
3种紫花苜蓿与草地羊茅单、混播越冬期根系生理变化及抗寒性
2016-04-18 08:59申晓慧李如来李建东
草业科学 2016年2期
申晓慧,姜 成,李如来,李建东,张 华,
郑海燕1,郭 伟1,孙 力1,冯 鹏1
(1.黑龙江省农业科学院佳木斯分院,黑龙江 佳木斯 154007;2.沈阳农业大学农学院,辽宁 沈阳110866;
3.佳木斯大学生命科学学院,黑龙江 佳木斯 154007)
3种紫花苜蓿与草地羊茅单、混播越冬期根系生理变化及抗寒性
申晓慧1,2,姜 成3,李如来1,李建东2,张 华1,
郑海燕1,郭 伟1,孙 力1,冯 鹏1
(1.黑龙江省农业科学院佳木斯分院,黑龙江 佳木斯 154007;2.沈阳农业大学农学院,辽宁 沈阳110866;
3.佳木斯大学生命科学学院,黑龙江 佳木斯 154007)
摘要:为了更好地提高紫花苜蓿(Medicago sativa)抗寒性,本研究以紫花苜蓿公农1号、Wega7F、WL319HQ及草地羊茅(Festuca pratensis)为试验材料,分别设置苜蓿单播区与苜蓿-草地羊茅混播区,于2013年10月15日、10月30日、11月15日、2014年3月30日、4月15日、4月30日对苜蓿根系可溶性糖、可溶性蛋白质、游离脯氨酸含量及过氧化物酶(POD)活性4项指标的动态变化进行监测,并对苜蓿越冬率进行调查。结果表明,品种不同,苜蓿单、混播越冬率表现不一。苜蓿与禾草混播效果好于单播,混播越冬率超过90%,公农1号单混播越冬率差异不显著(P>0.05),其它单、混播处理及品种之间差异显著(P<0.05)。各品种单播区与混播区苜蓿根系可溶性糖、可溶性蛋白及游离脯氨酸含量均随气温下降而增高,翌年春季随气温回升而降低,其中可溶性糖含量在11中旬达到最大值,单混播处理差异显著(P<0.05),可溶性蛋白含量在10月末达到最大值,除WL319HQ单、混播处理差异不显著外(P>0.05),其它两个品种单、混播处理差异达到显著水平(P<0.05),WL319HQ单、混播处理在10月末游离脯氨酸含量升到最高,其余各处理均是在11月中旬升到最高值,且同一观测期内,单、混播处理显著差异(P<0.05);POD活性则是随着温度变化在整个越冬期呈现先升后降再上升的变化趋势,各处理均在11月中旬升至最高。通过越冬率调查和运用隶属函数法进行抗寒性综合评判得出,苜蓿与草地羊茅混播处理的越冬率略高于苜蓿单播的,供试材料抗寒性大小顺序为公农1号+草地羊茅>公农1号>Wega7F+草地羊茅>Wega7F>WL319HQ+草地羊茅>WL319HQ。
关键词:紫花苜蓿;草地羊茅;混播;抗寒性;越冬率;生理指标
紫花苜蓿(Medicagosativa)是世界级优良牧草,因其营养价值高、适口性好,具有“牧草之王”的美称。然而在我国东北地区,因冬季严寒干冷,最低气温-30 ℃以下,全年降水也主要集中在盛夏季节,春旱、秋霜冻给苜蓿越冬造成了严重灾害,因此,在东北大部分地区普遍存在着苜蓿越冬率低,容易发生冻害甚至死亡现象,成为制约黑龙江省畜牧业发展的障碍之一[1]。研究紫花苜蓿对温度和寒冷适应的规律和特点是解决这一问题的关键所在[2-3]。南丽丽等[4]、梁慧敏等[5-6]从根系生理生化的变化上深入研究了苜蓿的抗寒性能。史纪安等[7]通过对不同秋眠级别的紫花苜蓿品种根系发育能力的研究表明,根颈分枝能力、根颈粗细及入冬前入土深度,与翌年苜蓿萌发和实现高产密切相关;同时,根颈随气温下降入土越深,其抗寒能力越强。孙启忠等[8]在研究苜蓿不同覆土厚度与根颈入土深度时发现,播种时覆土越深,越能促进根颈膨大,提高植株的耐寒性,进而提高越冬率。对于单播苜蓿抗寒方面的研究已有诸多报道,而苜蓿与禾草混播抗寒规律研究鲜见报道。
根系的抗寒性是决定紫花苜蓿安全越冬的首要因素之一[9-11]。在深秋至入冬前,大多数紫花苜蓿会随着气温的逐渐下降,将其营养物质从地上部逐渐向地下部转移,为安全越冬做准备,所以其根中的营养物质含量也会不断发生变化。冷胁迫下植物可积累更多的可溶性糖、脯氨酸[12]、可溶性蛋白质[13]等营养物质。这些生理指标与植物抗寒性关系密切[14],已有的混播研究主要集中在混播群体的动态变化,混播比例、产草量、混播后草的养分含量变化以及土壤改良等方面[15-16],本研究通过对3种紫花苜蓿单播及分别与草地羊茅混播后,在越冬期对苜蓿根系生理生化指标动态进行分析,检验通过混播播种方式提高苜蓿抗寒性的可行性,了解并揭示紫花苜蓿单、混播在越冬期间根系对低温的适应变化及混播对抗寒性的影响,以期为栽培草地建植和紫花苜蓿生产提供技术支撑。
1材料与方法
1.1试验地概况
试验地土壤为草甸黑土,土壤有机质含量为2.65%、碱解氮135.41 mg·kg-1、速效磷44.50 mg·kg-1、速效钾112.76 mg·kg-1、全氮0.19%、全磷0.14%、全钾1.60%、pH为6.10。
1.2供试材料
试验材料为3个紫花苜蓿品种:公农1号(简称GN1st)、Wega7F由黑龙江省畜牧所提供,WL319HQ为引进品种;1个草地羊茅(FestucapratensisHuds),简称CD,由北京正道生态科技有限公司提供。
1.3试验设计
试验于2013-2014年进行。2013年5月28日播种,间条单、混播,试验小区面积 3 m×4 m, 行距30 cm, 播深2.5~3 cm,苜蓿单播播量22.5 kg·hm-2,混播小区苜蓿(播量4.5 kg·hm-2)+草地羊茅(播量15.75 kg·hm-2),3 次重复,播种时试验小区施底肥,其中尿素(含N 46%)60 kg·hm-2、过磷酸钙(含P2O520%)150 kg·hm-2、氯化钾(含K2O 60%)80 kg·hm-2。小区之间以0.5 m步道相隔。常规田间管理,生长季随时拔除杂草。
1.4测定项目及试验方法
1.4.1取样及保存试验取样于2013年的10月15日、10月30日、11月15日、2014年的3月30日、4月15日和4月30日,挖取紫花苜蓿的根,实验室内将根洗净,用硫酸纸包裹放入-80 ℃冰箱保存,3次重复,取靠近根颈处5 cm长的根系进行各项生理生化指标的测定。各取样日期的温度见图1。

图1 采样期间气温
1.4.2越冬率的调查试验于2013年冬天在各处理小区内选择1 m2大小面积,调查紫花苜蓿植株个数,并在每株苜蓿上系上挂牌,于2014年苜蓿返青后再次调查存活苜蓿植株数,试验设3次重复。计算公式为:
越冬率=返青植株数/植株总数×100%[17]。
1.4.3生理指标测定方法可溶性糖、可溶性蛋白、游离脯氨酸含量的测定见参考文献[18],POD测定采用愈创木酚法[19]。
1.5统计分析
试验数据作图应用Microsoft Excel 2003,利用DPS 7.05数据分析软件进行方差分析,分别对同一处理不同观测期、同一观测期内的不同处理进行单因素方差分析,采用新复极差测验,应用Fuzzy数学中隶属函数法进行抗寒性综合评价,公式如下:
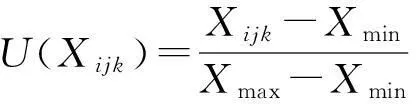
式中,U(Xijk)为第i个处理第j个温度阶段第k个指标的隶属度,且U(Xijk)∈[0,1];Xijk表示第i个处理第j个温度阶段第k个指标测定值;Xmax、Xmin为所有处理中第k项指标的最大值和最小值[20]。
2结果与分析
2.1紫花苜蓿单播及与草地羊茅混播越冬率
各处理单、混播紫花苜蓿越冬情况(图2)结果表明,苜蓿平均越冬率超过了90%,其中GN1st和CD混播处理越冬率最高,达到96.6%,WL319HQ单播越冬率最低,为87.2%。同一品种混播越冬率高于单播,GN1st单、混播越冬率差异不显著(P>0.05),其它两个品种单混播越冬率差异显著(P<0.05),且不同品种之间单播处理越冬率差异也显著 (P<0.05)。
2.2苜蓿根系可溶性糖含量变化
越冬期间随着温度变化,不同处理单播及混播紫花苜蓿根系可溶性糖含量变化呈现先升后降的变化趋势,且除WL319HQ外,其余品种混播根系可溶性糖含量高于单播(表1)。10月30日各品种根系可溶性糖含量开始上升,到11月中旬达到最大值,其中GN1st+CD的可溶性糖积累最多,达到了27.28%, 3月末气温逐渐开始回升,可溶性糖含量仍然较高,这说明在整个冬季紫花苜蓿GN1st和草地羊茅混播后GN1st根内可溶性糖含量都保持在较高水平上。至翌年4月,随着春季气温不断上升,可溶性糖作为春季再生的能源被转移利用,其含量下降,与越冬期含量差异显著(P<0.05)。同一观测期内GN1st和Wega7F单、混播处理差异显著(P<0.05),11月15日至翌年3月30日紫花苜蓿返青后,WL319HQ单混播处理差异显著(P<0.05)。不同品种之间根系可溶性糖含量变化与越冬率也不完全一致,在整个观测期内,Wega7F品种单播根系可溶性糖含量的积累都不是最高,但其越冬能力却较高,说明Wega7F品种能以最低的糖消耗来抵御严寒,提高抗寒性。

图2 不同处理紫花苜蓿越冬率
注:不同小写字母表示不同处理间差异显著(P<0.05)。
Note:Different lower case letters indicate significant difference among different treatments at 0.05 level.
2.3苜蓿根系可溶性蛋白含量变化
各单、混播处理苜蓿根系可溶性蛋白质含量变化总体表现为随田间气温逐渐降低而呈增加趋势,翌年春其含量又随田间气温逐渐上升而下降(表2)。从10月15日到10月30日,各品种及处理平均蛋白含量由15.73 mg·g-1上升到52.62 mg·g-1,上升了234.52%,且均积累到最高含量,除WL319HQ单、混播差异不显著外,其它处理混播可溶性蛋白含量显著高于单播(P<0.05),随着低温强度的增加,可溶性蛋白含量有所下降,但仍然保持较高含量,这对品种抗寒和生存有重要意义。10月15日即越冬初期,各个处理之间可溶性蛋白含量差异不显著(P>0.05),但同一时期不同品种之间可溶性蛋白积累量不一,在最冷月(11月15日)同一品种混播处理显著高于单播(P<0.05),且国内品种GN1st和Wega7F的可溶性蛋白含量高于引进品种WL319HQ,至4月30日,除GN1st单播显著大于混播外(P<0.05),其余两个品种单、混播处理差异不显著,且此时可溶性蛋白含量大幅下降,可能是此时苜蓿返青,植物体内物质和能量代谢加快所致。
2.4苜蓿根系游离脯氨酸含量变化
各处理根系游离脯氨酸含量随气温变化呈先升后降趋势,且混播含量高于单播(表3)。经过10月15日至11月15日的温度大幅下降,各处理根系游离脯氨酸含量显著升高(P<0.05),GN1st+CD混播的游离脯氨酸含量最高为22.84 mg·g-1,除WL319HQ单、混播处理在10月30日含量升到最高值外,其余各处理均是在11月15日升到最高值,之后游离脯氨酸含量逐渐下降,至翌年4月30日,WL319HQ单、混播处理游离脯氨酸含量平均下降了83.26%, GN1st单、混播处理下降80.88%, Wega7F单、混播处理下降最慢,为79.66%。GN1st和Wega7F在同一观测期内混播显著高于单播处理(P<0.05), WL319HQ单、混播处理在10月30至11月15日期间也表现出混播显著高于单播。作为渗透调节物质之一的游离脯氨酸,在植物体内除了具有低温胁迫时起到平衡细胞代谢的功能外,还能维持胞内环境的相对稳定,混播处理游离脯氨酸含量高于单播,说明苜蓿混播处理较单播处理更有利于抵御严寒。

表1 不同处理紫花苜蓿根系可溶性糖含量(%)变化
注:同行不同小写字母表示同一处理不同日期间差异显著(P<0.05);同列不同大写字母表示同一观测期不同处理间差异显著(P<0.05)。表2、表3、表4同。
Note: Different lower case letters within the same row indicate significant difference among different sampling dates for the same treatment at 0.05 level. Different capital letters within the same column indicate significant difference among different treatments for the same sampling date at 0.05 level, The same in Table 2, Table 3 and Table 4.

表2 各单、混播处理紫花苜蓿根系可溶性蛋白含量(mg·g-1)变化

表3 不同处理紫花苜蓿根系游离脯氨酸含量(mg·g-1)变化
2.5苜蓿根系过氧化物酶(POD)含量变化
不同苜蓿品种单播及与草地羊茅混播后,POD活性均呈现先升后降再上升的变化趋势(表4)。10月15日到11月15日随着气温的降低,各品种苜蓿根系POD活性增至最高,除WL319HQ外,其余品种苜蓿根系POD活性表现为混播略高于单播,越冬率高的品种GN1st和Wega7F高于越冬率低的品种WL319HQ。之后随着低温胁迫时间的延长,细胞受害程度增强,影响了细胞内蛋白质等物质合成,至翌年3月30日POD活性下降,但GN1st单、混播处理较其它品种仍维持较高水平,这就减轻了膜脂过氧化程度,增加了机体的抗寒性能,之后随着气温逐渐回升,胁迫解除,各品种紫花苜蓿单、混播处理根系POD活性又逐渐升高,10月15日至4月30日,同一观测期内各品种单、混播处理之间差异显著(P<0.05)。
2.6不同品种苜蓿单、混播后抗寒评价
对3种紫花苜蓿单播及与其草地羊茅混播抗寒性的研究发现,混播的抗寒性高于单播,但品种之间比较单播抗寒性高的品种混播处理后抗寒性仍然较高,然而抗寒性高的品种也不是每一项生理指标都表现为最高,为了减少单一指标对抗寒性评价引起的误差,本研究采用了Fuzzy数学中隶属函数法对供试单、混播的苜蓿品种的各项生理生化指标进行综合分析(表5),结果表明,其抗寒性依次为GN1st+CD>GN1st>Wega7F+CD>Wega7F>WL319HQ+CD>WL319HQ。
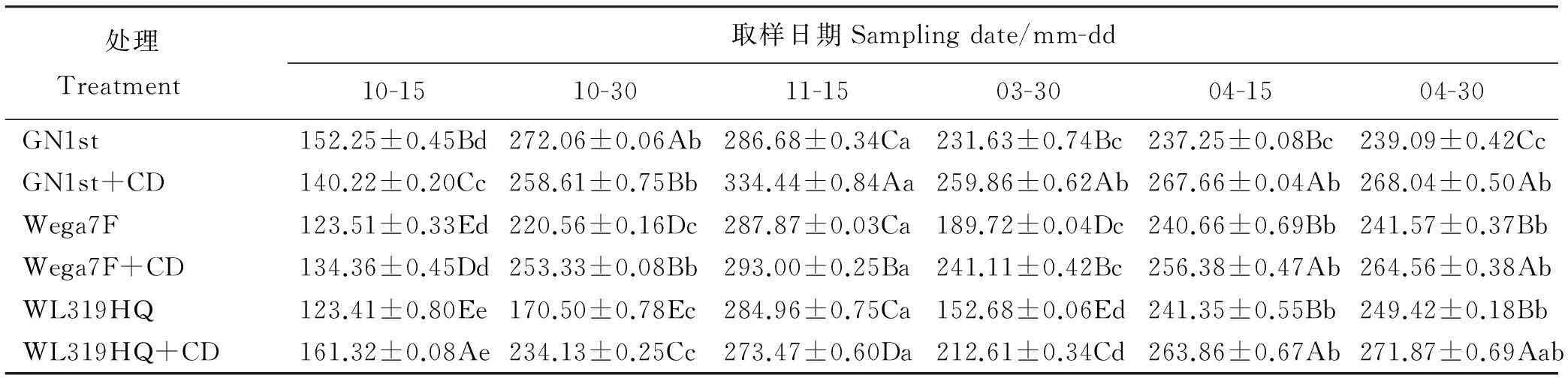
表4 不同处理紫花苜蓿根系过氧化物酶活性(OD470·min-1·g-1)变化

表5 3个苜蓿品种单播及与草地羊茅混播根系生理生化指标的隶属函数值
3讨论
植物体内可溶性糖含量的增加能增强胞内溶质浓度,降低细胞的冰点,增强持水力,降低胁迫对细胞的伤害,有报道称苜蓿主根中可溶性糖的浓度在环境温度较低的情况下明显增加,但在翌年苜蓿返青时下降[21];这与本研究中不同苜蓿品种单、混播根内可溶性糖含量变化情况基本一致,但本研究中的WL319HQ单、混播越冬率比供试其它品种低,而根系可溶性糖含量在11月15日高于Wega7F单播,且差异达到显著水平(P<0.05),可能不同品种对不同生态区适应性不同所致,因此,可溶性糖含量不能单独作为评价苜蓿抗寒性高低的生理指标[22]。
研究表明,可溶性蛋白质和脯氨酸的累积与植物的抗寒性并不存在直接相关关系[23-24],而本研究中各单播处理及单、混播处理之间比较,可溶性蛋白含量呈现出明显的季节动态,且在整个冬天维持较高含量。此外,不同品种之间可溶性蛋白含量在同一时期内积累量不同,在不同时期不同品种之间单、混播处理存在差异,另外,本研究发现引进品种的可溶性蛋白含量低于国内品种。游离脯氨酸含量作为判别苜蓿抗寒指标一直是人们争论的热点问题,很多研究均表明,游离脯氨酸可以作为指示植物抗寒性强弱的生理指标。低温胁迫条件下,游离脯氨酸含量随着温度的下降而上升,并随着胁迫时间的延长而增加,并已证明植物体内游离脯氨酸的积累有利于提高其抗寒性[25-27]。本研究中无论是单播还是混播,苜蓿根系游离脯氨酸含量均是从10月中旬开始,随着气温的逐渐下降而上升,引进品种WL319HQ在10月末游离脯氨酸含量达到峰值,其它品种及处理则是在11月中旬达到最大值,翌年苜蓿返青,WL319HQ根系游离脯氨酸含量较其它品种下降至最低,可能是由于引进品种WL319HQ对温度较为敏感,对生态环境的适应程度与国内品种不同所致。
POD酶活性与植物抗寒性呈正相关关系,POD酶活性越强,植物抗氧化能力越强,则对低温的适应性越强[28]。此外,POD在酶促防御系统中的主要作用是分解H2O2、阻止膜脂过氧化,从而抵御代谢过程中有害物质对细胞的毒害,其活力的变化可作为植物的耐寒指标[29-32]。本研究中不同苜蓿品种之间及单混播之间根系POD活性均呈现先增加后降低然后再增加的变化趋势,不同苜蓿单、混播处理在同一时期内差异显著(P<0.05)。10月末随季节气温逐渐下降,为了有效防止活性氧对膜脂和蛋白质的过氧化作用,根系中POD的活性开始加强,随着温度的进一步下降,至11月中旬,POD活性升至最高,之后随温度的上升又开始下降,此时可溶性糖、可溶性蛋白和游离脯氨酸等含量开始逐渐下降,细胞渗透调节能力降低,POD的活性也随之减弱,气温变暖后,解除胁迫,POD活性又开始增强,苜蓿开始返青,可溶性糖、可溶性蛋白及游离脯氨酸等指标含量开始下降,此时各处理苜蓿根系中营养由库端变成源端[31]。
已有研究表明,植物的抗寒性评价结果受不同生理生化指标的综合影响,本研究中各处理紫花苜蓿在低温胁迫下的可溶性糖含量、可溶性蛋白含量、游离脯氨酸含量及POD活性与综合评价值呈显著正相关,说明这几项生理生化指标可作为各单、混播处理紫花苜蓿抗寒性鉴定指标,来综合评价其抗寒性[4]。本研究应用隶属函数法对3种紫花苜蓿单播及与草地羊茅混播的抗寒性进行综合评价,结果表明,低温胁迫下,同一处理并不是在所有的指标方面均表现良好,而是在某个或某几个指标方面表现较好,所以说单凭某一指标不能评价苜蓿抗寒性强弱,应采用多项指标综合评判。3种紫花苜蓿单播及分别与草地羊茅混播后其抗寒性由大到小顺序为GN1st+CD>GN1st>Wega7F+CD>Wega7F>WL319HQ+CD>WL319HQ。另外,研究发现,在此比例混播组合下,两种牧草之间竞争小,个体和群体之间能够协调发展,提高了空间光能和土壤的肥料利用效率,竞争使苜蓿根颈入土深度更深,混播除了能够更好搭配营养结构和充分利用时间和空间的养分外,混播较单播还有助于提高其越冬和抗寒性能。
4结论
3 个不同苜蓿品种与草地羊茅单、混播越冬期根系可溶性糖、可溶性蛋白和游离脯氨酸含量均随越冬期气温降低而提高, 翌年春又随气温的回升而下降;而POD活性则是先随着气温下降而上升,进入最冷月开始下降,翌年返青后随气温回升又逐渐上升,综合各项指标结果表明,同一观测期内各指标含量不同,各生理指标在进入最冷月份(11月15日)单、混播处理均达到差异显著水平(P<0.05)。应用 Fuzzy数学中隶属函数法对供试各处理苜蓿根系生理生化指标进行综合分析[27],其抗寒性由大到小依次为GN1st+CD>GN1st>Wega7F+CD>Wega7F>WL319HQ+CD>WL319HQ,与越冬率的调查结果相一致。参考文献(References)
[1]罗新义,冯昌军,李红,沙伟.低温胁迫下肇东苜蓿SOD、脯氨酸活性变化初报.中国草地学报,2004,26(4):79-81.
Luo X Y,Feng C J,Li H,Sha W.Study on changes of SOD and proline activity during low temperature stress onMedicagosativaL.cv.Zhaodong.Chinese Journal of Grassland,2004,26(4):79-81.(in Chinese)
[2]邓雪柯,乔代蓉,李良,于昕,张乃胜,雷高鹏,曹毅.低温胁迫对紫花苜蓿生理特性影响的研究.四川大学学报(自然科学版),2005,42(1):190-194.
Deng X K,Qiao D R,Li L,Yu X,Zhang N S,Lei G P,Cao Y.The effect of chilling stress on physiological characters ofMedicagosativa.Journal of Sichuan University(Natural Science Edition),2005,42(1):190-194.(in Chinese)
[3]Brumme C E,Moore K J,Bjork C N.Agronomic consequences of dormant-non dormant alfalfa mixtures.Agronomy Journal,2002,94(4):782-789.
[4]南丽丽,师尚礼,陈建纲,朱新强,郭全恩,赵文翰.不同根型苜蓿根系对低温胁迫的响应及其抗寒性评价.中国生态农业学报,2011,19(3):619-625.
Nan L L,Shi S L,Chen J G,Zhu X Q,Guo Q E,Zhao W H.Field evaluation of the response and resistance to low temperature of alfalfa root with different root types during over-wintering.Chinese Journal of Eco-Agriculture,2011,19(3):619-625.(in Chinese)
[5]梁慧敏,夏阳,梁月香.碳水化合物含量和过氧化物酶酶性变化与苜蓿抗寒性的关系.甘肃农业大学学报,1995,30(4):307-311.
Liang H M,Xia Y,Liang Y X.Seasonal changes in carbo hydrate content and peroxidase activities related to cold hardiness of alfalfa.Journal of Gansu Agricultural University,1995,30(4):307-311.(in Chinese)
[6]梁慧敏,夏阳.苜蓿抗寒性及根蘖性状的表现与过氧化物酶同工酶关系的研究.草业学报,1998,7(4):55-60.
Liang H M,Xia Y.Relationship between isoperoxidase and expression of cold-tolerance in creep-rooted alfalfa.Acta Prataculturae Sinica,1998,7(4):55-60.(in Chinese)
[7]史纪安,刘玉华,韩清芳,贾志宽.不同秋眠级数的紫花苜蓿品种根系发育能力研究.西北农业学报,2009,18(4):149-154.
Shi J A,Liu Y H,Han Q F,Jia Z K.Root development ability of different fall dormancy alfalfa cultivars.Acta Agriculturae Boreali-Occidentalis Sinica,2009,18(4):149-154.(in Chinese)
[8]孙启忠,桂荣,韩建国.赤峰地区敖汉苜蓿冻害及其防御技术.草地学报,2001,9(1):50-57.
Sun Q Z,Gui R,Han J G.Research on winter injury and prevent technique of alfalfa (MedicagosativaL.cv.‘Aohan’)in Chifeng Region.Acta Agrestia Sinica,2001,9(1):50-57.(in Chinese)
[9]Small E,Jomphe M A,Synopsis of the genusMedicago(Legominosae).Canadian Journal of Botany,1998,67:3260-3294.
[10]王磊,李建勇,张振贤,葛均青.冻害低温下越冬甘蓝渗透调节物质的变化和作用.山东农业大学学报(自然科学版),2001,32(4):487-490.
Wang L,Li J Y,Zhang Z X,Ge J Q.Role and change of osmotic adjustment of winter cabbage under freezing tolerance.Journal of Shandong Agricultural University (Natural Science),2001,32(4):487-490.(in Chinese)
[11]Cunningham S M,Gana J A,Volenec J J.Winter hardiness,root physiology,and gene expression in successive fall dormancy selections from ‘Mesilla’ and ‘CUF101’ alfalfa.Crop Science,2001,41:1091-1098.
[12]孟凡珍,张振贤,于贤昌,艾希珍.田间低温胁迫对大白菜某些理化特性的影响研究.中国生态农业学报,2005,13(2):84-86.
Meng F Z,Zhang Z X,Yu X C,Ai X Z.Effect of low temperature stress on some physiological and biochemical characteristics of Chinese cabbage in field.Chinese Journal of Eco-Agriculture,2005,13(2):84-86.(in Chinese)
[13]周瑞莲,张普金.春季高寒草地牧草根中营养物质含量和保护酶活性的变化及其生态适应性研究.生态学报,1996,16(4):402-407.
Zhou R L,Zhang P J.The change in carbohydrate content and protecting enzyme activities in root of alpine forage related to its adaptation to cold in spring.Acta Ecologica Sinica,1996,16(4):402-407.(in Chinese)
[14]Garaham D,Patterson B D.Responses of plants to low,nonfreezing temperatures:Proteins,metabolism,and acclimation.Annual Review of Plant Physiology,1982,33:347-372.
[15]包乌云,赵萌莉,徐军,高新磊,赵巴音那木拉.苜蓿与禾本科牧草的混播效果.草业科学,2013,30(11):1782-1789.
Bao W Y,Zhao M L,Xu J,Gao X L,Zhao B.Study on mixculture of alfalfa with grasses.Pratacultural Scinence,2013,30(11):1782-1789.(in Chinese)
[16]祁亚淑,朱林,许兴.宁夏绿洲禾豆牧草混播组合及其比例效应.草业科学,2015,32(9):1463-1472.
Qi Y S,Zhu L,Xu X.Effects of mixed combination and proportion between legume inous and graminaceous forages grown in Ningxia desert steppe.Pratacultural Scinence,2015,32(9):1463-1472.(in Chinese)
[17]高婷婷,毛培春,田小霞,孟林,刘汉云.10个白三叶品种苗期耐寒性评价.草业科学,2014,31(9):1724-1731.
Gao T T,Mao P C,Tian X X,Meng L,Liu H Y.Cold tolerance assessment of 10Trifoliumrepenscultivars at the seedling stage.Pratacultural Scinence,2014,31(9):1724-1731.(in Chinese)
[18]张治安,张美善,蔚荣海.植物生理学实验指导.北京:中国农业科学技术出版社,2004.
Zhang Z A,Zhang M S,Wei R H.Plant Physiology Experiment Guiding.Beijing:China Agricultural Science and Technology Press,2004.(in Chinese)
[19]邹琦.植物生理学实验指导.北京:中国农业出版社,2000:8.
Zou Q.Plant Physiology Experiment Guiding.Beijing:China Agriculture Press,2000:8.(in Chinese)
[20]申晓慧,姜成,冯鹏,李如来,李建东,郑海燕.寒区6个紫花苜蓿品种根系中MDA含量及抗氧化酶活性的比较研究.作物杂志,2015(4):88-91.
Shen X H,Jiang C,Feng P,Li R L,Li J D,Zheng H Y.Comparison of MDA content and antioxidant enzymes activity of several alfalfa roots in cold region.Crops,2015(4):88-91.(in Chinese)
[21]刘香萍,李国良,崔国文.紫花苜蓿抗寒性研究进展.饲料博览,2006(12):11-13.
Liu X P,Li G L,Cui G W.The Research progress of cold resistance on alfalfa.Feed Review,2006(12):11-13.(in Chinese)
[22]王运涛,于林清,萨仁.苜蓿抗寒性研究进展.草原与草坪,2012,32(3):91-96.
Wang Y T,Yu L Q,Sa R.Research progress of alfalfa cold resistance in China.Grassland and Turf,2012,32(3):91-96.(in Chinese)
[23]刘祖祺,张石城.植物抗性生理学.北京:中国农业出版社,2014.
Liu Z Q,Zhang S C.Plant Resistance Physiology.Beijing:China Agriculture Press,2014.(in Chinese)
[24]刘娥娥,宗会,郭振飞,黎用朝.干旱盐和低温胁迫对水稻幼苗脯氨酸含量的影响.热带亚热带植物学报,2000,8(3):235-238.
Liu E E,Zong H,Guo Z F,Li Y C.Effects of drought,salt and chilling stresses on proling accumulation in shoot of rice seedlings.Journal of Tropical and subtropical Botany,2000,8(3):235-238.(in Chinese)
[25]冯昌军,罗新义,沙伟,王凤国.低温胁迫对苜蓿品种幼苗SOD、POD活性和脯氨酸含量的影响.草业科学,2005,22(6):29-32.
Feng C J,Luo X Y,Sha W,Wang F G.Effect of low temperature stress on SOD,POD activity and proline content of alfalfa.Pratacultural Science,2005,22(6):29-32.(in Chinese)
[26]陈雅君,崔国文,富象乾.低温对苜蓿品种幼苗体内游离脯氨酸含量的影响.中国草地学报,1996(6):46-50.
Chen Y J,Cui G W,Fu X Q.The effect of low temperature on free proline content in different alfalfa seedlings.Chinese Journal of Grassland,1996(6):46-50.(in Chinese)
[27]南丽丽,师尚礼,朱新强,郭全恩.田间越冬期不同根型苜蓿根系的生理生化特性.核农学报,2011,25(2):369-374.
Nan L L,Shi S L,Zhu X Q,Guo Q E.Physiological and biochemical characteristics of root in different root type alfalfa cultivars in field during overwintering period.Journal of Nuclear Agricultural Sciences,2011,25(2):369-374.(in Chinese)
[28]江福英,李延,翁伯琦.植物低温胁迫及抗旱生理.福建农业学报,2002,17(3):190-195.
Jiang F Y,Li Y,Weng B Q.Review on physiology of chilling stress and chilling resistance of plants.Fujian Journal of Agricultural Sciences,2002,17(3):190-195.(in Chinese)
[29]Mckerise B D,Murnaghan J,Jones K S.Iron-superoxide dismutase expression in transgenic alfalfa increases winter tolerance.Plant Physiology,2000,122:1427-1437.
[30]杜永吉,于磊,孙吉雄,鲁为华,韩烈宝.结缕草品种抗寒性和抗寒机理研究.草地学报,2008,16(4):347-352.
Du Y J,Yu L,Sun J X,Lu W H,Han L B.Study on cold-resistance and its mechanism of different cultivars ofZoysiajaponicaSteud.Acta Agrestia Sinica,2008,16(4):347-352.(in Chinese)
[31]鲁剑巍,邹娟,周世利,蒋细旺,丛红霞,徐艳丽.施磷对越冬期高羊茅生长、养分吸收及抗寒性的影响.草地学报,2008,16(5):436-441.
Lu J W,Zou J,Zhou S L,Jiang X X,Cong H X,Xu Y L.Effect of phosphorus application amount on the growth,nutrient absorption,and cold-resistance of tall fescue (FestucaarundinaceaSchreb.)in winter.Acta Agrestia Sinica,2008,16(5):436-441.(in Chinese)
[32]Wolf S,Marani A,Rudieh J.Effect of temperature on carbohydrate metabolism in potato plants.Journal of Experimental Botany,1991,238(42):619-625.
(责任编辑张瑾)
The study on the cold resistance and physiology change of root in meadow fescue━alfalfa mixture and monoculture in winter
Shen Xiao-hui1,2, Jiang Cheng3, Li Ru-lai1, Li Jian-dong2, Zhang Hua1,Zheng Hai-yan1, Guo Wei1, Sun Li1, Feng Peng1
(1.Jiamusi Branch of Heilongjiang Academy of Agricultural Sciences, Jiamusi 154007,China;2.Agricultural College, Shenyang Agricultural university, Shenyang 110866, China;3.College of Life Science, Jiamusi University, Jiamusi 154007, China)
Abstract:In order to improve cold resistance of alfalfa (Medicago sativa), three alfalfa cultivars (Gongnong1st(GN1st), the Wega7F and the WL319HQ ) and one meadow fescue (Festuca ovina) cultivar were used in this study. Soluble sugar, soluble protein, free proline and POD activity, and winter survival rate were measured at six different time. The results showed that the winter survival rate were different among different varieties, the average winter survival rate in mixture was more than 90%,better than monoculture,and the winter survival rate in the each field had significant difference between mixture and monoculture. Variations in soluble sugar, soluble protein, free proline contents increased along with temperature decrease in late fall, then decreased along with temperature increase in spring of the following year, soluble sugar content reached peaked in mid-November, and there were significant different between monoculture and mixture (P<0.05); soluble protein content reached peaked at the end of October, there were significant difference between monoculture and mixture (P<0.05), except WL319HQ; free proline content of WL319HQ reached peak at the end of October, other treatments reached peaked in mid-November, there were significant difference between monoculture and mixture in the same measure stage (P<0.05); the POD activity change showed up-down-up trend in the whole wintering stage, showed the highest activity in mid-November. Membership function analysis and winter survival rate showed the order of cold-resistance of alfalfa cultivars was GN1st+meadow fescue>GN1st>Wega7F+meadow fescue>Wega7>WL319HQ+meadow fescue>WL319HQ.
Key words:alfalfa; meadow fescue; mixture; cold resistance; winter survival rate; physiology index
Corresponding author:Feng PengE-mail:fengras@163.com
中图分类号:S816;S540.34;S344.1+4
文献标识码:A
文章编号:1001-0629(2016)2-0268-08
通信作者:冯鹏(1980-)男,内蒙化德人,副研究员,博士,主要从事牧草育种及栽培研究。E-mail:fengras@163.com
基金项目:黑龙江省政府博士后资助项目 (LBH-Z12239);中国博士后科学基金(2013M541423)
*收稿日期:2015-06-29接受日期:2015-10-09
DOI:10.11829/j.issn.1001-0629.2015-0365
申晓慧,姜成,李如来,李建东,张华,郑海燕,郭伟,孙力,冯鹏.3种紫花苜蓿与草地羊茅单、混播越冬期根系生理变化及抗寒性.草业科学,2016,33(2):268-275.
Shen X H,Jiang C,Li R L,Li J D,Zhang H,Zheng H Y,Guo W,Sun L,Feng P.The study on the cold resistance and physiology change of root in meadow fescue-alfalfa mixture and monoculture in winter.Pratacultural Science,2016,33(2):268-275.
第一作者:申晓慧(1980-)女,吉林扶余人,助理研究员,在读博士生,主要从事牧草抗寒性研究。E-mail:xiaohuishen@126.com
猜你喜欢
草原与草坪(2022年1期)2022-05-12
草业学报(2022年2期)2022-02-15
西藏农业科技(2019年1期)2019-07-25
吉林农业·下半月(2017年1期)2017-02-16
江苏农业科学(2016年8期)2017-02-15
安徽农学通报(2017年1期)2017-02-15
物联网技术(2016年12期)2017-01-21
现代经济信息(2016年7期)2016-05-19
天津农业科学(2016年4期)2016-04-20
杂草学报(2015年2期)2016-01-04
