体的取食偏好存在显著差异,这可能与中杆属线虫对5种细菌分泌物的辨识和取食具有选择性偏好有关。细菌分泌物对中杆属线虫的取食偏好机制还有待进一步研究。
关键词:食细菌线虫;细菌;取食偏好;分泌物
土壤线虫是土壤动物的主要功能类群之一,是土壤中最为丰富的后生动物[1]。土壤食细菌线虫在土壤根际的比例更高可达90% 左右[2],其通过取食不同细菌而改变微生物数量、活性和群落结构,从而对土壤的碳、氮、磷等(尤其是氮素)转化和循环产生重要影响[3-5]。
生物在取食时往往存在对食物资源的选择性或偏好性,这一现象也适用于低等的原生动物和线虫等[6-7]。例如,研究表明模式线虫Caenorhabditis elegans(秀丽隐杆线虫)会偏爱取食那些易于取食并能很好提供线虫生长的细菌,而远离难于取食的细菌[8]。Niu等[9]研究发现细菌产生的挥发性代谢产物能够吸引线虫,从而影响线虫的取食偏好。食细菌线虫选择性取食土壤细菌不但会改变土壤微生物数量、活性和群落结构,还会直接影响线虫的生存适应性及健康[10-11]。Venette 和 Ferris[12]发现细菌的类型和密度会影响食细菌线虫繁殖率和生长;模式线虫C.elegans的选择性取食还会显著影响其新陈代谢、线粒体DNA复制水平、以及其寿命和产卵数等[13]。因此,土壤食细菌线虫的选择性取食(即偏好性)近年来备受研究者的关注[14-15]。尽管研究人员发现许多的食细菌线虫具有取食偏好性,但这些报道大都是研究线虫对活菌体的取食偏好,而对取食偏好机制,特别是细菌分泌物在取食偏好中的作用等方面的研究尚十分薄弱。
为模拟一个食细菌线虫取食的真实生态环境,本实验从土壤中分离出2株土著食细菌线虫(拟丽突属和中杆属线虫)作为实验材料,同时从该土中筛选出4株优势细菌并以大肠杆菌OP50为对照,研究土著食细菌线虫对不同种类细菌的活菌体、灭活菌体及菌体分泌物(代谢产物)的取食偏好性,以探索菌体分泌物在食细菌线虫取食偏好中的作用。
1 材料与方法
1.1 实验材料
供试菌株:E.coli OP50以及实验室从土壤中分离得到的4株可培养的细菌为:解淀粉芽孢杆菌(B.amyloliquefaciens)、贪噬菌(Variovorax sp.)、巨大芽孢杆菌(B.megaterium)、荧光假单胞菌(P.fluorescens)(表1)。
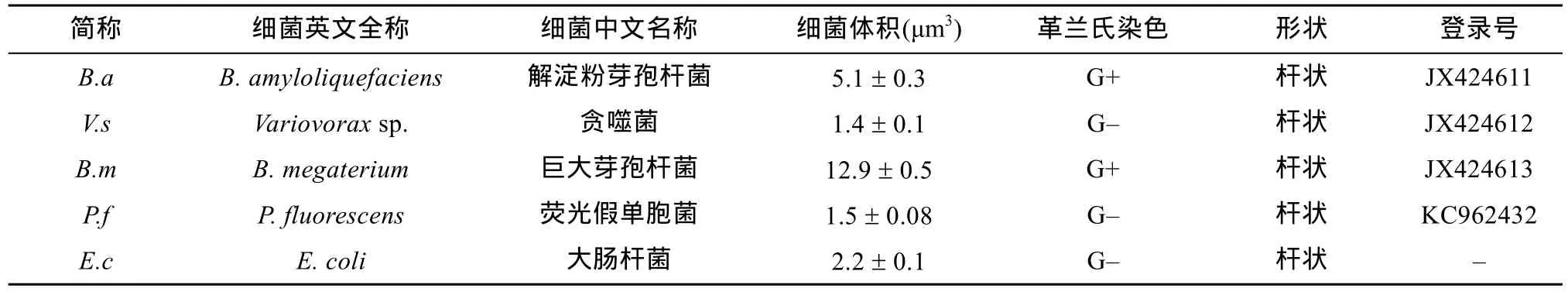
表1 供试细菌基本性质Table 1 Basic characteristics of the bacteria tested
供试线虫:拟丽突属线虫(Acrobeloides sp.):cp值为2,产卵量大,世代时间短,较耐环境压力。中杆属线虫(Mesorhabditis sp.):cp值为1,产卵量巨大,世代时间极短,在食物丰富的条件下种群爆发,极耐环境压力,典型的机会主义者。供试线虫图片见图1。

图1 线虫图片(左图为拟丽突属线虫,右图为中杆属线虫)Fig.1 Images of nematodes(left is Acrobeloides sp.,right is Mesorhabditis sp.)
培养基:线虫培养基(NGM):NaCl 3 g,蛋白胨2.5 g,琼脂 15 g,水 975 ml,121℃灭菌20 min之后,加入胆固醇1 ml,CaCl21 ml,MgSO41 ml,磷酸缓冲液25 ml。细菌液体培养基(LB):NaCl 10 g,胰蛋白胨 10 g,酵母粉 5 g,水 1 000 ml,121℃灭菌20 min。在培养液中加入15 g琼脂粉,即为固体LB培养基。
线虫的培养与制备:将食细菌线虫接种至长有E.coli的线虫培养基(NGM)上并置于20℃的恒温培养箱中黑暗培养。线虫培养20天后用贝尔曼漏斗法分离,收集到的线虫用无菌水反复冲洗后备用。
1.2 实验方法
按照“自助餐”式进行取食偏好实验设计[16],如图2所示,即将直径8.5 cm,高约1.2 cm的培养皿分成5个区域,每个区域中心分别接种上述5种活菌体(灭活菌体或者菌体分泌物),培养基的中心位置接种约1 000条食细菌线虫,共设置8个重复。在接种线虫后的第2、4、8、12、24 h置于体视镜下观察计算线虫在5种活菌体(灭活菌体或菌体分泌物)中的数量。
活菌体的制备和接种量:取5株细菌培养液,10 000 r/min离心,弃上清液,用无菌水冲洗3次后重悬,在5个区域中心分别接种20 μl活菌体(活菌体中含细菌分泌物)。
灭活菌体的制备和接种量:取5株细菌培养液,10 000 r/min离心,弃上清液,用无菌水冲洗3次后重悬,用γ射线灭活后,在5个区域中心分别接种20 μl灭活菌体(灭活菌体不含细菌分泌物)。
菌体分泌物的制备和接种量:取5株细菌培养液,10 000 r/min离心,收集上清悬浮液,经过除菌滤膜处理后,在5个区域中心分别接种20 μl菌体分泌物。
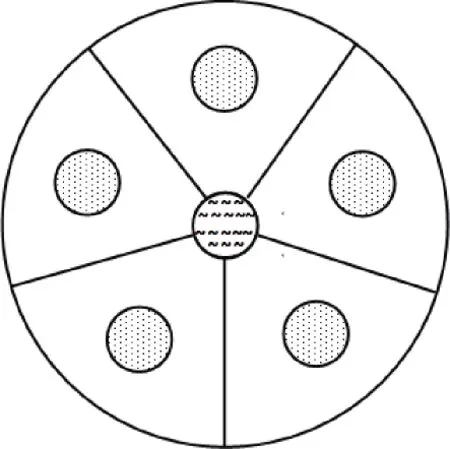
图2 食细菌线虫对不同种类细菌取食偏好实验设计Fig.2 Experimental design of feeding preferences of bacterial-nematodes for different bacteria
1.3 数据处理
用SPSS17.0进行单因素方差分析,用Duncan法进行显著性检验(P<0.05),采用SigmaPlot软件作图。
2 结果与分析
2.1 食细菌线虫对活菌的取食偏好性
从图3可以看出,接种拟丽突属线虫4 h后,5种细菌上的线虫数量较少且没有差异,而8 h后菌体上线虫数量明显增多,P.f 和E.c上线虫的数量显著高于其他几种细菌。在之后的整个培养期内线虫在细菌上的数量随着时间而不断增加,而且P.f上的线虫数量显著高于B.a、V.s、B.m和E.c上的线虫数。在24 h时E.c和V.s上的线虫数没有显著差异,而B.a 和B.m上的线虫数在4~24 h内均无显著差异。可见,拟丽突属线虫偏好选择P.f,其次是E.c和V.s,对B.a 和B.m偏好性较弱。中杆属线虫对5种活菌体的偏好和拟丽突属是一致的,但是中杆属线虫在4 h时对细菌已表现出偏好,而拟丽突属在8 h才表现出偏好。
2.2 食细菌线虫对灭活菌的取食偏好性
从图4可以看出,在接种拟丽突属线虫4 h后,在5种灭活菌上的线虫数量没有差异,直到8 h才出现差异,在24 h时,灭活菌E.c上的线虫数量显著高于其他4种灭活细菌上的线虫数量,灭活的B.a和B.m上的线虫数量在4~24 h内无显著差异。拟丽突属线虫偏好选择灭活的E.c,其次是P.f和V.s,而对灭活的B.a和B.m这两株菌的取食偏好最低。与取食活菌相比,拟丽突属线虫对灭活菌和活菌的取食偏好顺序基本一致。在接种中杆属线虫的整个培养期内,灭活的E.c上的线虫数量显著高于其他几种灭活的菌体上的线虫数量。中杆属线虫偏好选择灭活的E.c,其次是B.a,而对灭活的V.s、B.m和P.f的取食偏好很低。与取食活菌相比,中杆属线虫对灭活的P.f偏好性显著减弱,而对灭活的B.a偏好性显著增加。
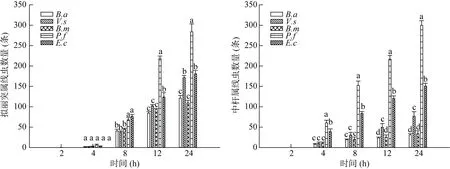
图3 两种线虫对不同种类细菌活菌体的取食偏好Fig.3 Feeding preferences of two nematodes for different live bacteria
图4 两种线虫对不同种类细菌的灭活菌体的取食偏好Fig.4 Feeding preferences of two nematodes for different killed bacteria
2.3 食细菌线虫对细菌分泌物的取食偏好性
接种拟丽突属线虫2 h后,在V.s分泌物上的线虫数量均显著低于其他4种细菌分泌物,而4 h之后的整个培养期内5种菌体分泌物上的线虫数量都没有显著差异。在12 h时,5种菌体分泌物上的线虫数量均有降低的趋势(图5)。中杆属线虫对菌体分泌物的取食偏好与对活菌体的偏好是一致的(图3,图5),偏好选择P.f的分泌物,其次是E.c,而对B.a和B.m这两株菌体分泌物的偏好选择很低。
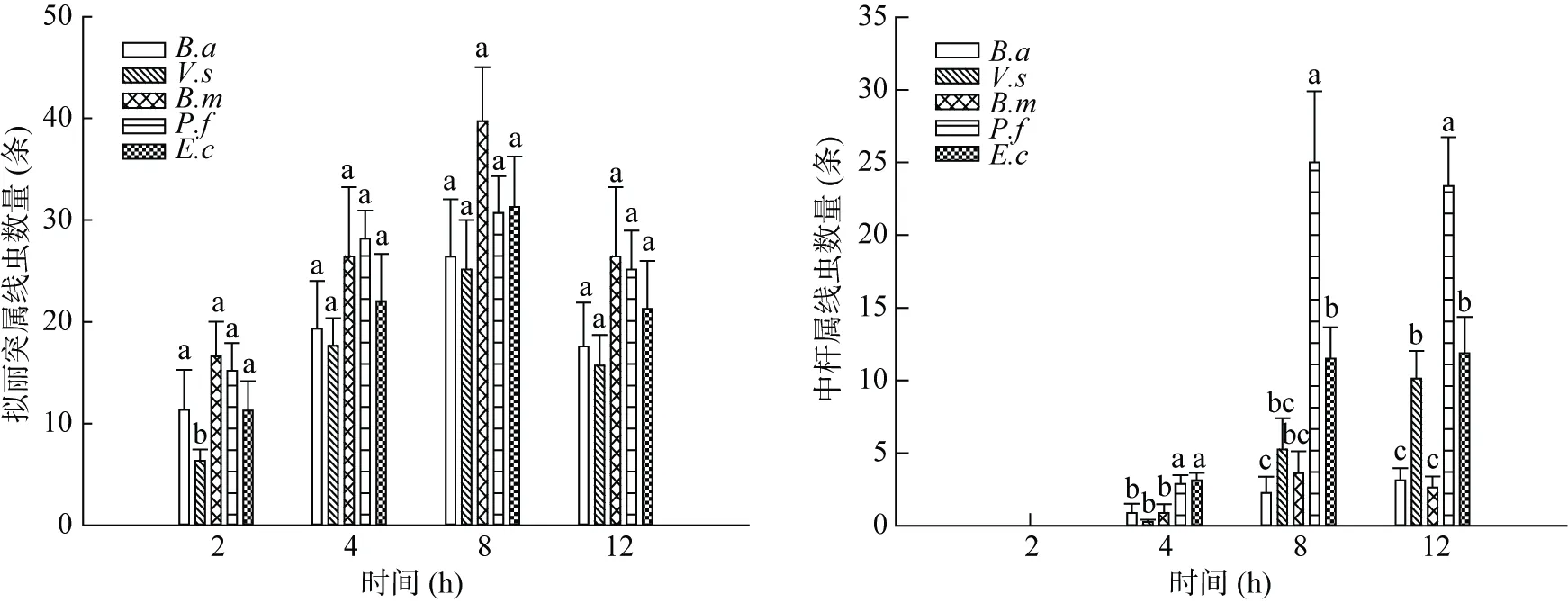
图5 两种线虫对不同种类细菌的分泌物的取食偏好Fig.5 Feeding preferences of two nematodes for different bacterial secretions
3 讨论
食细菌线虫对活菌体的取食存在偏好性,P.f细菌更吸引拟丽突属线虫和中杆属线虫,其次是E.c和V.s,这3株细菌(P.f、E.c、V.s)均为G-细菌,而偏好较差的均为G+细菌。肖海峰等[17]和Salinas等[18]的研究也发现食细菌线虫是对土壤中G- 细菌有更大的偏好。G+细菌的细胞壁比较厚,肽聚糖含量丰富;G-细菌的细胞壁较G+细菌薄,肽聚糖含量低,交联松散,这一差异使得G-细菌被食细菌线虫所取食后更容易被消化。Ronn等[19]研究证明G+细菌会因难以被消化而被原生动物排泄出来。尽管本实验的结果与大多研究者的研究结果一致,即食细菌线虫偏好取食G-细菌,但也有少数研究者发现了相反的结果。如Moens等[20]发现一种海洋中的食细菌线虫(Diplolaimella dievengatensis)偏好取食G+细菌(Halobacillus trueperi),这可能和线虫的种类不同有关。
食细菌线虫对不同灭活菌体的取食同样存在差异,拟丽突属线虫对灭活菌体的偏好与其对活菌的取食偏好顺序基本一致。中杆属线虫对灭活的菌体的偏好与其对活菌体的取食偏好顺序产生明显差异,产生这种现象的原因可能是灭活的P.f不适合中杆属线虫的生长。Moens等[20]在研究海洋食细菌线虫D.meyli的取食时发现,相对于活菌体,该线虫偏好取食灭活的菌体。
细菌分泌物上的线虫数量远远低于活菌体及灭活菌体上的线虫数量,并且在12 h后会有所减少,这与分泌物主要起到吸引线虫而缺少足够的食物供应有关。细菌的分泌物对拟丽突属线虫的取食偏好没有影响,拟丽突属线虫对不同细菌的取食偏好可能主要取决于细菌的其他性状(如大小、形状、G+或G-)。然而,中杆属线虫对细菌分泌物的取食存在明显的偏好;而且中杆属线虫对活菌(含分泌物)和灭活菌(不含分泌物)的取食偏好差异很大。中杆属线虫对灭活的B.a菌有很强的偏好,但是对其活菌和分泌物并没有偏好,这可能是B.a的分泌物对中杆属有抑制作用。事实上,有些细菌的代谢产物对线虫是有毒性的[21-22],至于细菌分泌物的组成和毒性还需进一步的深入研究。
食细菌线虫通过化学信号和趋化机制移至或远离细菌[23]。在本研究中,两种食细菌线虫并没有表现出对细菌的瞬时趋化反应,但8 h内在不同细菌之间的取食选择表现了线虫对细菌具有取食偏好性。取食偏好不仅仅是由趋化性引起的,它是由线虫对食物辨识、寻找、食物的适口性等因素共同影响[24],不同食细菌线虫对细菌取食偏好的机制还需要进行更深入的研究。
参考文献:
[1]江春,黄菁华,李修强,等.长期施用有机肥对红壤旱地土壤线虫群落的影响[J].土壤学报,2011,48(6):1 235-1 241
[2]Bouwman L A,Bloem J,van den Boogert P H J F,et al.Short-term and long-term effects of bacterivorous nematodes and nematophagous fungi on carbon and nitrogen mineralization in microcosm[J].Biology and Fertility of Soils,1994,17(4):249-256
[3]叶成龙,刘婷,张运龙,等.麦地土壤线虫群落结构对有机肥和秸秆还田的响应[J].土壤学报,2013,50(5):997-1 005
[4]Mamilov A S,Byzov B A,Zvyagintsev D G,et al.Predation on fungal and bacterial biomass in a soddy-podzolic soil amended with starch,wheat straw and alfalfa meal[J].Applied Soil Ecology,2001,16:131-139
[5]Fu S L,Ferris H,Brown D,et al.Does the positive feedback effect of nematodes on the biomass and activityof their bacteria prey vary with nematode species and population size[J].Soil Biology and Biochemistry,2005,37:1 979-1 987
[6]Egerton T A,Marshall H G.Feeding preferences and grazing rates of Pfiesteria piscida and a cryptoperidiniopsoid preying on fish blood cells and algal prey[J].Harmfull Algae,2006,5:419-426
[7]Blanc C,Sy M,Djigal D,et al.Nutrition on bacteria by bacterial-feeding nematodes and consequences on the structure of soil bacterial community[J].European Journal of Soil Biology,2006,42:S70-S78
[8]Shtonda B B,Avery L.Dietary choice behavior in Caenorhabditis elegans.Journal of Experimental Biology,2006,209:89-102
[9]Niu Q H,Huang X W,Zhang L,et al.A Trojan horse mechanism of bacterial pathogenesis against nematodes[J].PNAS,2010,107(38):16 631-16 636
[10]Jousset A,Lara E,Wall L G,et al.Secondary metabolites help biocontrol strain Pseudomonas fluorescens CHA0 to escape predation[J].Applied Environmental Microbiology,2006,72(11):7 083-7 090
[11]Zubkov M V,Sleigh M A.Comparison of growth efficiencies of protozoa growing on bacteria deposited on surfaces and in suspension[J].Eukaryot Microbiol,2000,47(1):62-69
[12]Venette R C,Ferris H.Influence of bacterial type and density on population growth of bacterial- feeding nematodes[J].Soil Biology and Biochemistry,1998,30(7):949-960
[13]Reinke S N,Hu X,Sykes B D,et al.Caenorhabditis elegans diet significantly affects metabolic profile,mitochondrial DNA levels,lifespan and brood size[J].Molecular Genetics and Metabolism,2010,100(3):274-282
[14]Zhang Y,Lu H,Bargmann C I.Pathogenic bacteria induce aversive olfactory learning in Caenorhabditis elegans[J].Nature,2005,438:179-184
[15]Jousset A,Rochat L,Pechy-Tarr M,et al.Predators promote defence of rhizosphere bacterial populations by selective feeding on non-toxic cheaters[J].ISME Journal,2009,3(6):666-674
[16]Kevin K N,Jochen R,David A P,et al.Differing preferences of Antarctic soil nematodes for microbial prey[J].European Journal of Soil Biology,2004,40(1):1-8
[17]肖海峰,焦加国,胡锋,等.食细菌线虫Caenorhabditis elegans的取食偏好性[J].生态学报,2010,30(24):7 101-7 105
[18]Salinas K A,Edenbornb S L,Sexstone A J,et al.Bacterial preferences of the bacterivorous soil nematode Cephalobus brevicauda(Cephalobidae):Effect of bacterial type and size[J].Pedobiologia,2007,51(1):55-64
[19]Rinn R,Mc Caig A E,Griffiths B S,et al.Impact of protozoan grazing on bacterial community structure in soil microcosms[J].Applied and Environmental Microbiology,2002,68(12):6 094-6 105
[20]Moen T,Verbeeck L,Maeyer A,et al.Selective attraction of marine bacterivorous nematodes to their bacterial food[J].Marine Ecology Progress Series,1999,176:165-178
[21]Gu Y Q,Mo M H,Zhou J P,et al.Evaluation and identification of potential organic nematicidal volatiles from soil bacteria[J].Soil Biology and Biochemistry,2007,39(10):2 567-2 575
[22]Huang Y,Xu C K,Ma L,et al.Characterisation of volatiles produced from Bacillus megaterium YFM3.25 and their nematicidal activity against Meloidogyne incognita[J].European Journal of Plant Pathology,2010,126(3):417-422
[23]Nicholas W L.The Biology of Free-living Nematodes[M].Oxfod:Clarendon Press,1975
[24]Coolon J D,Jones K L,Todd T C,et al.Caenorhabditis elegans Genomic Response to Soil Bacteria Predicts Environment-Specific Genetic Effects on Life History Traits[J].Plos Genetic,2009,5(6):e1000503
Feeding Preference of Bacterial-feeding Nematodes to Five Different Bacteria and Its Mechanisms
YAN Xiaomei,YU Li,LI Gen,ZHENG Wenbo,WU Jun,HU Feng,LI Huixin*
(College of Resources and Environmental Sciences,Nanjing Agricultural University,Nanjing 210095,China)
Abstract:Bacterial-feeding nematodes can influence the function of soil ecosystems by altering bacterial activity and community structure.There is a feeding preference when bacterial-feeding nematodes confront towards different bacteria.A “cafeteria” diet experiment was set up to study the feeding behavior of Acrobeloides sp.and Mesorhabditis sp.to the living bacteria,killed bacteria and bacterial secretions of five bacteria in order to help us understand better the interactions between bacterial-feeding nematodes and bacterial community structure in soil ecosystems.The results demonstrated that the feeding preferences of Acrobeloides sp.and Mesorhabditis sp.to five living bacteria were similar.The two nematodes both preferred G-bacteria and the feeding preference order was P.fluorescens>E.coli> Variovorax sp.> B.amyloliquefaciens > B.megaterium.Acrobeloides sp.did not show preference to the five bacterial secretions which led to the feeding preference of Acrobeloides sp.to living bacteria and killed bacteria were similar.In contrast,there were big differences in feeding preference of Mesorhabditis sp.to living bacteria and killed bacteria,this may be related to the feeding preference of Mesorhabditis sp.to different bacterial secretions.However,the mechanism of bacterial secretions on feeding preference of Mesorhabditis sp.needs further study.
Key words:Bacterial-feeding nematodes; Bacteria; Feeding preference; Secretion
作者简介:闫小梅(1993—),女,安徽淮北人,硕士研究生,主要从事恢复生态学的研究。E-mail:2012103026@njau.edu.cn
* 通讯作者(huixinli@njau.edu.cn)
基金项目:①国家自然科学基金项目(41271270)资助。
DOI:10.13758/j.cnki.tr.2016.01.013
中图分类号:S154.5
猜你喜欢
小哥白尼(趣味科学)(2021年10期)2022-01-17
中国土壤与肥料(2021年5期)2021-12-02
学生天地(2020年10期)2020-08-25
幼儿画刊(2020年5期)2020-07-24
伴侣(2019年5期)2019-05-27
科技知识动漫(2017年12期)2018-03-07
现代园艺(2017年13期)2018-01-19
人人健康(2017年23期)2017-12-26
儿童故事画报·发现号趣味百科(2015年10期)2016-01-20
华南农业大学学报(2015年5期)2015-12-04
