
祁连山不同植被类型的物候变化及其对气候的响应
2016-04-14 02:19贾文雄俎佳星陈京华
生态学报 2016年23期
贾文雄,赵 珍,俎佳星,陈京华,王 洁,丁 丹
1 西北师范大学地理与环境科学学院,兰州 730070 2 中国科学院沈阳应用生态研究所森林与土壤生态国家重点实验室,沈阳 110016
祁连山不同植被类型的物候变化及其对气候的响应
贾文雄1,*,赵 珍1,俎佳星2,陈京华1,王 洁1,丁 丹1
1 西北师范大学地理与环境科学学院,兰州 730070 2 中国科学院沈阳应用生态研究所森林与土壤生态国家重点实验室,沈阳 110016
基于1982—2006年GIMMS NDVI和2000—2014年MODIS NDVI遥感数据,利用double logistic拟合方法提取了1982—2014年祁连山区不同植被的生长季始期、生长季末期和生长季长度3个重要的物候参数,分析了不同植被物候期的时间变化趋势、空间分异特征及对气候因子的响应。结果表明:(1)祁连山区不同植被的生长季始期和生长季末期随年际变化表现出波动提前或推迟,其中沼泽植被的变化波动最大;草甸植被、灌丛植被、阔叶林植被和栽培植被生长季长度出现延长趋势;(2)祁连山区植被生长季始期集中在5月初,其中阔叶林植被生长季开始最早,荒漠植被生长季开始最晚,植被生长季末期集中在9月,栽培植被生长季结束较早,荒漠植被、沼泽植被生长季结束较晚,植被生长季长度集中在110—140 d,其中阔叶林植被、针叶林植被生长季长度较长,而荒漠植被、高山植被生长季长度较短;(3)祁连山植被物候期变化趋势的空间分布表明植被生长季始期、生长季末期主要表现为提前不明显和推迟不明显,生长季长度主要表现为缩短不明显和延长不明显;(4)物候要素与气候要素相关性表明前期温度的积累有利于植被的开始生长,但当年3月的降水量对植被生长季始期同样有重要作用,不同植被生长季末期与8月、9月温度相关性较大,而与10月、11月降水的相关性较大。
物候参数;不同植被类型;时空变化;气候响应;祁连山区
物候学是研究自然界以年为周期重复出现的各种生物现象与环境因子的周期性变化相互关系的科学[1-2]。植物是陆地生态系统的主体,植物物候指植物受气候和其他环境因子的影响而出现的以年为周期的自然现象[3]。植被物候(包括生长季开始时间、结束时间、生长季长度)不仅能指示植被的节律变化,同时反映了植被年际变化对气候变化的响应,是全球变化的“积分仪”和景观生态环境变化的综合“指示器”[4]。IPCC第五次评估报告指出,1880—2012年全球地表平均温度上升约0.85℃,全球变暖毋庸置疑。随着全球气候变化,植被物候也发生显著的变化,如Tucker等[5]发现随着温度的升高,35°N以上地区的植被生长季有明显的延长趋势;Sparks等[6]提出英国地区因气温升高2.5℃导致开花时间提前了5—25 d;郑景云等[7]提出东北、华北等地春季平均温度升高导致物候期提前,而长江中游、华南等地春季温度下降造成物候期推迟。当前全球变化研究中最为活跃的“全球变化与陆地生态系统(Global Change and Terrestrial Ecosystems,GCTE)”项目[8]对植被物的研究越来越重视。
传统的物候研究方法以野外观测为基础,需耗费大量人力、物力,而且研究范围面积小,周期短且受时间限制。在计算机、遥感技术的推动下,使得遥感监测不同区域的植被物候并进行长时间、大范围的研究成为可能。Karlsen等[9]基于GIMMS NDVI数据分析得出北欧地区的植物生长季提前与温度有关;Delbart等[10]利用NOAA AVHRR和SPOT NDVI数据分析了西伯利亚地区的植被物候,得到生长季始期在1982—1991年提前了7.8 d,而在2000—2004年却延迟了7 d,呈现的这种波动状态与温度的波动性保持一致;刘玲玲等[11]基于GIMMS AVHRR NDVI数据,对欧亚大陆植被物候变化趋势进行监测分析,得出大部分植被类型的生长季开始时间呈提前趋势,其中森林提前幅度最大;余振等[12]利用NOAA/AVHRR NDVI研究了我国东部南北样带不同植被的物候期,指出温带草丛、亚热带热带草丛等植被返青期显著提前,寒温带、温带针叶林等植被休眠期显著推迟,大部分植被的生长季长度均出现延长趋势;王宏等[13]利用NOAA/AVHRR NDVI数据研究了我国北方植被生长季变化,得出大部分纬度的植被生长季始期提前,生长季末期推迟;李明等[14]利用SPOT NDVI数据得出长白山区林地生长季开始时间早于草地、耕地,结束时间却晚于草地、耕地;国志兴等[15]利用GIMMS NDVI数据得到东北地区沼泽、针叶林、阔叶林、草丛、草甸植被生长季延长,而农田、灌丛、草原、针阔混交林生长季缩短。张戈丽和张扬建等[16]发现利用1982—2000年GIMMS和2001—2011年SPOT—VGT合并提取的青藏高原1982—2011年植被物候期开始时间出现提前趋势,趋势为1.04 d/a,甚至有的地区提前时间达到近1个月左右,表明青藏高原植被生态系统对全球变化有着明显的正响应。国内利用遥感数据研究植被物候变化主要集中于东北地区,而对于西北地区遥感监测植被物候期的研究却很少。
祁连山区是西北地区重要的生态区,已有学者利用遥感数据对祁连山植被NDVI变化、植被覆盖变化及其与气候变化关系等方面作了深入研究[17- 19],但对于遥感监测分析该地区植被物候期的研究极少。本研究基于GIMMS NDVI和MODIS NDVI数据集,利用Savitzky—Golay滤波重构NDVI时间序列数据,并用double logistic拟合方法提取了不同植被的物候期(生长季开始日期、结束日期和生长季长度),在此基础上分析了祁连山1982—2014年不同植被的物候期时间变化趋势、空间分异特征及与气候因子温度、降水的相关性,以期全面认识在气候变化影响下研究区近33年来不同植被的物候期变化状况,进而在生产中根据不同植被物候期对气候变化的响应具体安排农、林业生产和生活,从而为农业生产计划、合理轮牧以及生态环境建设提供科学依据,并且有利于进一步理解大尺度区域上气候对不同植被物候期变化的影响。
1 研究区概况
祁连山(93°30′—103°00′E,35°43′—39°36′N)位于青藏高原的东北部,地跨青海、甘肃两省,由多条西北-东南走向的平行山脉和宽谷组成,是西北地区著名的高大山系之一,其地理位置及祁连山的植被分布如图1所示。祁连山是青藏高原、内蒙古高原和黄土高原的分界线,自然条件复杂,水热条件差异大,东部湿度大,降水多,西部干燥,降水少,具有典型的大陆性气候特征,气候还表现出多样性及垂直变化特征,如山前低山-中山下部-中山上部-亚高山及高山地区年降水约为150、250—300、400—500、800 mm;年均温约为6、2—5、0—1、-5℃。由于起伏不一的地形和复杂的水热条件,祁连山区植被类型丰富,主要有草甸、草原、荒漠、灌丛、高山植被等植被,分别占祁连山总面积的30.14%、25.94%、14.61%、9.29%、7.90%,其他植被零星分布。研究祁连山植被物候的变化趋势,能为区域尺度的可持续发展提供一定的指导研究。

图1 祁连山植被分布及气象站点图Fig.1 The distribution of vegetation and meteorological station of Qilian Mountains
2 数据来源
2.1 遥感数据
本研究所采用的遥感数据是GIMMS AVHRR NDVI和MODIS NDVI 数据集。GIMMS AVHRR NDVI数据集是由美国国家海洋与大气管理局(The National Oceanic and Atmospheric Administration,简称NOAA)推出的全球植被指数变化数据,可以在中国西部环境与生态科学数据中心(http://westdc.westgis.ac.cn)免费下载,时间范围为1982—2006年。MODIS NDVI 数据集采用NASA提供的MODIS13A2级植被指数产品,可在网站(https://wist.echo.nasa.gov/api/)直接下载,时间范围为2000—2014年。应用MRT(MODIS Reprojection Tools)软件对MODIS NDVI数据进行数据格式、投影转换和数据拼接等。
利用ENVI 4.7 工具和祁连山区边界矢量数据对下载的遥感数据分别进行裁剪,得到研究区的1982—2014年NDVI时间序列数据。NDVI值应在[-1,1]之间,区间以外的数值代表其它地物,如水体、裸地等。
由于采用两种数据,根据2000—2006年重叠的7a数据,对数据进行一致性检验(表1),由表可知,2000—2006年GIMMS NDVI数据和MODIS NDVI数据相关系数均在0.9以上,说明两种时间序列数据具有较好的一致性,可以同时用于植被物候的研究中。
2.2 其他数据
气候数据为1982—2014年祁连山及其周边地区共33个气象站点(图1)的日平均气温和降水量数据,资料来源于中国气象科学数据共享服务网(http://cdc.cma.gov.cn)。植被类型图为寒区旱区科学数据中心提供的1∶1000000矢量图。

表1 2000—2006年GIMMS NDVI和MODIS NDVI数据相关分析
**表示通过0.01的置信度检验
3 研究方法
3.1 Savitzky-Golay滤波法
为了进一步降低云层、大气等对NDVI时间序列的影响,必须对数据进行滤波平滑[11]。目前滤波平滑的方法有很多,如Savitzky-Golay滤波法、时间序列分析法(HANTS)、滑动平均法、中值迭代滤波法(MIF)、最小二乘法等[20- 24],其中S-G滤波法对传感器类型、NDVI尺度等没有严格的要求,且得到的重构数据质量较高[25],所以本文采用S-G滤波法进行NDVI数据的滤波平滑。
S-G滤波法是由Savitzky和Golay提出的一种最小二乘卷积拟合平滑滤波方法,广泛用于数据的平滑除噪。S-G滤波法应用在NDVI曲线滤波时,公式可表示为:
(1)
式中,Y为原始NDVI值,Y*为滤波拟合后的NDVI值,j指数据序列的第j个点,Ci为第i个数据点的滤波系数。N为滑动窗口的宽度即2m+1[26- 27]。
运用S-G滤波法分别对祁连山GIMMS NDVI和MODIS NDVI数据集进行了滤波平滑,得到平滑的NDVI时间序列,为祁连山不同植被物候要素的提取做好准备。
3.2 物候要素提取
目前,对于遥感图像的物候要素提取,国内外学者已经提出了多种监测方法,如阈值法,导数法,模型拟合法[9,28- 29]等。阈值法操作简单但受人为因素影响较大,影响了结果的准确性;导数法也限制于经验阈值;模型拟合法所拟合的精度直接影响物候参数的确定。本文采用导数和阈值方法结合的拟合方法来进行物候参数的提取,拟合公式double logistic[30]为:
(2)
式中,NDVIt表示在t时间的NDVI值;NDVImin表示年NDVI最小值;NDVIdiff表示年NDVI差值;ri表示NDVI左侧即上升时拐点的最大变化率;nSOS为理论上的生长季始期;rd表示NDVI在右侧即下降时拐点的最大变化率;nEOS为理论上的生长季末期。
采用该函数对滤波后的年NDVI时间序列的每个像元进行拟合,然后对函数求二阶导,定义NDVI二阶导数变化率最大的点为生长季始期(SOS);函数在下降的过程中NDVIt值达到年NDVI最大值的80%,即0.8×(NDVImin+NDVIdiff)定义为生长季末期(EOS);生长季末期与始期的差值为生长季长度(LOS)。运用该物候要素提取方法,得到GIMMS和MODIS数据集下的祁连山不同植被的物候要素(SOS、EOS、LOS)。
3.3 线性趋势分析
为了研究祁连山区每个栅格的物候要素变化趋势,采用一元线性回归分析法对其进行模拟,计算公式[31]为:
(3)
式中,n表示时间序列的长度;i表示年序列号;Ti为植被物候期在第i年的生长季始期、末期或生长季长度。θSlope指的是趋势线的斜率,若θSlope>0,说明物候期变化趋势是延迟或延长,反之θSlope<0,则说明物候期变化趋势是提前或缩短。此外,根据T检验对计算结果进行分类,将变化趋势分为不明显、明显、显著等类型。
3.4 相关分析法
通过地理要素之间的相关分析可以揭示其相互之间的响应关系,利用SPSS软件得到相关系数,并用此研究不同植被物候期与气温、降水间的关系,相关系数大于零,表示物候期随因子变化呈推迟趋势,相关系数小于零,表示物候期随因子变化呈提前趋势;相关系数的绝对值越大,说明植被物候期的变化与因子变化相关性越高;绝对值越小,则说明两者的相关性越低。
4 结果分析
4.1 不同植被类型的物候期时间变化
将1982—2014年祁连山区不同植被每年的物候期(生长季始期、生长季末期、生长季长度)进行一元线性回归分析,得到33a物候期变化趋势(图2,图3,表2)。
1982—2006年草甸植被、沼泽植被的生长季始期呈推迟趋势,变化速率(斜率)分别为0.023 d/a、0.285 d/a;草甸植被生长季末期呈推迟趋势,斜率为0.03 d/a,相比而言,生长季末期的推迟趋势比生长季始期的推迟趋势稍微明显,因此生长季长度主要因为生长季末期的推迟而呈延长趋势,但趋势不明显,方差为51.14;而沼泽植被生长季末期呈提前趋势,斜率为0.285 d/a,方差为150.79;生长季长度呈缩短趋势,方差为393.14;表明沼泽植被生长季始期、生长季末期、生长季长度的年际波动变化都很大。1982—2006年草原植被、荒漠植被、高山植被的生长季始期、末期均呈提前趋势,但趋势不一,生长季始期的斜率分别为0.031、0.045、0.252 d/a,生长季末期的斜率分别为0.107、0.329、0.272 d/a;3种植被的生长季长度却出现缩短,其中荒漠植被方差

为118.11,年际波动极大。1982—2006年灌丛植被、栽培植被、针叶林、阔叶林的生长季始期均呈提前趋势,斜率分别为0.054、0.013、0.002、0.306 d/a,生长季末期均呈推迟趋势,斜率分别为0.137、0.089、0.149、0.385 d/a,生长季长度呈延长趋势,年际波动不大。

图3 不同植被生长季长度的年际变化Fig.3 Annual change trends of the length of growth season of different type vegetations in Qilian Mountains

植被类型Vegetationtypes1982—2006年GIMMS数据集GIMMSNDVIof1982—20062000—2014年MODIS数据集MODISNDVIof2000—2014生长季始期Startofgrowingseason生长季末期Endofgrowingseason生长季长度Lengthofgrowingseason生长季始期Startofgrowingseason生长季末期Endofgrowingseason生长季长度Lengthofgrowingseason草甸y=0.023x+141.74y=0.0295x+250.44y=0.0065x+108.7y=-0.5168x+148.57y=-0.1194x+267.91y=0.3974x+119.34草原y=-0.0312x+138.13y=-0.1065x+251.22y=-0.0753x+113.09y=-0.155x+142.35y=-0.1394x+270.81y=0.0156x+128.46荒漠y=-0.0446x+147.36y=-0.3286x+256.67y=-0.284x+109.31y=-0.0756x+147.78y=-0.6837x+274.99y=-0.6081x+127.2灌丛y=-0.0537x+133.87y=0.1371x+248.73y=0.1908x+114.86y=-0.4021x+140.87y=-0.1442x+270.94y=0.2579x+130.07高山植被y=-0.2524x+148.48y=-0.2723x+254.47y=-0.0199x+105.99y=-0.6941x+142.85y=-0.0038x+264.05y=0.6903x+121.2针叶林y=-0.0015x+130.01y=0.1485x+246.83y=0.15x+116.82y=-0.0856x+136.96y=-0.2533x+271.91y=-0.1677x+134.95阔叶林y=-0.3063x+127.07y=0.3845x+243.32y=0.6908x+116.25y=-0.2889x+133.17y=-0.1191x+272.91y=0.1698x+139.74栽培植被y=-0.013x+131.31y=0.0889x+246.22y=0.1019x+114.91y=-0.3426x+144.11y=0.1707x+263.54y=0.5133x+119.43沼泽植被y=0.85x+133.59y=-0.2846x+260.98y=-1.1292x+127.36y=-0.148x+149.04y=0.0061x+278.42y=0.1541x+129.38
草甸 Meadow, 草原 Steppe, 荒漠 Desert vegetation, 灌丛 Shrub vegetation, 高山植被 Alpine vegetation, 针叶林 Coniferous forest, 阔叶林 Broad-leaved forest, 栽培植被 Cultivated vegetation, 沼泽植被 Swamp vegetation
2000—2014年草甸植被、草原植被、灌丛植被、高山植被、阔叶林的生长季始期、末期均出现提前;生长季始期的斜率分别为0.517、0.155、0.402、0.694、0.289 d/a;生长季末期的斜率分别为0.119、0.139、0.144、0.004、0.119 d/a;生长季长度主要受生长季始期提前趋势的影响而呈延长趋势,其中草甸植被方差为53.38,年际波动小。2000—2014年荒漠植被、针叶林植被的生长季始期、末期亦出现提前趋势,但生长季长度却出现缩短;生长季始期的斜率分别为0.076、0.086 d/a,生长季末期的斜率分别为0.6837、0.253 d/a,其中荒漠植被生长季长度年际波动大,方差为126.77。栽培植被、沼泽植被的生长季始期呈提前趋势,斜率分别为0.343、0.148 d/a;生长季末期呈推迟趋势,斜率分别为0.171、0.006 d/a;但两种植被的生长季长度呈延长趋势。
总体来看,祁连山区植被生长季长度受生长季开始时间提前(推迟)或生长季结束时间推迟(提前)等的影响,表现出延长或缩短趋势。从图3、表2可以看出草甸植被、灌丛植被、阔叶林植被和栽培植被在1982—2014年生长季长度总体上延长,而荒漠植被生长季长度却出现缩短,且植被的年际波动变化较大;1982—2006年草原植被、高山植被、沼泽植被生长季长度缩短,其中沼泽植被的年际波动较大,而生长季长度在2000—2014年却出现延长趋势,表明在近十几年内这些植被生长季长度延长;1982—2006年针叶林植被生长季长度呈延长趋势,而2000—2014年生长季长度呈缩短趋势,且年际波动较小。
4.2 不同植被类型的物候期空间变化
4.2.1 不同植被平均物候期的空间分布
由图4a得到1982—2006年祁连山阔叶林植被生长季开始最早,主要集中于第120天以前,其次是针叶林植被和栽培植被,生长季始期主要集中在第120—140天,灌丛植被和草原植被生长季始期主要集中在第120—150天,草甸植被生长季始期主要集中在第130—150天,沼泽植被生长季始期主要集中在第140—150天,高山植被生长季始期主要集中在第130—160天,荒漠植被生长季开始时间最晚,主要在第140天以后。图4b中祁连山1982—2006年针叶林植被、阔叶林植被、栽培植被生长季结束时间较早,集中在第240—250天内,草甸植被、草原植被、灌丛植被、高山植被生长季末期主要集中在第240—260天,沼泽植被的生长季末期主要集中在第250—260天,而荒漠植被生长季结束时间较晚,主要集中在第240—270天。植被生长季长度取决于生长季的始期和末期,由1982—2006年祁连山各植被生长季始期、末期得到图4c,从图中看出祁连山荒漠植被、高山植被生长季长度较短,集中在80—120 d,沼泽植被生长季长度分布比较分散,在80—140 d之间,草甸植被、草原植被、栽培植被生长季长度主要集中在100—120 d,灌丛植被、针叶林植被生长季长度则主要集中在100—140 d,而阔叶林植被生长季长度主要集中在120—140 d。

图4 1982—2006年祁连山植被平均物候期及变化趋势的显著性分布Fig.4 The spatial variation trend and its significance of vegetation average phenology in Qilian Mountains from 1982 to 2006图4a、4b、4c分别为平均物候期的始期、末期、生长季长度;图4d、4e、4f分别为平均物候期的始期、末期、生长季长度变化的显著性
图5a是祁连山2000—2014年不同植被平均生长季始期分布情况,得出阔叶林植被生长季开始时间较早,且主要集中在第130—140天,其次是针叶林植被、栽培植被、灌丛植被,生长季始期主要集中在第130—150天,草原植被生长季始期集中在第130—160天,草甸植被、沼泽植被生长季始期集中在第140—160天,荒漠植被、高山植被生长季始期跨度较大,主要在第130天以后。图5b中2000—2014年栽培植被、高山植被、草甸植被、荒漠植被生长季末期主要集中在第255—275天,草原植被生长季末期主要集中在第255—285天,针叶林植被、灌丛植被生长季末期主要集中在第265—275天,阔叶林植被生长季末期主要集中在第265—285天,沼泽植被生长季结束较晚,集中于第275—285天。图5c表示祁连山2000—2014年植被的生长季长度,可以得出荒漠植被、高山植被、草甸植被、草原植被、栽培植被生长季长度主要集中在110—130 d,沼泽植被、灌丛植被、针叶林植被生长季长度主要集中在120—140 d,阔叶林植被生长季长度主要集中在130—160 d。
由于2000—2014年的遥感数据分辨率高,识别植被绿度的能力增强,从而较之GIMMS NDVI数据得到的植被生长季开始时间较早,结束日期较晚,但两种数据得到的不同植被生长季开始先后和生长季结束先后的趋势基本一致。

图5 2000—2014年祁连山植被平均物候期及变化趋势的显著性分布Fig.5 The spatial variation trend and its significance of vegetation average phenology in Qilian Mountains from 2000 to 2014图5a、5b、5c分别为平均物候期的始期、末期、生长季长度;图5d、5e、5f分别为平均物候期的始期、末期、生长季长度变化的显著性
4.2.2 植被物候期多年变化的空间分布
从图4d、图5d得出,1982—2006年、2000—2014年总体祁连山植被生长季始期提前不明显和推迟不明显的区域面积最大,分别为95.87%、77.22%。不同植被生长季始期的年际变化有所不同,但极大多数表现为各自的提前不明显区域所占比例大于推迟不明显。1982—2006年草甸植被、草原植被、高山植被、灌丛植被、阔叶林、栽培植被、针叶林、荒漠植被生长季始期提前不明显的比例分别为49.59%、47.44%、60.82%、56.89%、64.29%、56.76%、52.27%、50.8%,推迟不明显比例分别为49.36%、45.12%、31.96%、42.76%、35.71%、43.24%、47.73%、36.9%。2000—2014年草甸植被、草原植被、高山植被、灌丛植被、阔叶林、栽培植被、针叶林始期提前不明显的比例分别为55.82%、45.79%、38.01%、67.56%、77.32%、51%、60.17%,推迟不明显比例分别为25.57%、33.58%、27.76%、22.93%、20.57%、36.11%、29.31%,荒漠植被始期提前不明显比例小于推迟不明显比例,分别为28.20%、28.89%。
从图4e、图5e得出,1982—2006年、2000—2014年总体祁连山植被生长季末期提前不明显和推迟不明显区域所占比例极大,分别为95.68%、90.87%。1982—2006年草甸植被、草原植被、灌丛植被、阔叶林、栽培植被、针叶林生长季末期表现为推迟不明显比例大于提前不明显,推迟不明显的比例分别为63.51%、47.44%、65.02%、78.57%、56.76%、70.45%,提前不明显的比例分别为35.32%、46.20%、34.28%、21.43%、43.24%、27.27%;高山植被、荒漠植被末期提前不明显的区域占52.06%、59.89%,明显提前占5.15%、5.35%,显著提前占2.58%、2.67%,推迟不明显占39.18%、31.02%。2000—2014年草甸植被、草原植被、高山植被、灌丛植被、阔叶林、针叶林、荒漠植被生长季末期表现为提前不明显比例大于推迟不明显,提前不明显的比例分别为64%、53.86%、51.02%、57.03%、51.20%、65.38%、49.61%,推迟不明显比例分别为32.99%、37.77%、36.98%、41.34%、48.23%、32.62%、22.28%;而栽培植被末期提前不明显的占44.49%,推迟不明显占49%。
从图4f、5f得出,1982—2006年、2000—2014年总体祁连山植被生长季长度年际变化缩短不明显和延长不明显区域所占比例极大,分别为95.15%、70.26%。1982—2006年草甸植被、草原植被、高山植被、灌丛植被、阔叶林、栽培植被、针叶林生长季长度表现为延长不明显比例大于缩短不明显,延长不明显的比例分别为58.25%、53.02%、48.45%、64.31%、64.29%、54.05%、65.91%,缩短不明显的比例分别为40%、40.93%、41.75%、35.34%、35.71%、45.95%、31.82%;而荒漠植被缩短不明显占51.34%,延长不明显仅占37.43%。2000—2014年草甸植被、草原植被、高山植被、灌丛植被、阔叶林、栽培植被、针叶林生长季长度表现为延长不明显比例大于缩短不明显,延长不明显的比例分别为42.11%、37.10%、31.15%、51.82%、60.86%、44.18%、45.25%,缩短不明显比例分别为34.38%、35.04%、26.50%、31.76%、34.93%、32.68%、38.32%,而荒漠植被生长季长度缩短不明显的为27.01%,延长不明显仅占20.05%。
1982—2014年祁连山植被物候期(始期、末期、长度)年际变化整体变化不明显,主要表现为提前不明显、推迟不明显或缩短不明显、延长不明显。同样不同植被类型的物候期年际变化趋势中的提前(缩短)不明显、推迟(延长)不明显所占面积比例极大。
4.3 物候期要素与气候因子关系
基于祁连山区及其周边33个气象站点的气象数据,得到1982—2014年祁连山区年均温呈上升趋势,斜率为0.0426 ℃/a,年平均气温为5.7 ℃,年均温最高出现在1998年,最低出现在1984年,分别为6.6、4.5 ℃;年降水量也呈上升趋势,斜率为0.8663 mm/a,平均年降水量为225 mm,年降水量最高为282.5 mm(2007年),最低为174.5 mm(1991年)。利用相关分析方法分析了植被物候期要素与气温、降水的关系,从而探究了不同植被物候对气象因子的响应(表3,表4)。
由表3得出植被生长季始期与2—5月各月气温的相关性均未通过显著检验。其中,沼泽植被的生长季始期与2—5月各月气温呈正相关,温度升高,蒸发加快,使土壤表层湿度不够,沼泽植被生长受到限制,所以沼泽植被的生长季始期随温度升高推迟。除沼泽植被外的其他植被生长季始期与2—4月各月气温呈负相关,即随温度不断升高,植被提前发芽,生长季始期提前。草原植被、荒漠植被、栽培植被生长季始期与5月气温呈正相关,其他植被生长季始期与5月气温负相关。植被生长季始期与2—5月降水的相关性较高,这与邓少福[19]研究得到的降水对祁连山草地植被返青影响更大的结果一致。其中沼泽植被的生长季始期与2月降水呈显著正相关,且通过了0.05置信度检验;各植被生长季始期与3月的降水均呈负相关,即降水增多,植被生长季始期提前。其中,高山植被生长季始期与3月降水的相关性通过了0.05置信度检验,草甸植被、草原植被、荒漠植被、灌丛植被、针叶林植被、栽培植被生长季始期与3月降水的相关性通过了0.01置信度检验,且相关系数较高;其他植被的生长季始期与各月降水都没有通过显著性检验。
由表4得出荒漠植被生长季末期与8月气温呈负相关,且通过了0.01置信度检验,随温度的升高,蒸发量加大,植被水分减少,导致提前发黄干枯;高山植被生长季末期与8—11月气温呈负相关,且与8月气温显著负相关并通过了0.05的置信度检验,随温度升高,高山植被提前枯黄;其他植被的生长季末期与各月气温相关性都没有通过显著性检验。各植被生长季末期与8月降水均呈正相关,即降水量增加,植被枯黄的时间推迟,其中草甸植被、栽培植被生长季末期与8月降水显著正相关,且都通过0.05置信度检验;高山植被生长季末期与9月降水呈负相关,通过0.05置信度检验;植被生长季末期与10月、11月降水相关性均没有通过显著性检验。

表3 祁连山植被生长季始期与气温、降水的相关系数
*表示通过0.05的置信度检验,**表示通过0.01的置信度检验
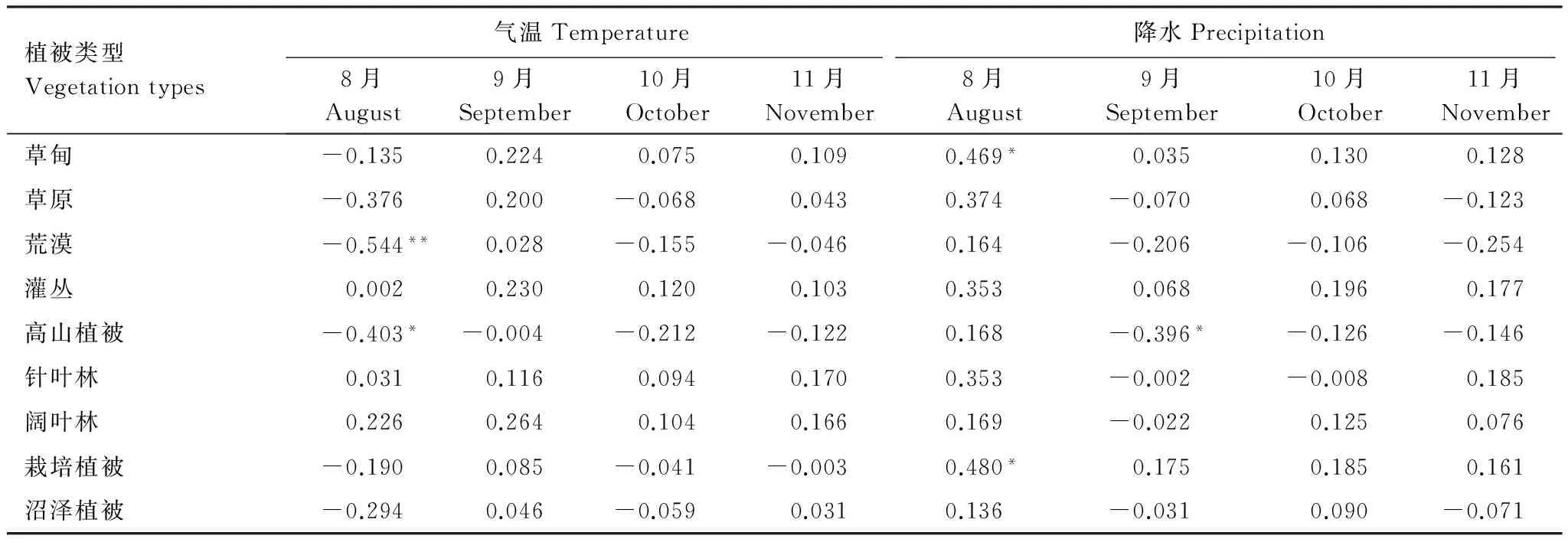
表4 祁连山植被生长季末期与气温、降水的相关系数
*表示通过0.05的置信度检验,**表示通过0.01的置信度检验
4.4 物候期要素与海拔的关系
图6显示了2000—2014年祁连山植被物候参数在不同海拔高程上的变化情况。从图中看出,当海拔在2700 m以下时,随着海拔的升高,植被生长季始期有提前的趋势;而当海拔在2700—3250 m之间时,随着海拔的升高,生长季始期有较明显的推迟趋势;当海拔大于3250 m时,随着海拔的升高,生长季开始时间出现缓慢推迟。总体上,祁连山植被的生长季始期随海拔变化的一元线性方程的斜率为0.0095,即生长季开始时间随海拔每升高1000 m推迟9.5 d,而生长季始期随海拔的升高呈推迟趋势的情况与物候期的空间分布情况一致,且与徐珂等[32]关于植被生长季开始时间与海拔关系的研究结果一致。
植被生长季末期随海拔的变化呈现出较大的波动变化,当海拔低于3250 m时,随着海拔的升高,植被生长季末期表现出比较明显的推迟趋势;而当海拔在3250—4100 m之间时,随着海拔的升高,生长季末期则表现出提前的趋势;而在4100—4500 m间,随着海拔的升高植被生长季末期出现推迟;当海拔超过4500 m时,生长季末期再次随海拔升高出现提前趋势。
随海拔的升高植被生长季长度整体呈缩短趋势,这与物候期的空间分布情况一致。植被生长季长度受海拔变化的影响较小,其与海拔变化的线性斜率为—0.0096,即海拔每升高1000 m植被生长季长度缩短9.6 d。研究结果与王力等[33]研究所得的海拔升高,生长期缩短的结果保持一致。
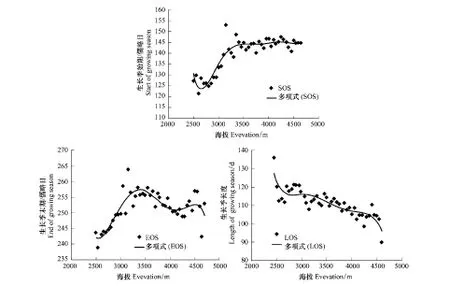
图6 不同海拔上物候参数的变化情况Fig.6 Variations of phenology parameter in different elevations
5 结果与讨论
本文利用Double logistic拟合方法提取并分析了研究区不同植被的物候参数时空分异特征,并结合同期气象数据,探讨了植被物候期对气候因子的响应。结果如下:
(1)1982—2014年祁连山草甸植被、灌丛植被、阔叶林植被和栽培植被生长季长度延长,而荒漠植被生长季长度却出现缩短;草原植被、高山植被、沼泽植被在1982—2006年生长季长度缩短,而在2000—2014年生长季长度却出现延长;针叶林植被在1982—2006年生长季长度延长,而在2000—2014年生长季长度缩短。
(2)祁连山各植被生长季始期主要在5月初,其中阔叶林植被生长季开始时间最早,荒漠植被生长季始期出现的最晚。各植被生长季末期主要在9月,栽培植被生长季结束较早,荒漠植被、沼泽植被生长季结束较晚。各植被生长季长度集中于110 d—140 d,其中阔叶林植被、针叶林植被生长季长度较长,而荒漠植被、高山植被生长季长度较短。
(3)各植被物候期在1982—2014年年际变化趋势不明显,生长季始期、末期主要表现为提前不明显、推迟不明显,所占面积比例极大,生长季长度主要表现为缩短不明显、延长不明显,所占面积比例极大。
(4)祁连山植被生长季始期前期温度的积累有利于植被的生长,而当年3月的降水也是影响植被生长季始期的重要因子。植被生长季末期与8—11月各时段的气温、降水相关性不明显,各植被生长季末期与8—9月温度相关性较大,而与10—11月降水的相关性较大。
植被物候变化不完全是气候因素造成的,但是实际调查中,气候因素的变化为植被物候变化提供了极大地可能性。植被生长季始期同样受温度影响,由于前期温度的积累,使得植物生长季始期所需的温度达到标准后,而此时对于干旱、半干旱地区降水因子却成了影响植被是否生长的关键因子,这与Xin等[34]得到的水分对干旱、半干旱地区植被返青影响更大的结果一致。不同高程的植被物候参数对气候变化的响应程度有所不同,海拔较低的阔叶林生长季始期受温度变化影响较大,而其他海拔较高的植被受水分变化的影响较大,这是由于海拔较低地区的阔叶林生长期始期早于海拔较高地区的植被,气温逐步回升或快速升高对其影响较大,而在气温全面回升后高海拔植被开始萌发,降水的多寡对其生长有明显的影响。不同植被类型生长季末期受温度和水分影响的关系较复杂,但高海拔地区的植被受温度影响更明显,这是因为在不同海拔高度上水热条件的不同组合导致不同植被类型生长季结束的时间并不一致,总体上随着气温的快速下降导致高海拔地区的植被进入生长季末期。本文只研究了气温、降水变化对其物候变化的影响,但其它气候因子、地形因子、土壤因子等对植被物候变化也有一定的影响,它们之间的关系有待于进一步研究。
2000—2006年,GIMMS AVHRR NDVI和MODIS NDVI数据重叠,这两种数据线性拟合的相关系数均在0.9以上,说明两种时间序列数据具有较好的一致性,可以同时用于植被物候变化的研究中。根据门源站实测数据植被生长季始期为第138天,结束期为第247天,从两套数据中得到的生长季始期分别为第131、130天,结束日期分别为第246、250天,两者结果基本一致。在已有的研究成果中,李娜[35]得出石羊河流域草地平均开始时间为第124天,结束时间为第266天,这与本文研究结果也基本一致。可见,利用遥感提取数据进行植被物候研究具有一定的准确度。张扬健等[16]基于1982—2000年GIMMS NDVI及2001—2011年的SPOT VGT遥感数据合并提取了1982—2011年青藏高原的植被物候,研究也指出遥感提取及多种数据合并监测植被物候期具有一定的参考价值和可行性。本文采用两种遥感数据分不同时段提取植被的物候参数,但如何将不同时空分辨率的数据融合,利用长时间序列数据开展物候变化研究将是未来研究的重要方面。
[1] 竺可桢, 宛敏渭. 物候学. 长沙: 湖南教育出版社, 1999: 1- 4.
[2] Lieth H. Phenology and Seasonality Modeling. Berlin Heidelberg: Springer-Verlag, 1974.
[3] 陆佩玲, 于强, 贺庆棠. 植物物候对气候变化的响应. 生态学报, 2006, 26(3): 923- 929.
[4] Li X B, Chen Y H, Fan Y D, Zhang Y X. Detecting inter-annual variations of vegetation growth based on satellite-sensed vegetation index data from 1983 to 1999 // Proceedings of IEEE International Geoscience and Remote Sensing Symposium. Toulouse, France: IEEE, 2003, 5: 3263- 3265.
[5] Tucker C J, Slayback D A, Pinzon J E, Los S O, Myneni R B, Taylor M G. Higher northern latitude normalized difference vegetation index and growing season trends from 1982 to 1999. International Journal of Biometeorology, 2001, 45(4): 184- 190.
[6] Sparks T H, Jeffree E P, Jeffree C E. An Examination of the relationship between flowering times and temperature at the national scale using long-term phenological records from the UK. International Journal of Biometeorology, 2000, 44(2): 82- 87.
[7] 郑景云, 葛全胜, 郝志新. 气候增暖对我国近40年植物物候变化的影响. 科学通报, 2002, 47(20): 1582- 1587.
[8] 周广胜, 张新时, 高素华, 白克智, 延晓东, 郑元润. 中国植被对全球变化反应的研究. 植物学报, 1997, 39(9): 879- 888.
[9] Karlsen S R, Solheim I, Beck P S A, Høgda K A, Wielgolaski F E, Tømmervik H. Variability of the start of the growing season in Fennoscandia, 1982—2002. International Journal of Biometeorology, 2007, 51(6): 513- 524.
[10] Delbart N, Toan T L, Kergoat L, Fedotova V. Remote sensing of spring phenology in boreal regions: a free of snow-effect method using NOAA-AVHRR and SPOT-VGT data (1982—2004). Remote Sensing of Environment, 2006, 101(1): 52- 62.
[11] 刘玲玲, 刘良云, 胡勇. 1982—2006年欧亚大陆植被生长季开始时间遥感监测分析. 地理科学进展, 2012, 31(11): 1433- 1442.
[12] 余振, 孙鹏森, 刘世荣. 中国东部南北样带主要植被类型物候期的变化. 植物生态学报, 2010, 34(3): 316- 329.
[13] 王宏, 李晓兵, 李霞, 莺歌, 符娜. 基于NOAA NDVI和MSAVI研究中国北方植被生长季变化. 生态学报, 2007, 27(2): 504- 515.
[14] 李明, 吴正方, 杜海波, 宗盛伟, 孟祥君, 张莲芝. 基于遥感方法的长白山地区植被物候期变化趋势研究. 地理科学, 2011, 31(10): 1242- 1248.
[15] 国志兴, 张晓宁, 王宗明, 方伟华. 东北地区植被物候期遥感模拟与变化规律. 生态学杂志, 2010, 29(1): 165- 172.
[16] Zhang G L, Zhang Y J, Dong J W, Xiao X M. Green-up dates in the Tibetan Plateau have continuously advance-d from 1982 to 2011. Proceedings of the National Academy of Sciences of the United States of America, 2013, 110(11): 4309- 4314.
[17] 戴声佩, 张勃, 王海军, 王亚敏, 李丹, 王兴梅. 1999—2007年祁连山区植被指数时空变化. 干旱区研究, 2010, 27(4): 585- 591.
[18] 武正丽. 2000—2012年祁连山植被覆盖变化及其对气候的响应研究[D]. 兰州: 西北师范大学, 2014.
[19] 邓少福. 祁连山气候变化对植被的影响研究(2000—2011)[D]. 兰州: 兰州大学, 2013.
[20] Savitzky A, Golay M J E. Smoothing and differentiation of data by simplified least squares procedures. Analytical Chemistry, 1964, 36(8): 1627- 1639.
[21] Roerink G, Menenti M, Verhoef W. Reconstructing cloudfree NDVI composites using Fourier analysis of time series. International Journal of Remote Sensing, 2000, 21(9): 1911- 1917.
[22] Malingreau J P. Global vegetation dynamics: satellite observations over Asia. International Journal of Remote Sensing, 1986, 7(9): 1121- 1146.
[23] Kogan F, Sullivan J. Development of global drought-watch system using NOAA/AVHRR data. Advances in Space Research, 1993, 13(5): 219- 222.
[24] 侯英雨, 王石立. 基于作物植被指数和温度的产量估算模型研究. 地理学与国土研究, 2002, 18(3): 105- 107.
[25] Bian J H, Li A N, Song M Q, Ma L Q, Jiang J G. Reconstruction of NDVI time-series datasets of MODIS based on Savitzky-Golay filter. Journal of Remote Sensing, 2010, 14(4): 725- 741.
[26] Chen J, Jönsson P, Tamura M, Gu Z H, Matsushita B, Eklundh L. A simple method for reconstructing a high-quality NDVI time-series data set based on the Savitzky-Golay filter. Remote Sensing of Environment, 2004, 91(3/4): 332- 344.
[27] 侯学会, 牛铮, 高帅, 黄妮. 基于SPOT-VGT NDVI时间序列的农牧交错带植被物候监测. 农业工程学报, 2013, 29(1): 142- 150.
[28] Zhang X Y, Friedl M A, Schaaf C B, Strahler A H, Hodges J C F, Gao F, Reed B C, Huete A. Monitoring vegetation phenology using MODIS. Remote Sensing of Environment, 2003, 84(3): 471- 475.
[29] Fisher J I, Mustard J F. Cross-scalar satellite phenology from ground, Landsat, and MODIS data. Remote Sensing of Environment, 2007, 109(3): 261- 273.
[30] Butt B, Turner M D, Singh A, Brottem L. Use of MODIS NDVI to evaluate changing latitudinal gradients of rangeland phenology in Sudano-Sahelian West Africa. Remote Sensing of Environment, 2011, 115(12): 3367- 3376.
[31] 宋怡, 马明国. 基于GIMMS AVHRR NDVI数据的中国寒旱区植被动态及其与气候因子的关系. 遥感学报, 2008, 12(3): 499- 505.
[32] 徐珂. 基于MODIS影像的北京地区生长季及其影响因子研究[D]. 北京: 林业大学, 2012.
[33] 王力, 李凤霞, 周万福, 李晓东. 气候变化对不同海拔高山嵩草物候期的影响. 草业科学, 2012, 29(8): 1256- 1261.
[34] Xin Q C, Broich M, Zhu P, Gong P. Modeling grassland spring onset across the Western United States using climate variables and MODIS-derived phenology metrics. Remote Sensing of Environment, 2015, 161: 63- 77.
[35] 李娜. 1999—2006年石羊河流域植被对气候变化的响应研究[D]. 兰州: 兰州大学, 2010.
Phenological variation in different vegetation types and their response to climate change in the Qilian Mountains, China, 1982—2014
JIA Wenxiong1,*, ZHAO Zhen1, ZU Jiaxing2, CHEN Jinghua1, WANG Jie1, DING Dan1
1CollegeofGeographyandEnvironmentScience,NorthwestNormalUniversity,Lanzhou730070,china2StateKeyLaboratoryofForestandSoilEcology,InstituteofAppliedEcology,ChineseAcademyofSciences,Shenyang110016,China
Vegetation phenology is used in the study of climate change because of stable distribution, and ease of observation and interpretation. In this study, a 15-day time series of averaged normalized difference vegetation indices (NDVI) derived from the daily Global Inventory Modeling and Mapping Studies (GIMMS) NDVI dataset and 16-day averaged NDVI values derived from the daily Moderate Resolution Imaging Spectroradiometer (MODIS) NDVI dataset were used to analyze trends in vegetation phenology. Firstly, using the ENVI tool to cut remote sensing data, NDVI time series data for the study area from 1982 to 2014 were obtained. Secondly, using a Savitzky-Golay filter, noise contamination caused by random factors was reduced by producing a smooth NDVI curve. Thirdly, using double logistic fitting, three important phenological parameters, including the start of growing season (SOS), the end of growing season (EOS), and the length of growing season (LOS) were extracted for different vegetation types. Temporal change trends, as well as its spatial distribution characteristics, of the nine major vegetation types in the Qilian Mountains were analyzed. These types included meadow, steppe, desert vegetation, shrub vegetation, alpine vegetation, coniferous forest, broad-leaved forest, cultivated vegetation, and swamp vegetation. In addition, the effects of climatic factors on phenology were analyzed by correlation analysis. The results showed that: (1) The annual variation of SOS and EOS of different vegetation types in the Qilian Mountains fluctuated in its advance or or delay; the maximum variation was observed in the swamp vegetation. The LOS of meadow, shrub, coniferous forest, and cultivated vegetation was longer, but the LOS of desert vegetation shortened. (2) The SOS of vegetation was primarily in May; the growing season of the broad-leaved forests began the earliest and desert vegetation began the latest. The EOS of vegetation occurred primarily in September; the growing season of cultivated vegetation ended earlier, whereas the growing season of the desert and swamp vegetation ended later. The LOS of vegetation was from 110 days to 140 days, among which the LOS of broad-leaved and coniferous forests was longer, whereas the LOS of desert and alpine vegetation was shorter. (3) The spatial distribution of variation trends for vegetation phenology indicated that SOS and LOS were advanced or delayed but not substantially, and the LOS was primarily shortened or prolonged, but not substantially. (4) The correlation of phenological metrics and climatic factors indicated that the accumulation of early stage temperature was beneficial to the growth of vegetation, but the amount of precipitation in March was also important to the SOS of vegetation. The EOS of different vegetation types was related with temperature in August and September, and correlated with precipitation in October and November, but the correlation was not significant. At different altitudes, the phenological parameters were different. In particular, the LOS appeared to shorten with increasing elevation, which is consistent with the spatial distribution of phenological parameters. These results explain the relationship between climate change and phenological phases. Climate change has an obvious effect on vegetation phenological parameters, and the phenological period can explicitly indicate the climate change.
phenological parameters; vegetation types; spatial-temporal variations; climate change; Qilian Mountains
国家自然科学基金资助项目(41161017)
2015- 10- 09;
2016- 06- 12
10.5846/stxb201510092039
*通讯作者Corresponding author.E-mail: wxjiaxy@163.com
贾文雄,赵珍,俎佳星,陈京华,王洁,丁丹.祁连山不同植被类型的物候变化及其对气候的响应.生态学报,2016,36(23):7826- 7840.
Jia W X, Zhao Z, Zu J X, Chen J H, Wang J, Ding D.Phenological variation in different vegetation types and their response to climate change in the Qilian Mountains, China, 1982—2014.Acta Ecologica Sinica,2016,36(23):7826- 7840.
猜你喜欢
河北地质(2022年2期)2022-08-22
热带作物学报(2022年6期)2022-07-08
读者·原创版(2021年2期)2021-02-26
现代园艺(2017年23期)2018-01-18
西南农业学报(2017年5期)2017-06-23
河北林业科技(2016年5期)2016-11-08
西南农业学报(2016年5期)2016-05-17
华人时刊(2016年17期)2016-04-05
黄河之声(2016年20期)2016-02-21
应用海洋学学报(2015年2期)2015-11-22
