
贵州省岩溶地区野生白三叶形态多样性
2016-04-11 16:12吴永洁张俞池永宽王元素
江苏农业科学 2016年2期
吴永洁++张俞+++池永宽++王元素+++李莉


摘要:在贵州省典型岩溶地区采集22份野生白三叶(Trifolium repens)种子于温室内栽培,以观测其形态特征多样性,探索白三叶的形态学差异与地理环境的关系。结果表明:贵州白三叶形态多样性丰富,形态特征变异大小为:匍匐茎长度>中叶面积>地上生物量>生长点个数>有无“V”形斑纹>株高>中叶宽>中叶长>生长习性。不同海拔地区白三叶的形态性状均具有广泛的变异,不同性状的变异程度也不同。聚类分析将22份材料分为3类。为进一步开展白三叶遗传多样性研究,选育出适应喀斯特特殊生态环境的优良白三叶品种奠定基础。
关键词:贵州;白三叶;形态学;多样性;岩溶地区
中图分类号: S326;S181文献标志码: A文章编号:1002-1302(2016)02-0291-06
收稿日期:2015-08-21
基金项目:国家自然科学基金(编号:31260115);贵州省科技厅社发项目(编号:黔科合SY字(2012)3170);贵州省重大科技攻关专项(编号:黔科合重大专项字[2011]6009号);贵州省自然科学基金(编号:黔科合[J]2009-2091)。
作者简介:吴永洁(1991—),女,广西贵港人,硕士研究生,研究方向为草地生态学。E-mail:wuyjiekaobo@163.com。
通信作者:李莉,博士,副教授,研究方向为分子生物学。E-mail:liligznu@163.com。在家畜放牧采食等干扰下,草地植物会发生相应的进化。短期干扰下植物发生生理反应以应对组织被采食后碳水化合物供给和光的不足;长期则发生形态学的变化,进化“回避”采食机制以减少被采食的机会而持久存在[1]。“回避”是指减少被采食的可能性和强度,主要表现为结构特性、物理特性等变化,形态特征多样性高[2]。贵州省于1905年引进白三叶(Trifolium repens)种植,后逸生为野生种[3],广泛分布于全省66个县市,占全省总县市的79.5%[4],白三叶已经成为贵州最主要的豆科牧草。经过100多年的进化,其形态特征发生了进化响应。开展贵州野生白三叶株高、匍匐茎长度、“V”形斑纹等表型特征的研究,探讨其形态多样性的进化机制,对合理开发利用和保护白三叶种质资源,进一步选育出适应喀斯特特殊生态环境的优良品种有积极的理论与现实意义。
1材料与方法
1.1贵州地理与气候
试验在贵州省花溪区贵州牧草推广站温室进行,种子收集于贵州省各地。贵州属于亚热带湿润季风气候,四季分明,平均气温在15 ℃左右,由于纬度较低,许多地区的气候冬无严寒,夏无酷暑,最冷的1月份平均温度在3~8 ℃之间,而最热的7月份平均温度也在18~26 ℃之间。贵州省由于受季风影响,冷暖气流交汇频繁,年降水量在1 100~1 300 mm之间,但降水季节分配不均,80%的雨水都集中在5—10月份。贵州省的地势起伏较大,地形也较复杂,导致气温的垂直变化十分明显,气候类型多样。
1.2材料收集
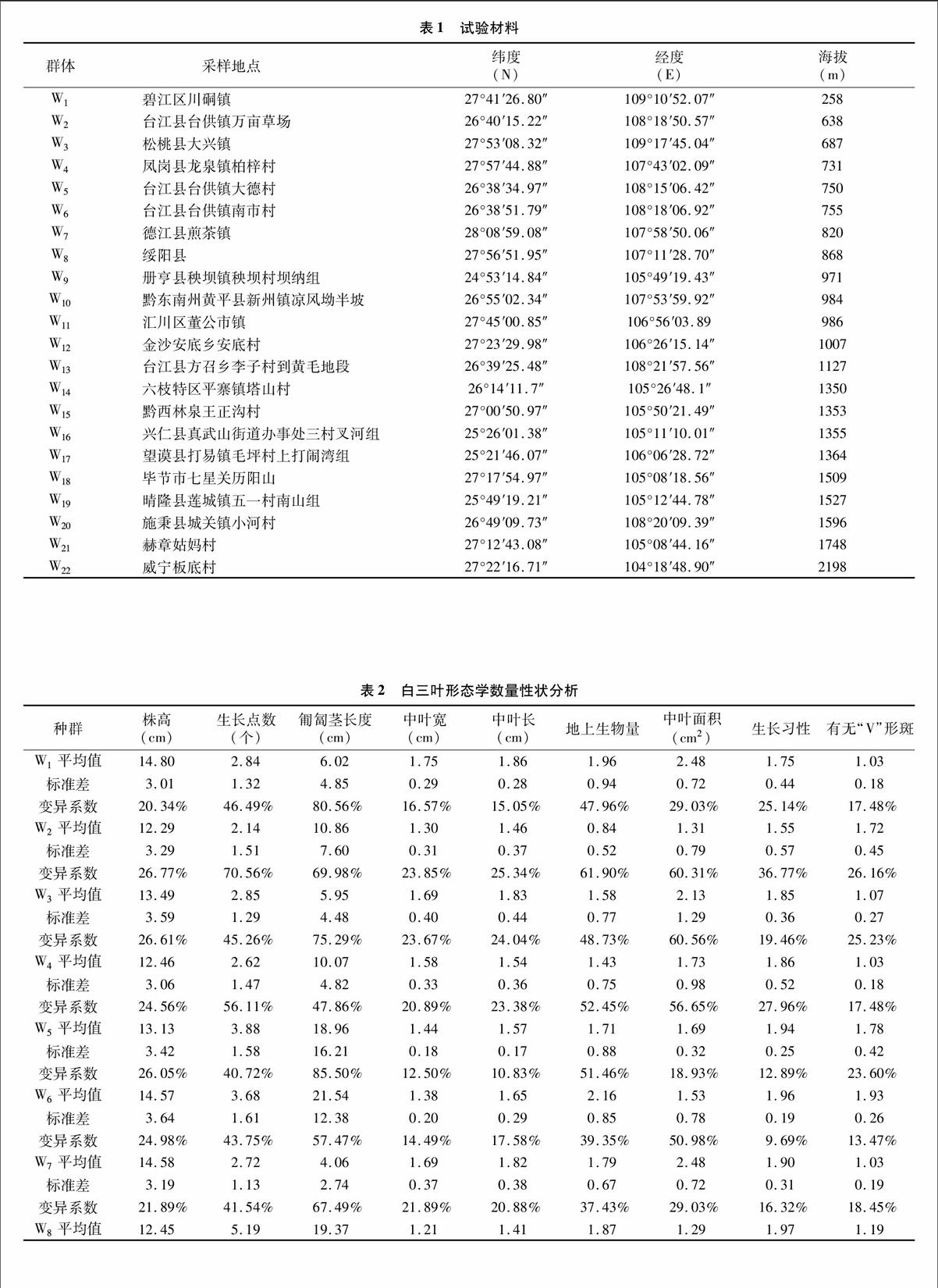
2013年5—8月在贵州省范围内采集野生白三叶种子,有效材料22份,详见表1。
1.3栽培与观测
1.3.1温室栽培于2014年3月20日在贵州省牧草推广站温室大棚播种。播种前种子进行低温处理破除硬实。培养基质按m石英砂 ∶m细土 ∶m腐殖质=3 ∶4 ∶3混合而成,高压灭菌后冷却装盘。播入32格种苗培养盘,育苗盘大小54 cm×28 cm,孔穴大小60 mm×60 mm。为保证出苗率,每格播10粒种子,覆土1 cm,浇透水。等长出小苗后留选出长势最好的1株苗。
1.3.2施肥管理苗期施尿素1次,使用量折合 0.012 kg/m2。以后隔3个月施复合肥1次,每次施用量 0.03 kg/m2。及时除杂,适时浇水。
1.3.3测量指标2014年9月1日观测“V”形斑纹、叶面积、株高、中叶宽、中叶长、匍匐茎长度、生长点个数,2014年10月3日测定地上生物量等。
为便于分析,将非量化性状特征进行标准化赋值。有无“V”形斑:有斑=1,无斑=2;海拔:1=500 m以下,2=501~1 000 m,3=1 001~1 500 m,4=1 501~2 000 m,5=2 001 m以上;生长习性:1=直立生长,2=匍匐分枝生长,3=匍匐不分枝生长。
1.4数据分析
用Excel和SPSS 16.0对试验数据进行统计分析,分析种内、种间各个性状的差异程度。用Person相关系数分析各个性状之间的相关性,并对各个指标标准化后用算术平均法(UPGMA)进行聚类分析。
2结果与分析
2.1白三叶形态学分析
2.1.1白三叶形态学性状差异分析贵州省不同种质资源表1试验材料
间的白三叶在形态上存在较大差异,表现出显著的形态多样性(表2和表3)。白三叶种内差异显著,9个形态特征之间的变异系数都在20%以上。匍匐茎长度变异系数最大,达86.60%,变异范围为1.00~87.00 cm。其余形态特征变异大小为:中叶面积>地上生物量>生长点数>有无“V”形斑纹>株高>中叶宽>中叶长>生长习性,生长习性最小,为 20.23%。
2.1.2不同地理位置白三叶形态学差异分析7个数量形态性状的分析结果(表4)表明,不同海拔地区白三叶的形态性状均具有广泛的变异,不同性状的变异程度也不同。总体上,株高、地上生物量及中叶面积随着海拔升高而呈现下降趋势,生长点数及匍匐茎长度随海拔升高呈上升趋势。
在海拔低于500 m的区域,白三叶总变异系数高达 252.31%,其中匍匐茎长度呈现出显著的差异,变异系数为80.56%,最长为20.00 cm,最短只有2.00 cm。其次是地上生物量、生长点数、中叶面积、株高、中叶宽、中叶长,其变异系数分别为47.96%、46.48%、25.35%、20.34%、1656%、15.05%。
在海拔为501~1 000 m的区域,白三叶匍匐茎长度的变异程度最大,达90.19%,变异范围为1.00~87.00 cm,其平均长度为12.84 cm;其中最长的是W6(台江县台供镇南市村,21.54 cm),最短的是W7(德江县煎茶镇,4.06 cm)。其余变异大小为生长点数>地上生物量>中叶面积>株高>中叶宽>中叶长,它们的变异系数分别为51.23%、50.00%、42.86%、25.21%、22.00%、20.99%。
在海拔为1 001~1 500 m的区域,白三叶总变异系数为298.21%,其中匍匐茎长度的变异系数最大,为78.21%,变化范围为2.00~62.00 cm,其总平均长度为15.60 cm,其中最长的是W16(兴仁县真武山街道办事处三村叉河组,19.33 cm),最短的是W17(望谟县打易镇毛坪村上打闹湾组,5.50 cm)。其余变异大小为地上生物量>生长点数>中叶面积>株高>中叶长>中叶宽,它们的变异系数分别为 70.34%、49.56%、35.63%、24.96%、20.39%、19.12%。
在海拔为1 501~2 000 m的区域,总变异系数高达 329.38%,其中白三叶匍匐茎长度变异系数最大,为 84.56%,变异范围为1.50~49.00 cm,其平均长度为13.54 cm,其中最长的是W21(赫章姑妈村,平均19.30 cm),最短的是W20(施秉县城关镇小河村,平均2.94 cm)。其余变异大小为地上生物量>生长点数>中叶面积>株高>中叶长>中叶宽,它们的变异系数分别为58.87%、53.38%、49.13%、33.02%、26.24%、24.18%。
在海拔高于2 000 m的区域,匍匐茎长度和地上生物量都呈极显著差异,其变异系数分别为79.99%、61.78%,变异范围分别为2~45 cm、0.07~4.49 g。其余变异大小为中叶面积>生长点数>株高>中叶长>中叶宽,其变异系数分别为47.29%、44.58%、32.76%、25.76%和25.00%。
2.2形态学性状相关性分析
2.2.1形态学特性相关性分析供试材料主要形态学性状的相关性分析结果见表5,各性状间采用Person相关系数分析。结果表明,株高与中叶宽、中叶长、中叶面积及地上生物量呈极显著正相关,与生长点个数、匍匐茎长度、生长习性呈极显著负相关;生长点个数与匍匐茎长度、地上生物量、生长习性呈极显著正相关,与中叶长、中叶宽、中叶面积呈极显著负相关;匍匐茎长度与中叶长、中叶宽、中叶面积呈极显著负相关,与地上生物量、有无“V”形斑纹呈极显著正相关;中叶长、中叶宽、地上生物量两两呈极显著正相关,同时,中叶长、中叶宽还与有无“V”形斑纹呈极显著负相关,地上生物量与生长习性呈极显著正相关;中叶面积与有无“V”形斑纹呈极显著负相关。分析说明白三叶植株越高,中叶长宽、中叶面积都较大,地上生物量也越高,而生长点个数越少,偏向于直立生长。生长点个数越多,则匍匐茎越长,中叶的长宽及面积都越小,偏向于匍匐不分枝生长。匍匐茎越长,则地上生物量越大,偏向于无斑的匍匐不分枝生长。
2.2.2形态学特性与地理分布的相关性分析海拔与株高、中叶宽、中叶长及中叶面积呈极显著负相关,与匍匐茎长度呈显著正相关,地上生物量与海拔高度呈显著负相关。表明随着海拔提高,白三叶长得越矮小,中叶长、宽、面积也越小,地上生物量也随海拔高度提升而减少。总体上,在高海拔地区,植株较矮小,匍匐茎较长,在低海拔地区,植株较高,匍匐茎较短。而生长习性、有无“V”形斑纹与海拔高度无显著相关性。
2.3聚类分析
以株高、中叶长、中叶宽、中叶面积、生长点数、匍匐茎长度、地上生物量、海拔、生长习性等作为表观形态性状指标,标准化后对22份白三叶材料进行聚类分析,聚类方法选用类平均法,距离测度采用欧氏距离。聚类分析如图1所示。
从种质资源水平上说,先聚到一起的种质材料之间的亲缘关系较近,即距离测度越近,亲缘关系越近。聚类分析(图1)表明,大部分来源地域距离邻近的种质材料先聚到一块,说明它们的亲缘关系较近。在欧氏距离为15.0处,将22份材料聚为3类。第1类:W1和W20聚为一类,植株最高(其平均值分别为14.80、15.58 cm),匍匐茎较短,中叶面积也最大,大多有斑;第2类:生长点数较多和匍匐茎也较长,地上生物量也较大的材料W15、W6、W13、W16、W22、W12、W10、W5、W8、W21、W11、W3、W18聚为一类。第3类:W19、W17、W4、W9、W2、W7及W14聚为一类,大多有斑纹,生长点数和匍匐茎长度介于第1类和第2类之间,地上生物量较小,少数直立生长。这说明,由于遗传具有稳定性,在同一环境中生长的不同种质资源的白三叶在形态上也存在较大的差异。
3结论与讨论
3.1白三叶的形态多样性与进化
贵州野生白三叶形态差异较大,多样性丰富。王玉祥等研究了来自新疆13份白三叶的表型性状,结果发现,其形态特征的变异系数在8.5%~24.75%之间[5];何俊对国内外62份白三叶草种质资源形态多样性进行了分析鉴定,其15个重要形态学性状指标变异系数范围为14.32%~76.51%[4];而本研究中白三叶9个形态特征的变异系数范围比以上地区更广泛,为20.32%~86.60%。喀斯特山区贵州毕节的12份野生燕麦的7个农艺性状遗传多样性的变异系数范围为 9%~75%[6],较新疆燕麦的1.7%~34.89%[7]和国内外燕麦的11.29%~52.65%[8]变异范围都大。从生物进化论看,任何物种都在随生境条件的变化而变化;植物的形态特征受本身的遗传组成和所生存环境的影响[5],由于贵州省境内海拔变化范围广,石漠化相当严重,并在长期的自然选择和人工选择作用下,白三叶为适应当地生境而形成了其特有的形态特征,因而具有丰富的形态多样性。
3.2白三叶形态多样性对环境的响应
白三叶的形态多样性与海拔等因素相关[9],李州等研究表明,白三叶形态变异与地理起源存在较密切的相关性[10]。本研究的Person相关系数分析发现,白三叶的形态特征与海拔高度呈显著相关,小叶白三叶主要分布于高海拔地区,大叶白三叶则分布于低海拔地区,这与Caradus等的研究结果[11-12]一致。植物的形态特征与其地理分布及其生境有关,孙雪梅等发现云南野生茶树形态多样性与其地理分布有关[13],肖苏等证明了川渝地区的野生鹅观草的形态特征与其地理分布和生境存在一定的关系[14]。在不同的生境条件下,植物形态学特征发生变化,有利于适应不同环境。白三叶形态变异大,形态可塑性大,生态适应性越强。利用植物的表型特征对其进行相关研究,不仅可以初步了解植物遗传变异水平,还可以研究其生物适应和进化的方式、机制及其影响因子等[15]。贵州省白三叶形态多样可能与贵州省特殊的喀斯特生态环境有关,还有待进一步研究,后续将进一步开展白三叶基因型多样性的研究。
3.3白三叶形态特征与育种
贵州省白三叶形态特征丰富,是白三叶育种的重要基础。在育种过程中可选择差异较大的品种作为亲本或重点研究对象,有望培育出植株高、匍匐茎长、生物量大的新品种。根据本研究结果,可利用海拔较低的区域,如碧江区川硐镇、台江
县台供镇万亩草场及松桃县大兴镇等叶大且高的白三叶育种成为刈割型白三叶;在海拔较高的区域,如威宁板底村和赫章姑妈村等匍匐茎较长的白三叶适合培育成耐放牧的品种。但白三叶因其耐盐碱、干旱能力差等原因致使其应用受到了广泛限制。目前,我国的白三叶育种工作还比较滞后,相关的研究报道还不多。安晓珂初步评价了新疆野生白三叶与三叶草属其他种的遗传多样性及遗传关系[16],何俊研究了国内外62份白三叶种质资源的遗传多样性[4],王玉祥等研究了新疆13份野生白三叶表型性状变异情况[5],张贤等对白三叶RAPD分析条件进行了优化[17]。因此,下一步工作可全面调查和收集国内外野生白三叶资源,应用分子标记和转基因等技术手段,增加其多样性,扩大其基因库,对合理开发和利用白三叶资源有重要的指导意义。
参考文献:
[1]Lemaire G,Chapman D. Tissue flows in grazed plant communities[M]//Hodgson J,Illius A W. The ecology and management of grazing systems. CABI Publishing,1996:3-36.
[2]Briske D D. Strategies of plant survival in grazed systems:A functional interpretation[N]//Hodgson J,Illius A W. The ecology and management of grazing systems. CAB International,1996.
[3]王元素,李莉,孔玲. 百年足球运动场白三叶草坪群落特征研究[J]. 草原与草坪,2012,32(2):55-57,60.
[4]何俊. 62份白三叶草种质资源遗传多样性初步研究与评价[D]. 贵阳:贵州大学,2008.
[5]王玉祥,张博. 新疆野生白三叶表型性状变异研究[J]. 草地学报,2012,20(6):1163-1168.
[6]蹇黎,秦小军,余丹凤,等. 喀斯特山区野生燕麦农艺性状的遗传多样性分析[J]. 河南农业科学,2013,42(6):27-31.
[7]马艳明,刘志勇,白玉亭,等. 新疆燕麦地方品种资源多样性分析[J]. 新疆农业科学,2006,43(6):510-513.
[8]张向前,刘景辉,齐冰洁,等. 燕麦种质资源主要农艺性状的遗传多样性分析[J]. 植物遗传资源学报,2010,11(2):168-174.
[9]Widdup K H,Caradus J R,Green J,et al. White clover ecotype germplasm from the USA for development of New Zealand and overseas cultivars grassland[J]. Research and Practice Series,1996,6:149-154.
[10]李州,彭燕,张婧源,马啸. 白三叶种质资源形态变异与地理起源的关系[J]. 草业科学,2012,29(11):1706-1714.
[11]Caradus J R,MacKay A C,Woodfield D R,et al. Classification of a world collection of white clover cultivars[J]. Euphytica,1989,42(1):183-196.
[12]Ellis W,Young N R. The characteristics of European,Mediterranean and other populations of white clover (Trifolium repens L.) [J]. Euphytica,1967,16(3):330-30.
[13]孙雪梅,黄玫,刘本英,等. 云南野生茶树的地理分布及形态多样性[J]. 中国农学通报,2012,28(25):277-288.
[14]肖苏,张新全,马啸,等. 野生鹅观草种质的醇溶蛋白遗传多样性分析[J]. 草业学报,2008,17(5):138-144.
[15]赵云青,陈菁瑛,慕小倩. 曼陀罗不同居群形态多样性比较研究[J]. 热带作物学报,2013,34(9):1836-1843.
[16]安晓珂. 三叶草属三种植物遗传多样性的RAPD分析[D]. 北京:中国农业科学院,2008.
[17]张贤,张英俊,吴维群. 白三叶RAPD分析条件优化[J]. 草地学报,2006,14(3):219-222.贺刚,方春林,陈文静,等. 鄱阳湖水道四大家鱼群落特征及幼鱼入湖格局[J]. 江苏农业科学,2016,44(2):297-299.
doi:10.15889/j.issn.1002-1302.2016.02.087
猜你喜欢
现代园艺(2023年3期)2023-02-01
农家致富顾问·上半月(2022年9期)2022-05-30
现代园艺(2017年22期)2018-01-19
大陆桥视野·下(2016年11期)2017-02-28
科技资讯(2016年19期)2016-11-15
人间(2016年27期)2016-11-11
草业科学(2013年11期)2013-04-25
