
再论干热河谷植被恢复过程中的“适度”造林
2016-04-11 08:12费世民蒋俊明张旭东周金星四川省林业科学研究院成都6008中国林业科学研究院林业研究所北京0009北京林业大学北京0008
四川林业科技 2016年1期
费世民,蒋俊明,张旭东,周金星(.四川省林业科学研究院,成都 6008;.中国林业科学研究院林业研究所,北京 0009;.北京林业大学,北京 0008)
再论干热河谷植被恢复过程中的“适度”造林
费世民1,蒋俊明1,张旭东2,周金星3
(1.四川省林业科学研究院,成都 610081;2.中国林业科学研究院林业研究所,北京 100091;3.北京林业大学,北京 100083)
摘要:本文在干热河谷植被恢复过程中的“适度”造林技术提出的基础上,以干热河谷的自然植被演替、土壤水分承载力、立地异质性为依据,从系统化、定量化的研究角度,在林分结构(微观)上探讨了造林“适度”密度,在区域景观尺度(宏观)上探讨植被恢复的群落“适度”配置,旨在进一步阐明“适度”造林技术与理论,以期为干热河谷植被恢复的研究与实践提供参考。
关键词:干热河谷;植被恢复;“适度”造林;“适度”密度;群落“适度”配置
人工植被目前是我国的主要森林植被类型之一。但现存的大部分人工林植被由于植被类型选择不当、群落密度过大和群落生产力过高等原因,普遍存在林地地力衰退的生态问题,在广大干旱、半干旱地区,特别是干热河谷区,植被恢复困难,林分稳定性差,以“土壤干化”为主的土壤退化极为严重,已经引起了学术界的广泛关注。
多年来,干热河谷的植被恢复策略一直以造林为主,旨在恢复或部分恢复森林植被,以现有植被起源于受人为干扰的原生森林植被或以局部地区种植林木生长状况为依据。从目前看,在传统的造林理念下,干热河谷人工造林与植被恢复,在结构、土壤等方面均表现出明显的退化特征。传统的造林技术与方法不但难以达到恢复生态功能的目的,而且还造成人力、物力和财力的巨大浪费(费世民等,2003,2004)。虽然干热河谷植被恢复重建的理论和方法方面的研究虽已有所积累,但是其理论体系和技术体系尚未形成。因此,深入研究干热河谷植被恢复的生态过程和及其机理,如何构建科学有效的造林与植被恢复理论,依然是目前十分迫切的任务。
长期以来,干热河谷被认为是世界上环境最恶劣的地区,其植被恢复一直深受重视。经长期努力,该区植被恢复在物种筛选、恢复模式、营造技术、人工植被的生态评价等方面的研究进展迅速,已筛选出一批树(草)种,营建了大面积的试验示范林。虽然部分研究已取得一些突破与进展,但干热河谷植被恢复依然还是尚未攻克的难题,由于理论贮备及技术研究的不足,还缺乏进一步的理论探索和系统研究成果,特别缺乏定量化、系统化、持续性的研究,理论体系仍不完善。(张信宝等,1997;李昆等,1999;杨忠等,2003;费世民等,2003,2004)。为此,本文在“适度”造林技术(费世民等,2003,2004)提出的基础上,旨在从系统化、定量化的研究角度,进一步阐明“适度”造林技术与理论,以期为干热河谷植被恢复的研究与实践提供参考。
1 “适度”造林的背景
在干旱、半干旱地区,由于以往造林多选用乔木,并且造林密度偏大,导致林木水分营养面积不足、土壤水分亏缺,从而引起林分衰退,甚至死亡。造林与土壤水分平衡的关系是现在和将来干热、干旱地区植被恢复研究的重点问题之一。
在北方,许多研究结果表明,固沙林下的土壤贮水量一般低于无林地,尤其是密度偏大的林分,在林分郁闭成林后,水分供应将十分紧张,会引起林木死亡,这是由于林分蒸腾耗水,导致根系分布区土壤贮水量减少,根系分布层的土壤含水量降至凋萎系数。宁夏中卫2 a~8 a生人工林地表以下0~3 m土层含水量从1988年的2%~3%降低到1990年的1% ~2%(杨维西等,1996);1964年营造的灌木和半灌木林地0~3 m土层含水量普遍在0.59%~1.0%之间,有的甚至降到0.59%以下,绝对含水量与无植被流沙区同一层次相差达60 mm~100 mm(郭普,1986)。1970年营造了樟子松固沙林,初植密度为4 444株·hm-2,1978年保存株数是4 000 株·hm-2,24 a后密度为2 500株·hm-2。由于该区降水年变率较大,1 a中降水又多集中于7月、8月份,如果樟子松固沙林密度偏大,将引起土壤水分亏缺,影响固沙林生态稳定性,甚至林木死亡。在科尔沁沙地,生长季内除差巴嘎蒿0~120 cm土层内土水势高于-15 bar以外,樟子松、小叶锦鸡儿、小叶杨林地同一土层的土水势均不同程度地处于- 15 bar(凋萎湿度)以下,已不能为林木所利用((陈荷生,1993)。在民勤绿洲和河西走廊的石羊河及黑河流域下游,由于河流补给量减少以及植物的强烈蒸腾,地下水位大幅度下降,以致干枯(杨文斌,1992),造成大片人工林和天然胡杨林及柽柳生长严重衰退.在黄土高原的许多地区,人工灌木林地土壤水分亏缺,虽然未大片死亡,但对植物已造成严重的生理伤害,生长极其缓慢(徐化成,1991)。在陕西安塞,6年生刺槐林地0.6 m~4.0 m土层也存在非常明显的水分严重亏缺层(徐化成,1991)。
金沙江干热河谷区气候干旱炎热,降水量较少且分配不均,土壤干旱贫瘠,是我国造林极端困难的立地之一。多年来,金沙江干热河谷区退化山地植被恢复重建一直被作为重要的研究课题。其中选择适宜物种,优化群落组合和配套的造林技术是植被恢复能否成功的关键因素。长期以来,植树造林一直作为几乎整个生态恢复的重要措施之一,但是效果一直不好。在干热河谷区山地有这样的说法“种树不见树,造林不见林”,许多地方出现的“小老头树”既不能发挥水土保持作用,又不产生经济效益,而且还引起以“土壤干化”为主的生态问题。
在金沙江干热河谷,20世纪50年代、70年代直播的云南松(Pinus yunnanensis)、思茅松(Pinus langbianensis)观测,干热河谷中种植的松树,第6年自然稀疏到1 050株·hm-2~6 750株·hm-2,第8年锐减到450株·hm-2~1 050株·hm-2,第13年进一步减为450株·hm-2~600株·hm-2,直到第18年后还在继续减少。随时间推移而退化为疏林或散生木,最后成为稀树草丛。攀枝花市曾在前进乡等地,采用云南松、思茅松、赤桉(Eucalyptus camaldulensis)、火绳树(Eriolaena malvacea)、合欢(Albizia julibrissin)、铁刀木(Cassiasiamea)等60个树种的营养袋造林,次年保存率为70%,但到第3、第4年,土壤水分、养分得不到满足而大量死亡。这说明:造林初期,林木水分需求量小,随着林分的郁闭和林木的生长,林木对水分需求量增大,土壤水分不能满足地上植物生长的需求,地上植物只有通过个体竞争,进行自然稀疏,达到自我调节的目的。
干热河谷人工赤桉林随林分密度的增大,土壤水环境有进一步恶化的趋势。13 a生的林分旱季0 ~100 cm土层内平均含水率6.84%~9.16%,土壤含水率普遍偏低,已均无有效水分,表现出严重的“土壤干化”(王克勤等,2004)。在较好立地条件的地带,10 a~13 a生的赤桉纯林,造林保存率达80%,在立地条件差的地带,4 a生的赤桉纯林,造林保存率仅为43%;在较好立地条件的地带,11 a生的新银合欢纯林,造林保存率72%,而在立地条件差的地带,11 a生的新银合欢纯林,造林保存率仅有45%(高文学等,2005)。据调查,1990年至1996年,攀枝花干热河谷地区造林1 333余hm2,20年后保存率不足10%。
水分条件是金沙江干热河谷植被恢复的一个主要制约因素,水分因子是影响植物生存、生长发育和环境对植被支持力的关键因素。上世纪70年代以来,采用提水灌溉的方法,攀枝花市在花岗岩荒山上成功营造了攀枝花公园、东区公园和攀钢后山绿化区,其植被以阔叶林为主的常绿森林植被。主要树种有羊蹄甲、台湾相思、桉树、山麻柳、栎类等。这3处森林内,均布设有输水灌溉管道系统。据初步调查,苗期旱季每月浇水3次,以后浇水次数逐渐减少,成林后每年3月~5月,每月浇水1次,灌溉用水量差别为180 m3·hm-2·a-1。这3处人工林均抽提金沙江水灌溉,水费高昂。攀钢后山绿化区67 hm2人工林,每年水费和维护费高约40万元;目前,由于停止供水,已开始死亡。2000年,长江造林局在三堆子附近,抽提金沙江水,营造128 hm2人工林,水利设施投资120万元,0.94万元·hm-2。可见,大面积采用提灌造林恢复植被显然是不现实的,经济上是不可行的。
因此,干热河谷植被恢复应该在区域生态环境大背景下,应坚持科学、合理、适度、可持续的原则,不应该盲目地追求数量和经济效益,试图在短期内使稀树灌丛草坡变成森林,并提供木材产品,而忽视了其严酷的自然条件,超越了干热地区的生物容量和土壤承载能力。虽然不惜一切代价,可以种植一些树木,但最终得不偿失。
2 “适度”造林的理论依据
2.1自然植被演替
自然植被是经过漫长的地质运动、气候变迁和植物群落演化而形成。在干旱、半干旱地区,由于环境对植被的支持力较小,形成了独特的区域性植被。干热河谷植被具有独特的群落外观与区系组成,系世界植被中萨王纳植被的干热河谷残存者,属河谷型萨王纳植被(Savanna of valley type),所以是我国珍稀濒危的植被类型之一(金振洲等,2000)。
金振洲等(2000)研究表明,干热河谷植被多为“稀树灌木草丛”,以中草和禾草草丛为背景构成大片草地植被,在草丛之上散生2 m~5 m为主的稀散乔木和0.5 m~2 m为主的稀散灌木,人为干扰下可成为“稀树草丛”、“稀灌草丛”和草丛外貌。群落结构上,多数分乔、灌、草3层或灌、草两层。群落种类多为热带性或热带起源的耐旱种类,热带种、温带种和中国特有种分别占47.59%、14.88%和37.42%,具有长期适应干热河谷的群落特征种或区系标志种。优势或常见种多数为生态适生种或耐于热种类。草丛优势种如扭黄茅(Heteropogon corttortus(L.)Beauv.)、孔颖草(Bothriochloa pertusa(L.)A.Camus.)、双花草(Dichanthium annulatum(Forsk.)Stapf.)等,稀树灌木如滇榄仁(Terminalia franchetii Gagnep.)、石山羊蹄甲(Bauhinia comosa Craib.)、坡柳(Dodonaea viscose Jacg.)、疏序黄荆(Vitex negundo L.f.var.laxipaniculata Pei.)等。调查资料表明,1957年元谋干热河谷区植被已为稀树灌草丛(周麟,1996)。
在干热河谷区,原始自然植被为南亚热带干性常绿阔叶林,非石灰岩山地为锥连栎林;石灰岩山地为铁橡栎林,格里坪一带石灰岩山地分布有苏铁林;沟菁、阴坡等土壤水分条件较好的地段为黄栎林。攀枝花干热河谷现代自然植被以南亚热带干热河谷稀树灌丛草坡为主要类型,草本植物以扭黄茅占绝对优势;灌木主要为车桑子、牛筋条、余甘子等;乔木主要为滇合欢、栎类。人为干扰少、土壤水分条件较好的地段,如阴坡(特别是土层深厚的缓坡和裂隙发育的石质坡地)和沟菁一带,乔木密度大,部分地段成林,多为栎树林。历史时期以来的植被演变是人类活动和气候变化的叠加结果。史前时期植被的自然演化,主要受制于气候的变化。中更新世以来,地壳隆升,河流强烈下切,岭谷高差增大,河谷焚风效应增强,干热化是数十万年来金沙江河谷气候变化的总体趋势。由于植被群落的抵抗效应,植被的演化往往滞后于气候的变化。人类活动,特别是近代人类活动对攀枝花市视野区植被演替的影响是不争的事实。数百年前,区内以栎类为主的南亚热带干性常绿阔叶林广布。森林植被砍伐后,自然植被迅速退化为干热河谷稀树草灌。森林植被破坏,土壤侵蚀加剧,土层变薄变瘠,持水性能降低,土壤水分状况变差,不再适宜原生森林植被的繁衍,人类活动加速了自然植被由南亚热带干性常绿阔叶林向干热河谷草灌植被演替的进程。目前大部分坡地的自然环境,已很难恢复原始南亚热带干性常绿阔叶林植被,只能恢复干热河谷稀树灌草植被。
2.2土壤水分植被承载力与林分稳定性
承载力(Carrying Capacity)属于物理学的范畴,它是用以控制或限制发展的一个常用概念,是可持续发展的核心问题,最早进行承载力研究的是群落生态学,它是指在一定环境条件下,单位面积某种生物个体存在数量的最大值(曲仲湘等,1983)。土壤水分植被承载力(Vegetation Carrying Capacity of Soil Water,VCCSW)是土地植被承载力(简称植被承载力)的一个特殊类型。植被承载力是指一定环境条件下单位面积某种生物个体存在数量的最大值(曲仲湘等,1983)。当森林密度接近承载力时,林木胸高断面积停止增加(MANION P.D.,D.H.GRIFHN,2001)。维持森林、灌丛和草地生态系统稳定的土壤水分来自天然降雨和地下水,地下水也来自于降水。常年雨量大且季节分配合理,或是地下水位较高的地区,水分不是植物生长的限制因子。降雨量稀少、土壤水分补给能力有限的地区,土壤水分是植物生长的限制因子。在干旱、半干旱地区,土壤水分是决定植物生长的限制因子。该类地区的植被承载力实质上由土壤水分决定。
土壤水分植被承载力是土壤水分承载植物的最大负荷,是指在较长时期(1年~多年)内,在现有的条件下,当植物根系可吸收和利用土层范围内土壤水分消耗量等于或小于土壤水分补给量时,所能维持特定植物群落健康生长的最大密度,即雨水资源中补给土壤的部分水量所能维持植物健康生长的最大数量。在雨养条件下,一定时期内森林植被系统的土壤水分补给和消耗构成一个微循环系统。在一个较长时期内,雨水补给土壤水分过程和土壤水分消耗过程一般是交互进行、相互影响的。一方面森林植物生长发育影响土壤水分补给深度和补给量,从而影响土壤水分状况和对植物的有效性;另一方面,土壤水分消耗与森林植物生长密切相关。二者是一个有机的统一体。
人工林植被的稳定性与功能发挥必须建立在有适合水资源供应的合理结构基础上,维持林分稳定性的实质是要保持一定时期内系统的动态平衡,其基本条件之一是保持一定时期内土壤的水量平衡。水是土壤—植被—大气连续系统(SPAC)的纽带。在SPAC传输体中,森林植被生态系统的土壤水分与植物生长相互影响,缺一不可。在一定立地条件下,某种森林植被群落能够生长发育,那么客观上存在这样一个密度,它能使一个时期内生态系统内土壤水分的补给和消耗达到平衡且植物生长正常,有利于生态系统稳定,也不会形成土壤旱化,此时的密度即为土壤水分植被承载力。由于在一个生长季内,植物的生长和发育由发芽、展叶,开花、果实膨大和落叶(落叶树种)等环节构成,无论哪个环节遇到干旱、土壤水分胁迫都可能导致植物个体的死亡和种群数量的下降。一年内,土壤水分在降雨、径流、土壤蒸发和植物蒸腾等因素影响下处于动态变化之中,因此研究土壤水分植被承载力的时消周期为“年”。由于土壤含水量必须大于或等于萎蔫系数,植物才能正常生长发育,植物生长和发育只能吸收和利用一定深度土层的土壤水分,不可能利用整个土层的土壤水分,特别是对较厚土层的土壤,如干旱地区。
干热河谷区旱季的土壤水分状况是影响林木生长的更关键的因子,决定着林分的稳定程度。因此,干热河谷地区营造高密度的人工乔木生态林势必会导致林分微环境较差,难以形成结构良好的林分,从而也影响生态功能的发挥。研究表明,干热河谷土壤水分承载力较小的扭黄茅群落自然草坡的土壤水分状况明显优于土壤承载量较大的乔木林和灌木林,但干热河谷地区也存在生长良好的稀树和散生乔木,从而证实稀疏的乔木林能够正常生长。因此,为了维持人工生态林生态系统的水分平衡,在干热河谷植被恢复中应尽量降低造林密度,减小土壤水分承载力,减少水分消耗,确定出合理、适度的造林密度(费世民等,2004,2006)。
2.3立地异质性
制约干热河谷植被恢复和生长的主要限制条件是不同立地条件下土壤层旱季(11月至翌年5月)的有效蓄水量和干旱后期土壤的有效含水量。阴坡和阳坡土壤含水量有很大差异。旱季含水量阴坡总的趋势是下层(40 cm~60 cm)土壤最高。旱季6个月的4种整地方式平均含水量以上层(0~20 cm)最小,为11.98%;中层(20 cm~40 cm)次之,为16.42%;下层为18.03%;而阳坡上、中、下3层含水量分别为9.76%、13.80%、13.71%。阴阳坡旱季不同土层间的土壤水分差异导致了干热河谷阴坡和阳坡间植被分布的差异(蒋俊明等,2007)。同时,不同质地土壤水分条件差异较大。金沙江干热河谷中沙砾石覆盖的阶地、丘陵,以沙质土为主,及裂隙特别发育的基岩山地土壤,水分条件较好,在人工灌溉补肥条件下,植树造林成活率较高;而第三系和第四系的粘土、亚粘土地层和泥质岩类组成的山地发育的土壤,水分条件差,特别是燥红土、变性土,粘粒含量高,只适宜栽种灌草;该区域环境的空间异质性极大,具体体现为微地型和微气候差异、土壤及其土壤母质的变化.这种空间异质性的变异极易误导植被恢复的方向,也就是说,相临地块,由于立地条件的不同,其植被恢复所选用的树种、模式及造林技术理应不同,但因是规模性造林,加之立地生境的脆弱性,在生产上极易忽视这种差异,最终导致造林失败。
因此,干热河谷植被恢复除了“适度”林分结构外,针对立地条件差异性和自然植被景观多样性,还应该保留一定面积的自然植被,使人工林与自然植被交错镶嵌分布,以恢复具有生物多样性特点和林分稳定性人工植被。
3 “适度”造林密度
土壤的自然含水量扣除植物的凋萎系数以后,才是可供植物利用的有效水分。根据土壤供给的有效水分总量和各树种的单株水分消耗量,可估测出土壤对各树种的水分承载能力。根据调查,在旱季0~25 cm土层、25 cm~50 cm土层、50 cm~70 cm土层的土壤平均含水量分别为9.34%、10.24%和11.35%。林地土壤表土层(0~25 cm)水分可供地表草本植物利用;大部分根系的垂直分布在25 cm ~75 cm土壤层;在旱季降雨一般20 mm~50 mm,蒸发量为降雨量的两倍以上,表土水分有一部分用于蒸发,则植物需从深层土壤消耗。因此,植物耗水过程水分复杂,利用Li-1600测定植物蒸腾量,不能反映土壤水分消耗的真实情况,土壤水分蒸发对土壤水分消耗影响很大。为此,采用水浮式盆栽实验方法,以土壤水分变化为中心,把水分消耗作为整体概念来对待,通过对土壤消耗量与水面蒸发量进行拟合,建立拟合实验模型方程,再根据方程,计算出各树种在整个旱季的单株水分消耗量(旱季为11月至翌年5月,计212 d),即可估测出每公顷林地对各树种的最大造林密度。
3.1土壤水分供应能力
根据上述攀枝花市干热河谷土壤水分的研究结论,为了确定土壤对植物的供水范围,对干热河谷现有主要植物根系进行了调查,结果见表1。由此,本研究确定土壤有效供水的土壤深度为70 cm,以此,计算土壤的有效蓄水量作为土壤水分供应能力。

表1 攀枝花干热河谷主要树种根系分布
但是,在干热河谷,荒山荒地的土壤层一般在50 cm,有部分水分需要来自于母质层。根据现有主要林分土壤调查与测定,由表2可见,荒草坡的土壤水分供应能力最小,达到1 143 T·hm-2,五色梅林地、台湾相思林地、加纳比松林地的土壤水分供应能力比较高,达到1 808.8 T·hm-2、1 775.6 T·hm-2、1 725.63 T·hm-2,说明植被对土壤的良好改良作用;同时,现实中的林分在一定程度上已经具有相对稳定性。为此,为了计算“适度”造林的林分密度,考虑到土壤厚度等因素,确定以荒草坡为标准,即干热河谷的基准土壤水分供应能力为1 143 T·hm-2。

表2 攀枝花干热河谷主要林地土壤水分供应能力(有效蓄水)
3.2植物耗水量的计算
在攀枝花干热河谷由于干湿季节分明,从10月至第2年5月为旱季,长达212 d,土壤的贮水量是旱季植物耗水的唯一来源,因此,土壤在雨季贮水量的多少决定了植物能否安全渡过旱季,决定了林种结构、组成和林分密度。对攀枝花干热河谷主要造林树种进行盆栽试验,通过测试某一时段(或某一点)的蒸腾量或蒸散量来计算植物的耗水量明显不合理,因为植物耗水是一个连续的过程,同时蒸腾量与整个旱季的气候状况有关。通过整个干旱期间盆栽试验,以Metcherlich模型拟合结果最理想,可以反映土壤水分的消退过程以及土壤的结构等参数及土壤水分的动力学特征,拟合的数学模型在土壤学上有明确的物理意义。模型方程为:

根据方程拟合了各树种单位水面蒸发量作用于1 cm2林冠蒸腾量(或蒸散量)的累积水面蒸发量模型。对不同种类植物,拟合方程的系数求解,得到结果见表3。

表3 拟合方程的系数
根据拟合方程1,各树种的拟合土壤水分消退过程见图1,来计算不同树种旱季的耗水量。
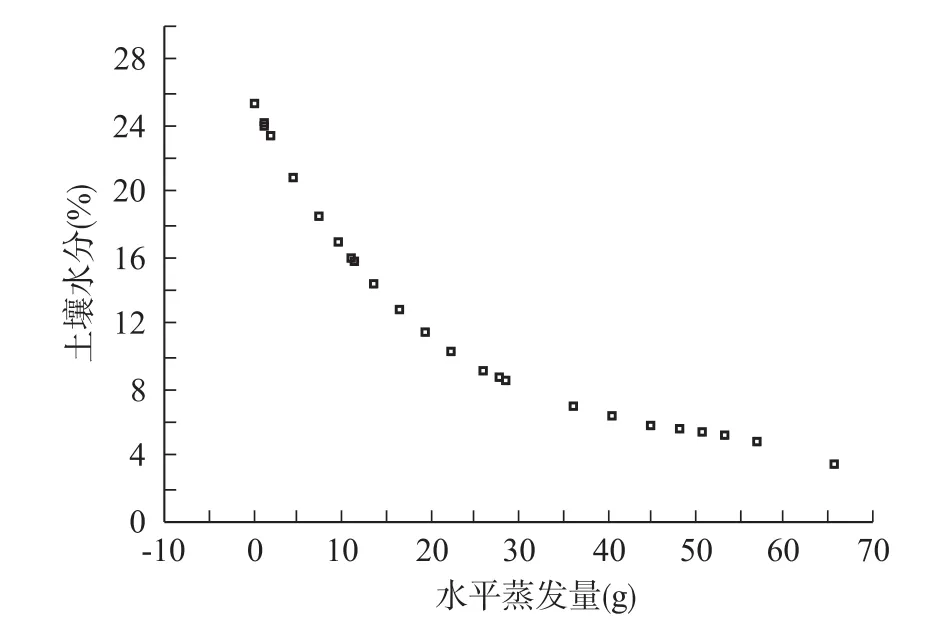
图1 土壤水分消退曲线
研究采用室外水浮式盆栽实验,实际上把单株植物形成的生态空间作为一个计量单位,不同于以往研究,其结果更具有真实性。以土壤水分含量为纵坐标,以水面累积蒸发量为横坐标作土壤水分递减曲线图(以实验拟合的土壤水分消退模型:ET= a + b×e0- k·E),从图1和图2可以看出,土壤水分的递减变化曲线较平滑,从曲线变化趋势看,水分在前期减少较快,到后期土壤水分减少较慢,最后至稳定,但从土壤水分消退曲线看土壤蒸发的3个阶段:起始恒速阶段、中期速降阶段和后期的慢速阶段。

图2 土壤水分消退过程示意图
通过对ET= a + b×e- k.E0求导,得出的拟合方程,按照方程的生态意义:在整个旱季,植物蒸腾耗水是一个动态变化过程,不断消耗土壤水分,逐步累积,因此,以图2中的阴影部分,计算出植物单株耗水量。同时,植物耗水与水面蒸发密切相关,根据前面研究结果,该区水面蒸发量为8 mm·d-1,计算结果见表4。在5个乔木树种中,以直干蓝桉的耗水量最高,达4.6 T,其次是台湾相思,为1.2 T,加勒比松、银合欢、印度楝分别为0.65 T、0.55 T和0.3 T。在灌木中,以五色梅为最高,达0.94T,最低是剑麻,仅为9.5 kg。

表4 攀枝花干热河谷主要造林树种旱季耗水量
3.3“适度”造林的密度
从干热河谷的实际情况看,制约森林形成的主要因素是水分。旱季的土壤水分状况和树木的耐旱性是决定树木能否生存的两个因素。本文所研究的几个树种,经前人研究成果和造林实践证明都表现出一定的抗旱性。因而,这些树种能否形成稳定的人工林,主要取决于土壤的水分状况;从土壤水分供应能力的角度,所计算出的结果是每公顷林地对各树种所能够承载的最大理论株数。
根据目前该区的造林要求,其密度为3 300株· hm-2,按表5的“适度”的密度要求,如印度楝、台湾相思、直干蓝桉、银合欢、加勒比松的林分密度分别为3 771株·hm-2、966株·hm-2、250株·hm-2、2 068株·hm-2和1 769株·hm-2,因此,印度楝的密度较合适,而台湾相思、直干蓝桉、银合欢、加勒比松的林分密度却远远超过了,这样,实际上造成了资金的浪费,台湾相思、银合欢、加勒比松造林分别浪费了约2/3、2/5和1/2的资金,而直干蓝桉造林将节省更多的费用,实际上,形成了“稀树灌草丛”,或者说,在干热河谷,直干蓝桉不适合进行造林。按目前该区造林9 000元·hm-2计,则每hm2至少分别浪费6 000元、3 600元和4 500元。这对该区原本有限的造林经费来说,这种浪费是非常可惊人的。大面积高密度的造林不仅引起土壤水分亏缺,而且造成生态效益与经济效益不统一、长期效益与短期效益的矛盾。
对于灌木来说,小桐子、车桑子和五色梅的造林密度分别为3 161株·hm-2、6 371株·hm-2和1 220株·hm-2,剑麻的造林密度为40 000株· hm-2,由于剑麻抗旱性能比较强,其根系一般在20 cm的土层中,因此,在实际造林中,应以其1/3来造林,即40 000株·hm-2,比较合适。
在实际生产中,还要考虑到其他的立地条件,如土壤肥力、坡位和坡向等,以及林地灌木和草本层等等。因此,如果大于这个造林密度,就会造成旱季土壤供水不足。当然,为了提早郁闭,尽早发挥林分的防护效果,造林的初植密度可以适当大于这个密度。从树木个体发育角度看,当树木地上部分达到一定的生物量,总叶面积的耗水量超过土层的供水能力时,会促使植株生长停止。虽然在以后的雨季里有比较充足的水分供应,也不能恢复其旺盛的生长,因土壤水分胁迫导致林木生长衰退,从而形成“小老头树”,甚至成片死亡。这种现象在我国干旱地区相当普遍。从林分角度来看,同一树种随着森林生物量的增加,树木对水分的消耗加大,有可能因为对水分的竞争加剧而导致林分的自然稀疏,从而无法形成稳定的人工林,林木因为水分营养面积不足而导致部分林木死亡。一方面,从人工林的长期稳定性和改善林地小环境这两方面来考虑,造林初期为了迅速覆盖林地,尽早地形成森林环境,仍需要适当密植。但随着林木的生长,对水分的消耗增大,为保持林分的相对稳定,保证林木的存活与生长并逐步提高林地的生产力,应人为调节其密度,预防林木大范围的死亡而导致人工林的衰退。
因此,“适度”造林的林分即可充分利用水分资源,又不会造成土壤干旱胁迫现象,此时的林分应属于“疏林”(注:指密度低、郁闭度小于0.3的林分)。在干旱区,在选择节水、耐旱树种的前提下,应该在生态、经济和生态经济上可行的考虑,进行适度造林,以维护林分的持续稳定性。

表5 不同树种的适度造林密度估算
4 植被恢复的“适度”配置
20世纪90年代初,中国林科院资源昆虫研究所、四川省林业科学研究院、中科院等科研单位在四川、云南金沙江干热河谷开展植被恢复与造林技术研究,营造大面积人工林,推动了干热河谷的植被恢复工作。“十五”以来,恢复模式逐渐向乔、灌、草结合的多层次立体恢复模式过渡,植被恢复取得较好效果。
目前,根据掌握的文献资料,金沙江干热河谷退化生态系统恢复与重建模式的试验示范研究还停留在一些小的、局部的区域范围内或单一的群落或植被类型,缺乏从区域景观水平或整体系统的区域尺度的综合研究与示范,也缺乏对已有微观的具体模式的宏观配置研究。已有植被恢复技术或成果由点至面的尺度转化过程准确性缺乏,而不能大量被采纳和推广,个别成功案例推广放大后难以适应而失败。自然植被与人工植被景观的结构、动态、功能和相互作用的资料缺乏,它们的适应性难以判断(杨振寅等,2007)。
4.1从植被景观的多样性探讨区域植被群落分布的“适度”格局
据调查研究(费世民等,2004,2006),攀枝花市景观的镶嵌结构发生较大的改变。20世纪50年代初期,稀疏灌草丛和干热河谷禾草灌草丛大约在海拔1 300 m左右一带,木棉和红椿在沿江河两岸还比较多。后来,由于1958年的“大跃进”、人口增长迅猛、建市以来的近700次森林火灾和20世纪60年代~70年代扩建渡口钢铁基地和成昆铁路等,使得整个植被资源遭受巨大破坏和损失,次生植被越来越多,森林覆盖率下降很快。从20世纪70年代至90年代,森林植被的覆盖率下降了4.44%。而同期的陡坡和一些宜林地纷纷被辟为耕地,造成自然植被面积减少,原生植被退缩到高山或陡峻偏远的地方,次生植被扩展,干热河谷线上升;植被镶嵌体的镶嵌关系发生很大变化。而且,就每种镶嵌体而言,其内部结构在这几十年间也发生较为明显的变化,森林结构日趋简单化,乔木减少,灌木、草本植物增多。最破碎的植被景观出现在人类活动最剧烈的两江(金沙江和雅砻江)、两河(安宁河和大河)干热河谷地区。干热河谷区占优势的植被为草丛、灌草丛、稀树草丛、稀树灌草丛。
据杨振寅(2007)对元谋干热河谷植被景观动态与植被恢复研究表明,元谋干热河谷车桑子灌木林和稀树灌草丛占明显优势,为元谋干热河谷景观的基质,其面积和斑块数达540.60 km2和339.51个,分别占干热河谷区景观总面积和总斑块数的40.52%和25.45%。植被斑块为细粒结构,以中小斑块为主,小于50 hm2的斑块占植被斑块数的91.58%。其中,稀树灌草丛和车桑子灌木林的小斑块数量最多,占植被景观小斑块总数的75.15%。植被景观面积主要集中在中、小斑块,稀树灌草丛和车桑子灌木林中、小斑块的面积占植被总面积的41.94%。
可见,在干热河谷区,植被恢复除了按自然植被演替规律营建“适度”林分结构外,还应该从区域尺度上,依据自然植被景观格局,不宜实施大规模的人工造林,因地制宜地营建块状人工林,形成人工植被与自然植被交错镶嵌分布的“适度”格局,以恢复具有景观多样性特点的区域性植被生态系统。
4.2从立地条件的空间异质性探讨区域植被群落的“适度”配置
在干热河谷区,同一区域的降雨差异不大,但由于地形和下垫面的差异,导致植物水分供应的较大差异,从而决定了干热河谷区植被群落的空间格局。影响干热河谷区植被生长与分布的主要因素有海拔高度、坡向、母岩类型、坡度、土层厚度等(费世民,2004)。
目前,攀枝花干热河谷整体上属于稀树草原类型,但在地势较低、水源条件和土壤条件较好的地段仍有很多林木生长,在非石灰岩山地为锥连栎(Quercus fianchetii)林;石灰岩山地为铁橡栎(Quercus cocciferoides)林;目前格里坪一带石灰岩山地分布有苏铁(Cycas revoluta)林;沟菁、阴坡等土壤水分条件较好的地段为黄栎林。
据杨忠(2003)研究,在金沙江干热河谷区,泥岩坡地土体黏重板结,入渗能力弱,天然降水入渗少,对土壤水分的有效补充较少。在干旱季节土体极其干旱,林木生长停止,甚至受到干旱的生理伤害枯死,林分生产力低,极难恢复森林植被。片岩坡地砾石层坡地砂砾层坡地等石质山地土体裂隙发育,入渗能力强,天然降水入渗多,对土体水分的有效补充较多,在干旱季节岩土深层有少量有效储水供林木吸收利用,维持其正常生理活动的水分需要,林木生长较泥岩坡地上的林木生长快,林分生产力高。
据研究(费世民,2004;蒋俊明等,2204,2005,2007),攀枝花干热河谷的四种土壤中以山地粗骨质红壤全年含水率最低,含水率在5%以下的时间长达5月。因此,造林十分困难,深垦整地,作好蓄水保墒工作是提高造林成效的关键措施。山地碳酸盐红褐土是本区土壤含水率最高的一种土壤,全年变动幅度不大,且较稳定,一年中除旱季3月~5月表土层(9 cm~10 cm)含水率在5%~10%以外,其余时间各层土壤含水率都>20%。这种土壤造林效果较好,阴坡撒播云南松可以成功。山地红色石灰土,含水率虽然也高,但是由于土壤物理性粘粒含量较大,水分多以束缚水的形式保存于土壤之中,可供植物利用部分较少,因此,植物生长差,造林仍很困难。山地黄红壤,含水率仅次于山地碳酸盐红褐土,干湿季明显,一年之中只有两个月时间(4月、5月)表土层(0~20 cm)比较干燥,含水率在9%左右,其余时间都处于潮润状态,插花性干旱对土壤含水率影响不大,造林比较容易成功。
从不同地形条件土壤含水率的观测来看,由于蒸发量大于降雨量,地形因素对土壤水分再分配的影响特别明显。山地粗骨质红壤,虽然含水率很低,若分布在海拔1 500 m以上,土壤仍较潮湿;山地碳酸盐红褐土含水率虽然较高,若处于阳坡,土壤仍很干燥。上述研究说明,在研究金沙江河谷荒山造林的时候,不仅要注意到不同质的土壤含水率的差异,而且还必须考虑到,由于地形条件引起的土壤水分变化,只有这样,才能做到因地制宜,区别对待。
可见,干热河谷植被恢复除了“适度”林分结构外,还应针对立地条件的差异,考虑不同立地类型生境的土壤水分条件,依靠优势生活型植物种类,宜乔则乔、宜灌则灌、宜草则草,适地适树,在不同立地条件的地块上,营建“适度”密度的不同类型人工植被,进行灌草丛、稀树草丛、稀树灌草丛等植被的“适度”配置,构建不同植被与生境条件相协调、稳定的群落生态关系,形成不同植被景观交错镶嵌分布的干热河谷区自然生态系统稳定格局。
因此,在干热河谷植被恢复过程中,应充分考虑区域空间尺度的不同植被类型“适度”配置,是确立科学的植被恢复目标、提高植被恢复与造林成效的关键。
5 结果与讨论
(1)过去北方干旱地区的樟子松、干热河谷的云南松、思茅松等造林的教训,由于造林密度偏大,将引起土壤水分亏缺,影响固沙林生态稳定性,甚至林木死亡。因此,在干热河谷地区,大气水热条件的不平衡,生境对植被的支持力较低,植被恢复要从自然植被演替规律、土壤水分承载力等方面,参照地带性和隐域性原生植被的特征,根据土壤水分承载力与植物水分需求的水分平衡,从林分结构上确定林分生态稳定的“适度”造林密度,适地适树,宜乔则乔、宜灌则灌、宜草则草,进行不同生活型植物类型的合理配置,构建干热河谷的稀树灌草丛植被,避免进入人工造林的林分密度过大、盲目追求生产力的误区。
(2)在干热河谷植被恢复过程中,从区域空间尺度的植被景观多样性、立地条件的空间异质性方面,进行在区域尺度上不同植被类型的“适度”配置,构建不同植被类型与生境条件相协调、稳定的“板块镶嵌”群落配置结构,形成干热河谷区不同植被景观交错镶嵌分布的自然生态系统稳定格局,避免进入不切实际的大规模成片造林而成效难以巩固的误区。
(3)上述分析表明,以干热河谷的自然植被演替、土壤水分承载力、立地异质性为依据,实施“适度”造林,在微观上,构建“适度”密度的林分结构,建立适地、适树、适度结构的植被恢复技术;在宏观上,构建区域植被群落“适度”配置模式,建立区域尺度的不同植被景观“板块镶嵌”空间格局;从而提高与巩固干热河谷植被恢复与造林的成效。因此,干热河谷植被恢复的目标应以“适度”造林为恢复策略,进行人工促进恢复,与封山、禁伐、禁牧等自然恢复相结合,建立群落结构稳定、植被景观配置多样的区域生态系统。
(4)以往研究多以植物蒸腾来计算,而且以某段时间的测定结果为依据,计算出来的结果往往偏小,与实际情况差异大。本研究参照蒸渗仪法,采取室外自然条件下的水浮式盆栽试验,提出的耗水量是一个综合概念,是地表的蒸发和植物蒸腾的总和,即蒸发散,就是植物在地表所占的空间内总的水分消耗;通过整个干旱期间的连续测定,由于干旱期间基本没有降雨,土壤得不到水分补充,土壤水分越来越少,不可能象正常水分供应条件下的一次性消耗,土壤水分消退呈幂指数函数递减,所以,比较切合实际情况。在计算耗水量时,以拟合方程为基础,进行幂指数求解计算出来,实际上是水分消耗的不断积累,对于以土壤水分平衡为中心的水分消耗来说,更加符合实际情况。目前,植物耗水量的测定主要有植物蒸腾仪、蒸渗仪、树干茎流仪等方法,但难以代表植被的耗水量。植被耗水量受不同植物、地表覆盖、土壤等诸多因子的影响,准确测定植被的实际耗水量很难。本研究采取的室外盆栽试验,而且在实验中主要以苗木或幼树为对象,取得的试验数据偏小,以此计算出的林分密度偏大,在实际应用中,应根据具体情况降低林分密度。随着林木的生长,林木对水分的消耗增大,而地表覆盖程度也增大,蒸发散是增大,还是减小,还需要进一步深入研究。
(5)本研究采用同一土壤、同一环境条件下进行测定不同植物的耗水量,是以土壤水分消耗来说明的,主要满足于不同植物的比较,但不同土壤、立地、微环境等不同,存在诸多差异,还需要进一步深入系统研究。
(6)本研究尽管考虑了植被恢复的空间尺度,但还应考虑时间尺度的植被恢复问题,可根据植被演替进程,调整改造人工植被建设的可适度,进行“渐进式”的植被恢复,加快干热河谷植被恢复的进程。(7)本研究立足于自然条件下植被恢复的生物措施,但干热河谷植被恢复是一项生态治理的系统工程,涉及环境、经济、社会诸因素,涉及多部门配合、多学科研究,在制定植被恢复方案时,必须把生物措施与水土流失治理、荒漠化治理、国土整治等工程措施相结合,才能有效推进干热河谷生态建设,加快植被恢复进程。因此,应多方面考虑植被恢复与生态建设方案的现实性和操作性,根据不同区域的特点,制定可行的技术路线,开展对恢复结果的预测和评价,在完整的科学体系下进行有序、合理的干热河谷区植被恢复与生态建设。
参考文献:
[1]费世民,王鹏,陈秀明,等.论干热河谷植被恢复过程中的适度造林技术[J].四川林业科技,2003,24(3):10~16.
[2]费世民.川西南山地生态脆弱区森林植被恢复机理研究[D].北京:中国林业科学研究院,2004.
[3]张旭东,费世民,周金星.中国西部山地生态脆弱区植被恢复研究[M].北京:科学出版社,2006.
[4]周麟.云南省元谋干热河谷的第四纪植被演化[J].山地研究,1996,14(4):239~243.
[5]金振洲,欧晓昆.元江、怒江、金沙江、澜沧江千热河谷植被[M].昆明:云南大学出版社,2000.
[6]李昆,曾觉民.金沙江干热河谷主要造林树种蒸腾作用研究[J].林业科学研究,1999,12(3):244~250.
[7]杨忠,张信宝,等.金沙江干热河谷植被恢复技术[J].山地学报,1999,17(2):152~156.
[8]王克勤,沈有信,等.金沙江干热河谷人工植被土壤水环境[J].应用生态学报,2004,15(5):809~813.
[9]杨振寅,苏建荣,等.干热河谷植被恢复研究进展与展望[J].林业科学研究2007,20(4):563~568.
[10]张信宝.陈玉德.云南元谋干热河谷区不同岩土类型荒山植被恢复研究[J].应用与环境生物学报,1997,3(1):13~18.
[11]蒋俊明,费世民,等.干热河谷几种主要造林树种抗旱能力比较[J].造纸学报,2004,19(增刊):345~348.
[12]蒋俊明,费世民,等.植物抗旱性指标的初步研究[J].山地学报,2005,23(增刊):1~6.
[13]蒋俊明,费世民,谭中月..攀枝花干热河谷阴坡和阳坡土壤水分的动态比较[J].四川林业科技,2005,26(5):20~25.
[14]蒋俊明,费世民,等.金沙江干热河谷植被恢复探讨[J].西南林学院学报,2007,27(6):11~15.
[15]费世民.王鹏.陈秀明,等.攀桂花市生物多样性与生物资源可持续性分析[J].四川林业科技,2002,23(3):14~22..
[16]曲仲湘,吴玉树.王焕校,等.植物生志学[M].北京:高等教育出版社,1983,152~170.
[17]MANION P D,D H GRIFHN.Large land scale analysis of tree death in the Adirondack Park[J].New York,For sci.2001,47 (4):542~549.
[18]费世民,彭镇华,等.我国封山育林研究进展[J].世界林业研究,2004,17(5):29~32.
[19]费世民,杨玉坡.论四川林业在“长江上游生态屏障”建设中的地位与作用[J].四川林业科技,2002,23(1):27~34.
[20]陈荷生.腾格里沙漠东南缘沙坡头地区防护林建设与生物气候关系的研究[J].干旱区资源与环境,1993,7(1):1~8.
[21]郭普.干旱地区灌木林资源保护性培育与利用[J].北京林业科技,1986,(1):8~11.
[22]徐化成.发展人工林与生态学原理[J].河北林学院学报,1991,6(3):128~225.
[23]杨文斌,杨明.樟子松,等.人工林土壤水分收支状况及其合理密度的初步研究[J].内蒙古林业科技,1992(4):47~54.
[24]杨维西.中国北方地区人工植被土壤干化的初步探讨[J].林业科学,1996,32(1):78~85.
[25]高文学,王志和,等.金沙江干热河谷稀树灌草丛植被恢复方式研究[J].林业调查规划,2005,30(3):87~91.
A discussion on“Moderation”Afforestation in the Process of Vegetation Restoration in the Dry-hot Valley
FEI Shi-min1JIANG Jun-ming1ZHANG Xu-dong2ZHOU Jing-xing3
(1.Sichuan Academy of Forestry,Chengdu 610081;2.Research Institute of Forestry,CAF,Beijing 100091;3.Beijing Forestry University,Beijing 100083)
Abstract:In this paper,on the basis of the“moderate”afforestation technology proposed in the process of vegetation restoration in the dry hot valley,and according to the natural vegetation succession,soil water carrying capacity and site heterogeneity,discussion was made on the“moderation”density of afforestation in stand structure and the“moderation”community allocation of vegetation restoration on the regional landscape scale from the perspective of systematic and quantitative research.The“moderation”afforestation techniques and theories were further illustrated in order to provide reference for research and practice of vegetation restoration in the dry hot valley.
Key words:Dry-hot Valley,Vegetation restoration,“Moderation”afforestation,“Moderation”density of afforestation,“Moderation”community allocation
作者简介:费世民(1966-),男,博士,研究员,主要从事森林生态,森林培育研究工作。
基金项目:国家林业局“四川森林生态与资源环境重点实验室”资助项目。
收稿日期:2015-12-10
doi:10.16779/ j.cnki.1003-5508.2016.01.003
中图分类号:S728
文献标识码:A
文章编号:1003-5508(2016)01-0013-10
猜你喜欢
小哥白尼(神奇星球)(2021年3期)2021-07-22
小哥白尼(神奇星球)(2021年4期)2021-07-22
Zoological Research(2019年3期)2019-06-28
乡村地理(2018年4期)2018-03-23
高原山地气象研究(2016年2期)2016-11-10
成才之路(2016年26期)2016-10-08
塔里木大学学报(2014年3期)2014-03-11
自然资源遥感(2014年2期)2014-02-27
自然资源遥感(2014年2期)2014-02-27
