
Taxonomic revision of lizards from the Paleocene deposits of the Qianshan Basin, Anhui, China
2016-03-29 04:24:31DONGLiPingSusanEVANSWANGYuan
古脊椎动物学报(中英文) 2016年3期
DONG Li-PingSusan E. EVANSWANG Yuan
(1Key Laboratory of Vertebrate Evolution and Human Origins of Chinese Academy of Sciences,Institute of Vertebrate Paleontology and Paleoanthropology,Chinese Academy of SciencesBeijing 100044, China dongliping@ivpp.ac.cn)
(2Department of Cell and Developmental Biology,University College LondonLondon WC1E 6BT, UK)
Taxonomic revision of lizards from the Paleocene deposits of the Qianshan Basin, Anhui, China
DONG Li-Ping1Susan E. EVANS2WANG Yuan1
(1Key Laboratory of Vertebrate Evolution and Human Origins of Chinese Academy of Sciences,Institute of Vertebrate Paleontology and Paleoanthropology,Chinese Academy of SciencesBeijing 100044, China dongliping@ivpp.ac.cn)
(2Department of Cell and Developmental Biology,University College LondonLondon WC1E 6BT, UK)
Although the Late Cretaceous lizard fauna of China and Mongolia is relatively well-known, information on Paleocene lizards from the same region is currently limited. Several species of lizards have been reported from the Paleocene Wanghudun and Doumu formations of Qianshan Basin on the basis of fragmentary specimens, namelyAgama sinensisHou, 1974,Anhuisaurus huainanensisHou, 1974,Anqingosaurus brevicephalusHou, 1976,Changjiangosaurus huananensisHou, 1976,Qianshanosaurus huangpuensisHou, 1974, andTinosaurus doumuensisHou, 1974. In this paper, we review all the reported material of these taxa with the aid of new technology, including CT scanning, and according to current views of squamate relationships and classi fi cation. Revised descriptions and classi fi cations are given for each taxon, leading to changes in our understanding of faunal composition. This, in turn, reveals greater morphological and ecological diversity among the Paleocene lizards of the Qianshan Basin, including the occurrence of a varaniform (IVPP V 22767), and the reinterpretation ofAnqingosaurusas a possible burrower. Further work on the Paleocene Qianshan lizards is ongoing and the discovery of new specimens may help to solve the puzzles these strange lizards have posed.
Qianshan Basin, Anhui, Paleocene, lizards, phylogeny, taxonomic revision, CT-scan
1 Introduction
The Paleocene is a key period for our understanding of the extent of the end-Cretaceous faunal turnover and the evolution and diversification of modern lineages (Alroy, 1999, Longrich et al., 2012, Brusatte et al., 2015, Sibert and Norris, 2015), including squamates(lizards, snakes and amphisbaenians). However, Paleocene lizard fossils are very scarce outside North America (Gilmore, 1942, Estes, 1976, Sullivan, 1982, Sullivan, 1991, Sullivan and Lucas, 1996), with fragmentary material reported from Belgium (Folie et al., 2005), Romania (Gheerbrant et al., 1999), Morocco (Gheerbrant, 1994, Augé and Rage, 2006), and Bolivia (Rage, 1991). In Asia, there are currently only four known Paleocene squamate localities: a marine locality in Kazakhstan yielding sea snake remains (Averianov, 1997), one in Pakistan with Gondwanan madtsoiid snake remains (Rage et al., 2014), and the Nei Mongol Subeng section and Qianshan Basin of China. The latter two localities are the only Paleocene localities in Asia that have yielded lizard fossils (Hou, 1974, Hou, 1976, Van Itterbeeck et al., 2007).
The Qianshan Basin yields one of the important Paleocene deposits in the world. The horizons include the Early to Middle Paleocene Wanghudun Formation and the overlying Middle Paleocene Doumu Formation (Qiu et al., 1977, Wang et al., 2016). It has produced several key mammal genera, such asMimotonaandHeomys(Li, 1977), as well as six species of lizards, namelyAgama sinensisHou, 1974,AnhuisaurushuainanensisHou, 1974,AnqingosaurusbrevicephalusHou, 1976,ChangjiangosaurushuananensisHou, 1976,QianshanosaurushuangpuensisHou, 1974, andTinosaurusdoumuensisHou, 1974. In this paper we re-assess the existing material of each lizard taxon using both traditional paleontological methods and high-resolution micro-CT scanning. The new data obtained permit us to re-evaluate the position of each taxon within the current phylogenetic framework for squamates.
2 Materials and methods
All the specimens included in this paper were originally reported and named in Hou (1974, 1976) from the Early to Middle Paleocene Wanghudun and Doumu formations of Qianshan Basin (see Table 1).
To obtain additional anatomical data, several of the fossils (IVPP V 4450, 4450.1, 4452, 22767) were scanned using 225 kV micro-computerized tomography (developed by the Institute of High Energy Physics, Chinese Academy of Sciences (CAS)) at the Key Laboratory of Vertebrate Evolution and Human Origins, CAS. Two scans were conducted. The specimens, V 4450, 4450.1, and V 4451 were scanned once with a beam energy of 130 kV and a flux of 100 uA at a detector resolution of 10.98 um per pixel using a 360° rotation with a step size of 0.5° and an unfiltered aluminium reflection target, whereas the specimen V 22767 was scanned with a beam energy of 150 kV and a fl ux of 100 uA at a detector resolution of 39.20 um per pixel. A total of 720 transmission images were reconstructed for each scan in a 2048×2048 matrix of 1536 slices using a two-dimensional reconstruction software developed by the Institute of High Energy Physics, CAS. Models were rendered using VG Studio 2.1 and Mimics Research 17.0, and the 3D models were manipulated in Meshlab (http://meshlab.sourceforge.net/) and Meshmixer (http://www.meshmixer.com/).

Table 1 The Paleocene lizard-bearing horizons of the Qianshan Basin and the species yielded from each horizon
Abbreviations add.fs, adductor fossa; an, angular; an.pr, angular process; ar, articular; ar.s, articular surface; a.san.f, anterior surangular foramen; bsp, basisphenoid; cho. fs, choanal fossa; co, coronoid bone; condy, condyle; den, dentary; deps, depression; diap, diapophysis; ds. fl, descending fl ange; ecpt.ft, ectopterygoid facet; for, foramen/foramina; f.pr, facial process; fr, frontal; frpa.st, frontoparietal suture; fr.pr, frontal process; frprf.st, frontoprefrontal suture; im.s, intramandibular septum; in.pr, incisive process; io.ca, infraorbital canal; j.ft, jugal facet; j.pr, jugal process; la.ft, lacrimal facet; man, mandible; max, maxilla; max.ft, maxillary facet; Mk.gr, Meckel’s groove; mt.f, mental foramina; ne.sp, neural spine; n.pr, nasal process; occ, occipital; on. fl, orbitonasal fl ange; o.s.a.c, opening for superior alveolar canal; ot.cap, otic capsule; p.pr.an, posterior process of the angular; p.pr.as, pit for processus ascendens; p.san.f, posterior surangular foramen; pa, parietal; pal, palatine; pal. slf, palatal shelf; par, prearticular; pmax.pr, premaxillary process; popa.pr, postparietal process; prf, prefrontal; pro, prootic; prsa, presacral; przygo, prezygapophysis; pt, pterygoid; qu, quadrate; ra.pr, retroarticular process; sa, sacrum; san, surangular; sbd.rg, subdental ridge; sbd.slf, subdental shelf; sc.ca, semicircular canal; so, supraoccipital; spl, splenial; sy, symphysis; synap, synapophysis; ver.col, vertebral column; v.pr, vomerine process.
3 Systematic paleontology

Holotype IVPP V 4454, an incomplete right maxilla (Fig. 1).
Locality and horizon Fossil pit 71079(72), 300 m south of Hanlaowu (Hanhuawu), Huangpu Village, Qianshan County; Lower Member, Doumu Formation (Middle Paleocene).

Fig. 1Agama sinensisHou, 1974, the holotype IVPP V 4454, right maxilla in labial (upper) and ventromedial (lower) views
Diagnosis (of Hou, 1974) Small agamid with the following characteristics:teeth acrodont and heterodont, laterally compressed and conical, and enlarged from front to back; maxilla with one foramen at the base and developed posterior jugal process that extends postero-laterally; jugal positioned below the orbit.
Description The holotype is the posterior part of a poorly preserved right maxilla, including part of the facial process and palatal shelf, the jugal process, and fi ve teeth.
The facial process is missing except for the base, the broken edge of which demonstrates the original thickness. The lower part of its posterior margin is gently sloped and seems intact. At the level of the fi rst preserved tooth, along the junction of the facial process and the palatal shelf, there is a large foramen. The jugal process is nearly complete, and tapers into an edentulous posterolateral tip. The process is dorsoventrally compressed, providing facets for the jugal and probably the ectopterygoid. The facets face dorsally, indicating that the jugal bone was positioned right below the orbit. The lateral edge of the jugal process extends beyond that of the tooth row. In medial view, the palatal shelf is visible, but incomplete. It grades posteriorly into the horizontal jugal process.
The five acrodont teeth are all monocuspid, and are laterally compressed cones with relatively broad bases. The teeth are enlarged successively from front to back. Their bases do not overlap, although they seem tightly spaced in lingual view (well spaced in labial view). The teeth are tightly ankylosed with the maxilla, with a dorsolingual attachment so that tooth height appears greater in lingual view. In this view, the tooth is fan-shaped, with a slight central bulge. There are no discernible resorption pits, indicating that this part of the tooth series was not replaced.
Remarks Hou (1974) reported that IVPP V 4454 was from the Upper Member of the Wanghudun Formation. However, a combination of locality data and geology (Qiu et al., 1977) suggests the specimen was actually from the Lower Member of the Doumu Formation.
The taxonomic position ofAgamasinensishas been problematic for a long time due to its poor preservation. Attribution to the genusAgamawas based on ‘the strongly developed posterior part, the placement of the jugal below the orbit, the laterally compressed acrodont and heterodont teeth which are enlarged from front to back’ (Hou, 1974). Scott Moody (1980; in litt.) stated that this specimen (V 4454) more closely resembledMimeosaurus(q.v.) from Late Cretaceous Gobi Desert than theAgama/Stelliogroup species (Estes, 1983), but without any morphological discussion. Li et al. (2008) suggested that the specimen could be put intoother families because its teeth are homodont and pleurodont.
Placement of the specimen has to be based on its morphological characteristics. Among living taxa, acrodont teeth are found only in acrodontan lizards, trogonophid amphisbaenians, and rhynchocephalians. The non-overlapping arrangement of the teeth, their lingual placement, and the absence of anterior and posterior flanges excludes Rhynchocephalia, whereas jaw length (allowing for breakage) renders amphisbaenian attribution unlikely. The specimen is therefore likely to be acrodontan. It is broadly comparable with Chamaeleontidae (Evans, 2008, Anderson and Higham, 2013), and members of the “Agamidae” such as Leiolepidinae (Smith, 2011), Amphibolurinae (Hocknull, 2002) and Agaminae (Xiong and Wang, 2006, Fathinia et al., 2011, Baig et al., 2012). However, attribution to the Cretaceous Priscagaminae (Borsuk-Białynicka and Moody, 1984) cannot be ruled out, although the jugal may be fused to the maxilla in members of this group (e.g.Mimeosaurus, IVPP V 10031-10037). V 4454 is therefore too fragmentary to classify andAgama sinensisshould be treated as anomen dubium.
Holotype IVPP V 4448, a nearly complete lower jaw (Fig. 2A).
Paratype IVPP V 4449, a pair of incomplete lower jaws (Fig. 2B).
Referred specimens IVPP V 22768, an incomplete right maxilla (Fig. 2C); V 22769, part of a vertebral column (thirteen trunk vertebrae), a possible pelvic element and a possible quadrate (but see Remarks with respect to this specimen).
Type locality and horizon Fossil pit 70021, about 300 m east of Lijialaowu, Huangpu Village, Qianshan County, Anhui; Upper Member, Wanghudun Formation (Early-Middle Paleocene).
Distribution Type locality and Fossil pit 71002 (about 100 m northwest of Haixingdi, Huangpu Village, Qianshan County); Lower and Upper Members of Wanghudun Formation, Early-Middle Paleocene.
Revised diagnosis Large acrodontan that is distinguished from other living and fossil acrodontans by the following character combination: posterior position of dentary symphysis; coronoid process of the dentary present; splenial large; coronoid bone without labial process; angular strongly developed with great expansion at the level of the coronoid process; adductor fossa small; prearticular process broad; teeth sub-rectangular in lateral or medial view with low apices and lingual attachment.
Description (based on all the available specimens) The dentary is robust with a deep anterior end. There seems to be a small notch as the lower part of the dentary protrudes anteriorly. The labial surface is perforated by four foramina, running in a line from the anterior notch (V 4449). Posteriorly, the dentary extends posterior to the apex of the coronoid bone. The anterior surangular foramen is located in the suture between the dentary and the surangular. Lingually, the well-developed symphysial surface (right mandible of V 4449) is oval and anteroposteriorly elongated, facing medially. Its position on the dentary is specialized, in thatit lies slightly posterior to the anterior tip of the dentary, rather than along the margin. The subdental shelf is consistently deep until it narrows sharply from the level of the eighth/ninth tooth. Meckel’s groove is open ventrally below the symphysis but is closed by the splenial along the posterior half of the bone. The ventral edge of the dentary is inturned medially to form the fl oor of Meckel’s groove.

Fig. 2QianshanosaurushuangpuensisHou, 1974 A. the holotype IVPP V 4448, a lower jaw: photograph (A1) and line drawing (A2) in lateral view; photograph (A3) and line drawing (A4) in medial view; B. photographs of the paratype V 4449, a pair of lower jaws in lateral (B1) and medial (B2) views; C. photographs of the referred right maxilla V 22768 in lateral (C1) and medial (C2) views. The red lines in the line drawings are the sutures between different elements
There are eighteen dentary teeth, all similar in shape, but increasing somewhat in size posteriorly. The teeth are closely spaced and those in the posterior part of the row overlap to a small extent. The teeth are acrodont and laterally compressed. In labial view they are rectangular with a low central cusp, but in lingual view the teeth expand from a restricted, columnar base to a broad, shovel-like crown. There are no obvious resorption pits at the tooth bases, indicating that the marginal teeth were not replaced.
The splenial is large and subtriangular. The posterior end of the bone reaches the level of the coronoid process.
The coronoid bone bears a tall dorsal process, a well-developed posteromedial process, a posterior process, but no labial process. The anteromedial process is not preserved on any specimen. A ridge runs from the apex of the dorsal process toward the ventral end of the posteromedial process, marking the line of attachment of thepseudotemporalis posteriormuscle. The posterior process extends horizontally, overlying the dorsal margin of the surangular.
The surangular is short and deep, with a slightly concave lateral surface. Its dorsal border is broad, but overlain by the posterior process of the coronoid bone. The posterior surangular foramen is located anterior and dorsal to the articular surface. The medial adductor fossa is unusually small and shallow.
The angular is highly developed. It lies mainly on the medial surface of the mandible, where it expands gradually from the level of the tenth tooth, reaching its greatest depth just anterior to the level of the coronoid process. From there, the angular is twisted onto the lateral surface of the lower jaw and extends posteriorly to reach the articular.
The articular bears a deep prearticular process whose depth is greater than half of the depth of the lower jaw at the same level. The articular surface faces dorsally and slightly medially. There is no ridge or tubercle in front of the joint surface. The retroarticular process is broad-based, and narrows posteriorly, but it is incomplete. Only the base of the angular process is preserved, but it appears to extend posteroventrally from the base of the prearticular process.
Taken together, the features of the lower jaw ofQianshanosaurushuangpuensisare suggestive of an unusual feeding mechanism in which the main external and posterior adductor muscle mass (attaching to the surangular and adductor fossa respectively) may have been less important than the internal adductors (pseudotemporalis and pterygoideus, attaching to the coronoid fl ange and angular process respectively).
The maxilla (Fig. 2C) has a deep premaxillary process, the anterior tip of which is raised to form the articular facet for the premaxilla. A large foramen, probably the opening of the superior alveolar canal, penetrates the horizontal lamina of the process. The facial process has a wide base, and its anterior margin is angled more steeply than the posterior margin. The jugal process points laterally and posteriorly, and the jugal facet faces dorsally and laterally. The broad palatal shelf rotates clockwise along its junction with the alveolar margin, creating an acute angle between the palatal shelf and the facial process. The maxillary teeth are similar to those of the dentary.
A possible partial quadrate (V 22769) lies on the left side of the vertebral column. The slightly concave bone fragment may represent the anterolateral lamina of the quadrate, but no further interpretation is possible.
The preserved vertebral column (not fi gured) is composed of thirteen successive trunk vertebrae, nine of which are preserved on a sandstone block in ventral view, and the four preceding articulated vertebrae were prepared out from the same sandstone block. The vertebrae show no features of particular interest. The centra are procoelous, elongated, andslightly tapered posteriorly. The ventral surface of each centrum is smooth (i.e. without ridges). The neural arches of the separate four vertebrae preserve bases of the spines along the midline (the neural spines are incomplete). It is dif fi cult to tell whether the anterior margin of the arch is notched, or whether the zygosphene is present, due to the preservation. The synapophysis is small, and elongated along an axis from posterodorsal to anteroventral, on the pedicle of the prezygapophysis.
On the same side of the vertebral column as the quadrate, a small piece of bone bears a depression and may be the acetabulum of the pelvis.
Remarks The maxilla (V 22768) was described by Hou (1974), but not given a catalogue number. It was recovered from the fossil pit 71002 within the Lower Member of Wanghudun Formation. As mentioned above and in Hou’s (1974) paper, the maxilla can be assigned toQianshanosaurus huangpuensisbased on the similarity of the teeth to those of the holotype dentary. The vertebral column (V 22769) was originally included in the paratype specimen and, according to Hou (1974) was discovered together with the paratype (a label of V 4448, the holotype number, was attached to the specimen, but must be a mistake). However there is no evidence that the paratype (a pair of lower jaws) or the vertebral column came from the same individual and it is better to treat them separately. Therefore a new catalogue number has been given to the vertebral specimen. Moreover, as the vertebral column was on a separate block there is also no certainty that it is referable toQ.huangpuensis. There were three more vertebrae in the original paper (see Hou, 1974: pl.I), which are not available now.
When the posterior half of the holotype lower jaw was glued to the anterior half, it was misaligned slightly posteriorly and ventrally. However, the specimen has not been re-prepared to avoid further damage.
Hou (1974) assignedQianshanosaurusto the Iguanidae (Pleurodonta) based on the homodont and pleurodont teeth, the large dentary, the medial expansion of the coronoid bone, and the long anteromedial process of the coronoid which reaches the level of the penultimate tooth. Estes (1983) consideredQianshanosaurusto beincertae sedis, and Li et al. (2008) followed this opinion, although they agreed with Hou (1974) in interpreting the teeth as pleurodont. We regard the teeth ofQianshanosaurusas acrodont based on the complete coalescence between the teeth and the tooth-bearing bones, although they do show a lingual rather than apical attachment. The development of the coronoid process of the dentary and the homodont and acrodont (or pleuroacrodont) teeth support the recognition ofQianshanosaurusas an acrodontan lizard similar toUromastyx. This partly agrees with the opinion of Alifanov (2009) who referredQianshanosaurus,Creberidentatfrom Middle Eocene of China, andAcrodontopsis,Agamimus,Graminisaurus,Khaichinsaurus,LavatisaurusandLentisaurusfrom Middle Eocene of Mongolia to the acrodontan family Changjiangosauridae (Superfamily Uromastycoidea). The similarity of these taxa to one another, in the morphology of dentary, maxilla and teeth, does suggest they are closely related to each other and probably to the extantUromastyx. However, these taxa do not share any derived characters withChangjiangosaurus(see below), so without evidence of relationship they should not be regarded as belonging to Changjiangosauridae.

Holotype IVPP V 4453, a nearly complete left maxilla, a nearly complete left lower jaw, and a possible fragmentary pterygoid (Fig. 3).
Locality and horizon Fossil pit 71018, 150 m southwest of Chongliwu, Huangpu Village, Qianshan County, Anhui; Upper Member, Doumu Formation (Middle Paleocene).
Diagnosis (revised from Hou, 1974) Medium-sized acrodontan lizard with the following characteristics: heterodont, anterior teeth pleurodont and cone-like, and posterior teeth subacrodont and laterally compressed; maxillary acrodont teeth triangular, unicuspid, and isodont; dentary acrodont teeth tricuspid with the large main cusp slightly separated from the small lateral cuspids; lower jaw slender anteriorly; Meckel’s groove open medially.

Fig. 3 Photograph ofTinosaurus doumuensisHou, 1974 (holotype, IVPP V 4453) The maxilla and lower jaw are disarticulated from each other
Description Although the maxilla and the lower jaw are disarticulated from each other, their close approximation on the same block indicates they are likely to be from the same individual, as does their similarity in size and texture.
The maxilla bears a relatively short premaxillary process which is inturned medially to form a horizontal lamina. The anterior tip of the maxilla is broken and therefore it is impossible to determine whether there was a dorsal projection from the horizontal lamina, or whether the two maxillae met in the midline behind the premaxilla. The facial process has a broad base, with shallow anterior and posterior margins. The jugal process is long with a horizontally positioned jugal facet, but it only extends a short distance beyond the tooth row. There is a medial palatal shelf, and, probably, a maxillary lappet (Evans, 2008).
There are at least four anterior pleurodont teeth separated by a short gap from the posterior acrodont series. The pleurodont teeth are small and cylindrical due to the broken tips. The acrodont teeth increase gradually in size posteriorly and the spacing between individualteeth narrows, so that the dentition changes from being loosely spaced to closely spaced posteriorly. The acrodont teeth are laterally compressed, and triangular in labial view.
The dentary is slender and narrows anteriorly to the symphysis, but this surface is obscured by matrix. A subdental shelf is present, but it is not clear how well developed it is. The suture between the dentary and the post-dentary elements is indistinct and therefore the shape of the posterior margin of the dentary is unknown. The dentary dentition is composed of four anterior pleurodont teeth and a posterior acrodont series, probably separated by a gap. The acrodont teeth number eight or nine according to the preserved bases and the gaps between them. The second acrodont tooth, preserved unbroken, is laterally compressed and triangular in labial view. Two weak cuspids seem to be present to the sides of the main cusp.
The coronoid bone only preserves the anteromedial process which reaches the level of the third tooth from the rear. Judging from the position of the anteromedial process, there seems to be no gap between the tooth row and the dorsal process.
The posterior part of the surangular is preserved, with the posterior surangular foramen located anterior and ventral to the articular surface. The adductor fossa is small and faces medially.
The articular surface is horizontally oriented, and is divided by a weak ridge into the lateral and medial halves. A retroarticular process is not preserved.
There is a fragmentary bone posterior to the maxilla, which bears a small but deep circular depression. The depression suggests this may be a fragment of the pterygoid where the epipterygoid was articulated.
Remarks The species was diagnosed on the basis of the small, conical pleurodont teeth on the maxilla and dentary, the absence of any overlap between the acrodont teeth, the triangular maxillary acrodont teeth, the slender lower jaw, and the non-triangular facial process of the maxilla (Hou, 1974). However, the facial process ofTinosaurusdoumuensisis broken, and it is dif fi cult to determine its general shape. A re-examination ofT.lushiensisfrom China, represented by the posterior part of the maxilla preserved in medial view, found no obvious differences betweenT. doumuensisandT. lushiensis. Moreover, the genusTinosaurusseems to be a ‘waste-bin’ taxon, that probably includes more than one genus (Estes, 1983, Smith et al., 2011). The assignment toTinosaurusis generally based on the general similarity of teeth and the geological age. InT.doumuensis, the uncertainty of the tricuspid nature of the teeth cast further doubt on its attribution. Nonetheless, herein we retain the original attribution pending revision ofTinosaurusas a whole.

Material IVPP V 22767, a nearly complete right dentary, partial vertebral column with six articulating vertebrae, and a sacrum with an associated last presacral (Fig. 4).
Locality and horizon Fossil pit 71017, Yangxiaowu, Qianshan County, Anhui; UpperMember, Doumu Formation (Middle Paleocene).

Fig. 4 Varaniformes gen. et sp. indet. A. photograph (A1) and 3D rendering from CT scans (A2) of IVPP V 22767, the arrows point out the precondylar constriction of the centra; B. the partial vertebral column in right lateral view; C. the right dentary in labial (C1) and lingual (C2) views; D. the last presacral and first sacral vertebrae in dorsolateral view. B, C and D are not to scale
Description The partial vertebral column (Fig. 4A, B) consists of six posterior trunk vertebrae exposed in ventral view. The ventral surfaces of the centra are flat and without ridges, and there is a clear precondylar constriction. The specimen was CT-scanned and thereconstruction of the column reveals the lateral and dorsal parts of the vertebrae, and the depressed shape of the condyles, which are otherwise indiscernible. Each condyle faces mainly dorsally. Laterally, the synapophysis is seen to be located on the pedicle of the prezygapophysis. It is small and short mediolaterally, and its lateral end does not extend beyond the lateral edge of the prezygapophysis, which is different from the condition inVaranus(IVPP Anatomy La.b specimen). The synapophysis is elongated along an axis from posterodorsal to anteroventral. A lateral view of the column shows the low, broad neural spines. Each spine inclines posteriorly, but does not extend beyond the level of the postzygapophyses. It has a straight dorsal edge bearing an epiphysis. The posterior neural spines show gently sloped anterior edges, which, if correctly reconstructed (the possibility that it is artifact of segmentation cannot be ruled out), is different from that ofVaranuswhere the anterior edge is nearly vertical. There is no sign of a pseudozygosphene, which is present inSaniwa ensidens(Rieppel and Grande, 2007), and the anterior margin of the arch is not notched.
The last presacral and the fi rst sacral vertebrae are preserved posterior and to the right of the column (Fig. 4D), the two being loosely articulated with a slight displacement. Both centra and the left transverse process of the sacral are missing. The last presacral bears a small synapophysis which is better preserved on the right side, and a low neural spine bearing an epiphysis on the dorsal edge. The right pre- and post-zygapophysis are complete, and are oriented vertically rather than horizontally. The right sacral transverse process (diapophysis) has a forked distal end which would have articulated with the ilium in life. The neural spine is taller in the fi rst sacral than in the last presacral and also bears an epiphysis.
A nearly complete dentary (Fig. 4C) is preserved on the left side of the column, with its anterior tip overlapped by the column. The posterior and ventral margins are partially broken. The dentary is slender, with its anterior tip medially curved. The labial surface is smooth and delineated with several foramina that connect with the internal alveolar canal. The posterior margin was probably vertical as in most varaniforms. The shape and degree of development of the symphysis is unknown due to poor preservation. Meckel’s groove, opening ventrally due to the development of the subdental shelf, extends to the anterior tip of the dentary. The intramandibular septum is well developed, lacking a ventral free margin as in Anguidae. The subdental shelf is deep but a subdental ridge is lacking. The depth of the subdental shelf decreases posteriorly and ends in a narrow tip, suggesting that the dentary, at least its dorsal part, is nearly complete. The ventral border of the dentary shows a probable articulation facet for the splenial in its midsection, indicating that the splenial might extend anteriorly to at least the level of the third tooth. Medially, the deep alveolar border bears four sharp, recurved, and laterally compressed pleurodont teeth which are widely spaced along the anterior half of the dentary. Based on the length of the dentary, there would not have been more than ten teeth in total. No further detail of tooth morphology (e.g. the presence of plicidentine) is available due to the preservation and scan resolution.
Remarks This specimen (V 22767) was originally included in the holotype ofAnhuisaurus huainanensis. Hou (1974) wrote that the specimen IVPP V 4450 consists of “an incomplete skull, partial mandibles and a section of vertebral column”, although the vertebral column is actually preserved in a separate block. Our re-examination showed that the partial vertebral column was varaniform whereas the holotype skull ofAnhuisauruswas clearly not (see below). The vertebral column specimen was therefore given a new catalogue number. In Hou (1974), the sacrum and dentary were described as part of the pelvic girdle and hind limb respectively and were not given a detailed description due to the poor preservation.
The precondylar constriction, the dorsally facing condyle, and the sub-rectangular shape of the neural spines suggest the partial vertebral column is that of a varaniform lizard. The size and morphology of the two disarticulated vertebrae (the low neural spine of the presacral and the absence of the pseudozygosphene) are consistent with those of the rest of the vertebral column. The dentary is also varaniform and its size is consistent with that of the vertebrae (based on the dimensions of a specimen ofVaranussp. in the Anatomy Lab of the IVPP). Therefore the three parts of the specimen are here considered to represent a single individual.

Holotype IVPP V 4450, rostral portion of the skull (premaxilla, maxilla and dentary) (Fig. 5A).
Paratype IVPP V 4450.1, anterior part of the skull (maxilla, dentary, prefrontal and palatine) (Fig. 5B).
Locality and horizon Fossil pit 71017, southwest of Yangxiaowu, Huangpu Village, Qianshan County, Anhui; Upper Member, Doumu Formation (Middle Paleocene).
Revised diagnosis Medium to large lizard that differs from other named species in the following combination of characters: slender and long nasal process of the premaxilla; incisive process of the premaxilla present, ventrally directed, and single; broad horizontal lamina of the premaxillary process of the maxilla; smooth oblique anterior narial margin of the maxilla; fi ve pleurodont teeth on maxilla and premaxilla, and six on dentary, followed by a series of apparently acrodont teeth that increase in size from anterior to posterior.
Description (based on both the holotype and the paratype) The premaxilla is preserved on the holotype (Fig. 5C) and is rotated along its mediolateral axis so that the alveolar plate is horizontally positioned. The premaxilla is unpaired, and its nasal process is slender and triangular in section. If it were restored to its original position, the preserved nasal process would be directed dorsally rather than posteriorly, suggesting the skull was tall. The premaxillary palatal shelf is well-developed and a single, ventrally directed incisive process is located on the posterior margin of the shelf. The articulation between the premaxilla and the maxilla cannot be reconstructed due to poor preservation and the resolution of the CT images. The alveolar border bears four preserved pleurodont teeth and one vacant tooth position.

Fig. 5Anhuisaurus huainanensisHou, 1974 A. the holotype IVPP V 4450, rostral portion of the skull: photograph (A1) and 3D rendered CT image (A2) in dorsal view; B. the paratype V 4450.1, anterior part of the skull: photograph (B1) and 3D rendered CT image (B2) in dorsal view; C. 3D rendering of the premaxilla in V 4450 in posterior view; D. 3D rendering of the right maxilla of V 4450.1 in labial view; E. restored 3D rendering of the left prefrontal and palatine of V 4450.1 in posterior (E1) and anterior (E2) views; F. 3D rendering of the left dentary of V 4450 in lingual view. C, D, E, and F are not to scale
Parts of the maxillae are preserved on both specimens, but the right maxilla of the paratype (Fig. 5D) is the most complete, although it probably preserves only the anterior half. The premaxillary process bears a broad horizontal lamina, the anterior part of which is slightly upturned. The facial process is not completely preserved in any of the maxillae, but the narial margin is seen to slope gradually and smoothly into the premaxillary process, andto curve medially into a lamina that is continuous with the dorsal surface of the premaxillary process. At the base of the narial margin there is a medial foramen (opening of the superior alveolar canal) through which the maxillary nerve and accompanying blood vessels passed. In lingual view, the palatal shelf is seen to be well-developed (Fig. 5A2). The alveolar border bears fi ve anterior pleurodont teeth. Based on the dentary morphology (see below), we would expect to fi nd a series of acrodont maxillary teeth posterior to the pleurodont ones, but none were discernible despite careful analysis of the CT images. This is probably an artefact of preservation. The alveolar border extends ventrally and becomes deeper behind the pleurodont teeth.
The left prefrontal (Fig. 5E) is partly preserved on the paratype, but is displaced. The frontal process is missing, but the orbitonasal flange is complete and can be brought into articulation with the palatine. In lateral view, there is a small depression on the antero-ventral corner, which is probably the facet for the maxilla or the lacrimal. The lateral margin of the orbitonasal fl ange is smooth, suggesting that a lacrimal foramen was present between it and the lacrimal (or the maxilla).
The left palatine (Fig. 5E) is also preserved on the paratype. It is broad, with anterior vomerine, lateral maxillary, and posterior pterygoid processes, although the latter is not complete. A choanal fossa is evident on the palatal surface, and dorsally the bone bears a ridge that articulates with the prefrontal. At the base of the ridge is a groove that extends laterally into the infraorbital canal, which is closed by the maxilla. Based on the original position of the palatine, the interpterygoid vacuity probably extended between the palatines, but whether it reached the vomers is uncertain. There were no palatine teeth present.
The dentary (Fig. 5F) is best preserved in the left side of the holotype. It is rather shallow, with a more or less straight ventral border. The narrow anterior tip bears a relatively large medial symphysial surface (clear in the paratype left dentary, Fig. 5B2) that forms an anteroposteriorly elongated oval. Meckel’s groove runs along the ventral border of the dentary from the ventral margin of the symphysis to the level of the last pleurodont tooth. Posterior to that point, the groove is partially fl oored by the medially curved labial wall of the dentary and faces medially. A splenial or angular may have been present to close the groove, but there is no discernible facet for either of these elements. A subdental ridge is present. The dentition is well preserved on the holotype left dentary. There are six anterior pleurodont teeth, the last three of which are angled slightly posteriorly. The fourth is the largest, but it is not caniniform compared with the rest. Posterior to this, there are a maximum of six acrodont teeth preserved in the available specimens. These teeth increase in size from front to back and are well spaced. Each tooth is compressed laterally and is triangular in labial view. The posterior tooth crowns appear tricuspid, but this may be an artefact of the CT image.
RemarksAnhuisauruswas referred to the Agamidae by Hou (1974) based on the acrodont and heterodont nature of the dentition. The original diagnosis also included the shallow and elongated nature of the skull, the posterior inclination of the posterior teeth, fewteeth, wide premaxilla, straight maxillary anterior margin, subrectangular maxillary facial process, straight dentary lower border, broad dentary symphysis, procoelous vertebrae and conical centra. However, as noted above, the vertebral features are based on the misassociation of a varaniform specimen.
Estes (1983) cautioned that it was difficult to interpret the features due to the poor preservation of the specimens and raised the possibility ofAnhuisaurusbeing an amphisbaenian due to the short tooth row in the holotype. Li et al. (2008) consideredAnhuisaurusto beincertae sedis.
The phylogenetic position ofAnhuisaurushas remained uncertain due to the ambiguous interpretation of its anatomy. To aid clarification, we CT scanned both the holotype and paratype. The digital renderings show that the anterior marginal teeth are pleurodont, instead of acrodont, but that acrodont teeth are present posteriorly where there were thought to be no teeth (Hou, 1974, Estes, 1983, Li et al., 2008). Therefore the dentition inAnhuisaurusis heterodont. The tooth row is longer than that found in known amphisbaenians.
The heterodont dentition and the posterior acrodont teeth, as well as the configuration of Meckel’s groove and the morphology of the palatine suggestAnhuisauruscould be a basal acrodontan, as Hou (1974) suggested, but as some of these features (tricuspidy, shallow alveolar shelf, strongly implanted posterior teeth) are also found in teioid/polyglyphanodontid lizards, the phylogenetic position of this lizard remains uncertain pending the recovery of more complete material.

Holotype IVPP V 4452, an incomplete skull (Fig. 6).
Locality and horizon Fossil pit 71001, about 150 m southeast of Wangdawu, Huangpu Village, Qianshan County, Anhui; Lower Member, Wanghudun Formation (Early Paleocene).
Revised diagnosis (after Hou, 1976) Small squamate showing the following characters:deep skull; unpaired frontal with cristae cranii developed anteriorly but not meeting in the midline; interdigitated frontoparietal suture; parietal foramen present; descending fl ange of the parietal not contacting the prootic; postparietal process of the parietal slender; articular surface of mandible facing posteriorly; maxillary and dentary teeth pleurodont and tooth row short.
Description The specimen is poorly preserved and its small size also hinders the delicate preparation. It is also coated with preservative which obscures some of the details.
As preserved (Fig. 6A), the skull is deep with a short, pointed preorbital region. Although the premaxilla is missing/damaged, along with the nasals and the anterior part of the frontal, the presence of the anterior parts of the dentaries shows that very little of the snout is missing. The maxilla has a well-developed facial process and teeth are visible along the alveolar margin. The orbit is framed anteriorly by a poorly preserved prefrontal and dorsally by the preserved parts of the frontal, but all other framing elements (e.g. the posteroventral part of the maxilla, jugal, or postorbital bones) appear to be missing. The frontal is unpaired and theparietal is long, with two slender postparietal processes (supratemporal processes) that run posterolaterally along the lateral margins of the braincase, which is therefore partly exposed in dorsal view. The posterior margin of the parietal is incomplete, but the pit for the ascending process of the supraoccipital is exposed. The supraoccipital bears no ridge or crest. The lower jaw is shallow anteriorly and deepens posteriorly. The sutures between elements are unclear but the coronoid bone is visible. The quadrate is straight and apparently narrow, and its ventral end articulates with the articular of the lower jaw.

Fig. 6 Skull ofAnqingosaurus brevicephalusHou, 1976, the holotype IVPP V 4452 A. photographs in dorsal (A1) and left lateral (A2) views; B-I. CT scanning images: B. coronal section through the frontal-prefrontal suture; C. horizontal section through the frontoparietal suture; D. sagittal section through the descending flange of the parietal; E. horizontal section through the otic capsule; F. horizontal section through the basisphenoid; G. sagittal section through the quadrate-articular joint; H. coronal section through the adductor fossa; B’-H’ show the location of each section in the specimen; I. sagittal section through the region of the maxillary teeth. CT scanning images are not to scale
The specimen was CT-scanned in an attempt to clarify its morphological features. However, the images obtained were too poor to segment out individual elements and the following description is based mainly on data from individual CT section images (Fig. 6B-I). Nonetheless, these do provide some important new data.
A palatal shelf is visible on the maxilla and the maxillary teeth are revealed to be pleurodont (Fig. 6I) rather than acrodont as described in the original paper (Hou, 1976). The teeth are large and few (seven teeth in total). The cristae cranii (descending processes) of the frontal extend ventrally to a relatively greater extent anteriorly than posteriorly, providing a broad facet for the prefrontal (Fig. 6B). Posteriorly the lateral margins of the frontal are thickened rather than crested. The frontoparietal suture is strongly interdigitated (Fig. 6C). The adductor shelf of the parietal is well-developed and bears a descending fl ange (Fig. 6D), although this does not reach the braincase (prootic). The adductor surface, where the adductor muscles originate, faces laterally in the anterior part of the bone where the descending fl ange of the parietal is well-developed, but the surface faces ventrally posterior to that. The parietal table extends laterally only to a small extent, restricting the surface of origin for adductor muscles. The parietal foramen is present within the parietal.
The central pillar of the quadrate is nearly straight (Fig. 6G), and the posterior end of the pterygoid reaches it. The braincase is long and the anterior margin of the basisphenoid reaches forward beyond the level of the frontoparietal suture. The otic capsule is huge, and there is little development of the paroccipital processes (shown by the close positions of the quadrate and the otic capsule). The semicircular canals run close to the internal surface of the vestibule (Fig. 6E). The crista sellaris of the basisphenoid is well-developed, but the basipterygoid processes seem short (Fig. 6F). The basioccipital preserves a small anterior part, and the suture between the basisphenoid and basioccipital is preserved as reversed V-shaped in ventral view.
The lower jaw has a broad ventral border. Meckel’s groove is large and is walled medially by different elements, splenial anteriorly, splenial and angular in the middle, and splenial, angular and coronoid bone posteriorly. There are seven dentary teeth that are large and pleurodont. The coronoid bone bears a well-developed dorsal process, an anteromedial process and a posteromedial process, but not a labial process. The anteromedial process is longer than the posteromedial one. The splenial is large, extending posteriorly beyond the level of the coronoid process, and overlapping the angular quite extensively in its anterior part. The surangular walls a large adductor fossa laterally (Fig. 6H). The articular surface of the articularbone faces posteriorly rather than dorso-posteriorly (Fig. 6G). This suggests the quadrate met the lower jaw at an oblique angle. There is a ridge dividing the articular surface into lateral and medial halves, as well as an erect tubercle just in the front of the articular surface, to prevent the anterior displacement of the quadrate condyle. The retroarticular process is present but small, and it lies medial and ventral to the articular surface.
Remarks Hou (1976) referredAnqingosaurusto Chamaeleonidae based on its (apparently) acrodont teeth, deep skull, large prefrontal, rough sculpture, and the absence of a retroarticular process. This interpretation was followed by Estes (1983) who noted that the quadrate inAnqingosaurusappeared vertical and columnar, as in chameleons, and that the postdentary region of the mandible is enlarged. However, a recent review of chameleon history (Bolet and Evans, 2014) concluded thatAnqingosaurusdid not show any definitive characteristics of Chamaeleonidae.
The CT-scan section images reveal several key characters, such as the posteriorly oriented articular surface for the quadrate, the strongly interdigitated frontoparietal suture, the short tooth row (and therefore short preorbital skull), and the relatively large otic capsule, all of which suggest thatAnqingosaurusmay have been a burrower. Unfortunately, its postcranial skeleton remains unknown. To assess the phylogenetic position ofAnqingosauruswithin Squamata, it is fi rst compared with extant burrowing squamates.
Anqingosaurusand amphisbaenians (Zangerl, 1944, Montero and Gans, 1999, Kearney et al., 2005, Bolet et al., 2014) share the strongly interdigitated frontoparietal suture, posteriorly facing articular surface, and short tooth row. However,Anqingosaurusis distinguished from known amphisbaenians by the weaker descending fl ange of the parietal (Müller et al., 2011), laterally expanded parietal table, slender postparietal processes, unpaired frontal, welldeveloped basipterygoid processes, and large adductor fossa. Dibamids (Rieppel, 1984) and the burrowing gekkotanAprasia(Stephenson, 1962) show clear differences fromAnqingosaurus, in that their frontoparietal suture is straight, the parietal is snake-like in having descending laminae, the dentary is tube-like. The frontals are also paired in dibamids. Burrowing scincoids likeAcontias(Rieppel, 1981, Evans, 2008) also differ fromAnqingosaurusin having a straight or less interdigitated frontoparietal suture and cristae cranii meeting in the midline. The dissimilarity betweenAnqingosaurusand the burrowing anguidAnniella(Bellairs, 1950) is even greater.Anniellahas paired frontals with the cristae cranii meeting in the midline, a W-shaped frontoparietal suture, a descending flange of the parietal meeting the braincase, and an anteriorly open Meckel’s groove.Anqingosaurusmost closely resembles burrowing members of the Gymnophthalmidae (e.g.Bachia), based on shared frontal and parietal features such as the interdigitated frontoparietal suture and weak descending flange of the parietal. However, herein we treatAnqingosaurusasincertae sedisdue to its poor preservation and lack of detailed morphology. More specimens and different methods are needed to understand this species, but it is clearly not a chamaeleonid or stem-chamaeleonid.
Holotype IVPP V 4451, a pair of incomplete lower jaws and a left quadrate (Fig. 7).
Locality and horizon Fossil pit 71001, about 150 m southeast of Wangdawu, Huangpu Village, Qianshan County, Anhui; Lower Member, Wanghudun Formation (Early Paleocene).
Revised diagnosis This taxon is distinguished from all other lizards by a unique combination of characteristics: dentary elongated posteriorly, bearing a coronoid process; splenial large; coronoid bone lacking labial process; angular with a huge posterior process creating a deep posterior emargination in the lower jaw; adductor fossa reduced; prearticular process broad; retroarticular process well-developed.
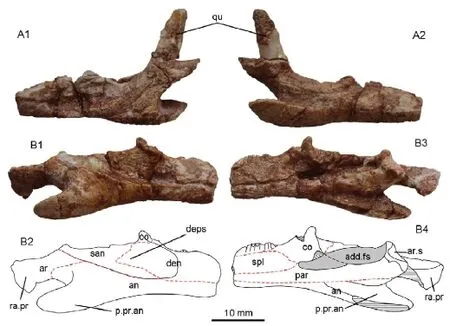
Fig. 7Changjiangosaurus huananensisHou, 1976, the holotype IVPP V 4451 A. left lower jaw and quadrate in lateral (A1) and medial (A2) views; B. right lower jaw:photograph (B1) and line drawing (B2) in lateral view; photograph (B3) and line drawing (B4) in medial view. The dashed red lines are sutures
Description In labial view (Fig. 7B), the dentary is seen to be long, extending posterior to the level of the coronoid process and having a triangular posterior margin. A coronoid process is present but it is only slightly elevated. The process bears a ridge along its anterodorsal margin, and this ridge forms the dorsal border of a depression that crosses the labial surface of the lower jaw. In lingual view, there is no development of the subdental shelf, and the seven preserved teeth are very small in relation to the depth of the underlying bone. They are tightly spaced, pleurodont, and cylindrical in shape.
The splenial is broad and would be triangular if complete. It lies dorsal to the anterior process of the prearticular and extends posteriorly almost to the level of the coronoid process.The coronoid bone has dorsal, anteromedial, posteromedial and posterior processes, but no labial process. The dorsal process is small and triangular, even allowing for damage. It bears a lateral ridge that runs anteroventrally from its tip and is continuous with the ridge on the dentary. The anteromedial process is short and deep, barely reaching below the posterior end of the tooth row. Posteriorly it grades into the fl anged posteromedial process, but this is broken on both jaws so it is unclear whether the ventromedial border of the coronoid was straight or divergent. The posterior process is also small and most of it is positioned on the medial side of the surangular. The large robust angular forms the ventral border of the mandible as preserved, and is unique in its morphology. It is lamina-like anterior to the level of the coronoid process, but expands and branches into a large fl ange which projects posteroventrally. This fl ange tapers from base to its posterior tip, and is almost as long as the articular/retroarticular process above it, creating a deep emargination in the posterior border of the mandible.
The lateral surface of the surangular bears a large deep depression that continues on to the rear of the dentary and marks the insertional area for external adductor muscles. Posteriorly, the surangular bears an upturned tubercle that lies immediately in front of the articular surface for the quadrate and would have limited forward movement of that bone. Medially, the surangular borders a narrow, but probably deep, adductor fossa. The long, laminar prearticular extends anteriorly from the articular. Anteriorly, the prearticular extends forward beyond the posterior end of the tooth row and posteriorly it floors the adductor fossa. On the articular, the surface for the quadrate faces dorsally and slightly posteriorly. It is subcircular and mediolaterally elongated. A weak ridge extends from the posterior tip of the surangular and divides the articular surface into medial and lateral parts. The laminar retroarticular process is broad and concave. The posterior half of the process is twisted so that the dorsal surface faces slightly medially.
The preserved ventral part of the quadrate is narrow and straight, and its articular condyle is not expanded.
Remarks Hou (1976) established the family Changjiangosauridae based onChangjiangosaurus, which is unique in the possession of the large fl ange-like posterior process of the angular.
Estes (1983) concluded thatChangjiangosaurusandQianshanosauruswere probably related, based on the greatly expanded projecting fl ange of the angular. However, the angular ofChangjiangosaurushas a robust, posteriorly projecting process that is separated from the main body of the angular and creates a deep notch at the back of the mandible, whereas that ofQianshanosaurusis expanded ventrally to form broad lamina. Based on tooth morphology, Alifanov (2009) includedChangjiangosaurus,Qianshanosaurus,Creberidentatfrom the Paleogene of China, and several species from the Middle Eocene of Mongolia (e.g.Acrodontopsis,Agamimus), into the Changjiangosauridae, and placed the family within an extended Uromastycoidea. As described above, the teeth ofChangjiangosaurusare different from the large, laterally compressed teeth ofQianshanosaurusand other related genera,and are instead small, cylindrical, and pleurodont. We follow Li et al. (2008) here and treatChangjiangosaurusas Squamataincertae sedis.
In the majority of lizards, the angular process, where present, is actually part of the prearticular and extends medially, ventromedially, or anteroventrally. It serves for the attachment of part of the bulkypterygoideusmuscle that wraps around the posterior part of the mandible. The large process inChangjiangosaurusis not homologous to the angular process because it is an extension of the angular rather than the prearticular, but from its position it must also have been associated with thepterygoideusmuscle. The large size of the process suggests thepterygoideuswas particularly well developed. Combined with a strong external adductor mass (as suggested by the deep labial depression), this suggestsChangjiangosaurushad a strong bite force (Daza et al., 2011, Gröning et al., 2013). However, this is not consistent with the small cylindrical teeth, and suggests this lizard had a highly specialised diet. Resolving the anomaly requires material of the skull and upper jaw as well as further comparative work among extant lizards.
4 Discussion
This revision of the Qianshan Paleocene lizards changes our understanding of the faunal composition. There are three acrodontans (Acrodonta, Iguania),Agama sinensis(nomen dubium),Qianshanosaurus huangpuensis, andTinosaurus doumuensis; the problematic heterodontAnhuisaurus huainanensis; a undetermined varaniform V 22767; an enigmatic burrower,Anqingosaurusbrevicephalus; and the bizarreChangjiangosaurus huananensis. The taxonomic diversity is higher than previously recognized. The assignment ofQianshanosaurusto Acrodonta eliminates the record of Pleurodonta (Iguania) in the Paleocene of the Qianshan Basin (Hou, 1974), whereas the recognition of a varaniform (V 22767) provides the earliest Cenozoic record of this group in China. The revision also increases the ecological diversity represented by the Paleocene lizards from China. The sharp and recurved teeth of the varaniform suggest the individual was probably a small carnivore, like its extant relativeVaranus(Pianka et al., 2004). The homodont acrodont teeth ofQianshanosaurus, combined with strong jaws and well-developedpterygoideusmuscle, may be indicative of a plant-eater, like the extantUromastyx, whereas the other acrodontans may have been insectivorous. The morphology ofAnqingosaurus(short snout, strongly interdigitated frontoparietal suture) suggests it may have been a burrower, exploring the subterranean environment. This would also be the only record of burrowing squamate in China, other thanSineoamphisbaenafrom the Late Cretaceous (Wu et al., 1993). The high morphological and ecological diversity suggests that lizards formed a successful component of the fauna in the Paleocene of the Qianshan Basin.
The similarity of the PaleoceneQianshanosaurustoCreberidentatandAcrodontopsisfrom the Middle Eocene of China and Mongolia respectively, opens the possibility thata group of acrodontan lizards similar toUromastyxwas widespread in Asia during the Paleocene-Eocene period. Moreover, other Asian records of PaleogeneUromastyx-like lizards (Uromastycinae) (Averianov and Danilov, 1996, Holmes et al., 2009, Rage and Augé, 2015) suggest this group originated in Asia.
Furthermore, the recognition of a Paleocene varaniform in the Qianshan Basin of South China provides new data on the historical biogeography of that group. Very few Paleocene occurrences of Varaniformes have been reported, and most are from North America (Sullivan, 1982, Sullivan, 1991). The new Chinese record expands the biogeographic range of varaniforms in the Paleogene and increases our knowledge of the group after the end-Cretaceous mass extinction.
In recent years, much of the paleontological work on Asian lizards has focused on the Cretaceous, especially the very rich and diverse Late Cretaceous faunas of the Gobi Desert (Gao and Norell, 2000). Very little is known of the post-Cretaceous fate of lizards in Asia. The Qianshan fossils, although fragmentary, provide an important glimpse of this fauna and suggest it was already diverse and successful in Paleocene. More work is needed to be done on these interesting Paleocene lizards, including phylogenetic analysis and functional morphology. The recovery of more material from the Paleocene Qianshan Basin and other regions is crucial in order to build up a more complete picture of the Paleocene lizard fauna of Asia.
Acknowledgements We thank Dr. Wang Yuanqing from IVPP for inviting us to contribute to this special issue and for discussing the stratigraphy of Qianshan Basin. We are grateful to Jean-Claude Rage (Muséum national d’Histoire naturelle, Paris) and Krister Smith (Senckenberg Research Institute and Natural History Museum) for providing helpful comments on an earlier version of the manuscript, and to Hou Yemao and Hao Xinxin (IVPP) for aid with the CT and stereomicroscope facilities, and software, in the IVPP lab. We also thank James Turbett (University College London), Gao Wei and Zhou Yachun (IVPP) for help with Meshlab and Meshmixer software and photographs, respectively. This work was supported by grant from NSFC (grant no. 41472018) and “973” Project (grant no. 2012CB821903). A State Scholarship Fund from the China Scholarship Council permitted Dr. Dong Liping to spend 12 months in the UK (University College London) in 2014-2015.
Alifanov V R, 2009. New acrodont lizards (Lacertilia) from the Middle Eocene of southern Mongolia. Paleont J, 43(6):675-685
Alroy J, 1999. The fossil record of North American mammals: evidence for a Paleocene evolutionary radiation. Syst Biol, 48(1): 107-118
Anderson C V, Higham T E, 2013. Chameleon anatomy. In: Tolley K A, Herrel A eds. Biology of the Chameleons. Berkely:Unversity of California Press. 7-56
Augé M, Rage J-C, 2006. Herpetofaunas from the Upper Paleocene and Lower Eocene of Morocco. Ann Paleont, 92(3):235-253
Averianov A O, 1997. Paleogene sea snakes from the eastern part of Tethys. Russ J Herpetol, 4(2): 128-142
Averianov A, Danilov I, 1996. Agamid lizards (Reptilia, Sauria, Agamidae) from the Early Eocene of Kyrgyzstan. Neues Jahrb Geol Paläont Monatsh, 12: 739-750
Baig K J, Wagner P, Ananjeva N B et al., 2012. A morphology-based taxonomic revision ofLaudakiaGray, 1845 (Squamata:Agamidae). Vert Zool, 62: 213-260
Bellairs A d’A, 1950. Observations on the cranial anatomy ofAnniella, and a comparison with that of other burrowing lizards. Proc Zool Soc Lond, 119(4): 887-904
Bolet A, Evans S E, 2014. Fossil history of chameleons. In: Tolley K A, Herrel A eds. The Biology of Chameleons. Berkely:University of California Press. 175-193
Bolet A, Delfino M, Fortuny J et al., 2014. An amphisbaenian skull from the European Miocene and the evolution of Mediterranean worm lizards. PLoS One, 9(6): e98082
Borsuk-Białynicka M, Moody S M, 1984. Priscagaminae, a new subfamily of the Agamidae (Sauria) from the Late Cretaceous of the Gobi Desert. Acta Palaeont Pol, 29(1-2): 51-81
Brusatte S L, O’Connor J K, Jarvis E D, 2015. The origin and diversi fi cation of birds. Curr Biol, 25(19): R888-R898
Daza J D, Diogo R, Johnston P et al., 2011. Jaw adductor muscles across lepidosaurs: a reappraisal. Anat Rec, 294(10):1765-1782
Estes R, 1976. Middle Paleocene lower vertebrates from the Tongue River Formation, southeastern Montana. J Paleont, 50(3): 500-520
Estes R, 1983. Sauria Terrestria, Amphisbaenia. Encyclopedia of Paleoherpetology, Part 10A. New York: Gustav Fischer Verlag. 1-249
Evans S E, 2008. The skull of lizards and tuatara. In: Gans G, Gaunt A S, Adler K eds. Biology of the Reptilia, Vol. 20 Morphology H: the Skull of Lepidosauria. New York: Ithaca. 1-344
Fathinia B, Rastegar-Pouyani N, Bahrami A M et al., 2011. Comparative skull anatomy and dentition ofTrapelus lessonaeandT. ruderatus(Sauria: Agamidae) from Ilam Province, Iran. Russ J Herpet, 18(2): 83-92
Folie A, Sigé B, Smith T, 2005. A new scincomorph lizard from the Palaeocene of Belgium and the origin of Scincoidea in Europe. Naturwissenschaften, 92(11): 542-546
Gao K Q, Norell M A, 2000. Taxonomic composition and systmatics of Late Cretaceous lizard assemblages from Ukhaa Tolgod and adjacent localities, Mongolian Gobi Desert. Bull Am Mus Nat Hist, 249: 1-118
Gheerbrant E, 1994. Les mammifères paléocenes du Bassin d'Ouarzazate (Maroc). II. Todralestidae (Proteutheria, Eutheria). Palaeontogr Abt A, 231: 133-188
Gheerbrant E, Codrea V, Hosu A et al., 1999. Découverte de vertébrés dans les Calcaires de Rona (Thanétien ou Sparnacien), Transylvanie, Roumanie: les plus anciens mammifères cénozoïques d'Europe Orientale. Eclogae Geol Helv, 92:517-535
Gilmore C W, 1942. Paleocene faunas of the Polecat Bench Formation, Park County, Wyoming Part II. Lizards. Proc Am Philos Soc, 85(2): 159-167
Gröning F, Jones M E H, Curtis N et al., 2013. The importance of accurate muscle modelling for biomechanical analyses: a case study with a lizard skull. J R Soc Interface, 10: 20130216
Hocknull S A, 2002. Comparative maxillary and dentary morphology of the Australian dragons (Agamidae: Squamata): a framework for fossil identi fi cation. Mem Queensl Mus, 48: 125-145
Holmes R B, Murray A M, Chatrath P et al., 2009. Agamid lizard (Agamidae: Uromastycinae) from the Lower Oligocene of Egypt. Hist Biol, 22(1-3): 215-223
Hou L H, 1974. Paleocene lizards from Anhui, China. Vert PalAsiat, 12(3): 193-202
Hou L H, 1976. New materials of Palaeocene lizards of Anhui. Vert PalAsiat, 14(1): 45-52
Kearney M, Maisano J A, Rowe T, 2005. Cranial anatomy of the extinct amphisbaenianRhineura hatcherii(Squamata, Amphisbaenia) based on high-resolution X-ray computed tomography. J Morph, 264(1): 1-33
Li C K, 1977. Paleocene eurymyloids (Anagalida, Mammalia) of Qianshan, Anhui. Vert PalAsiat, 15(2): 116-118
Li J L, Wu X C, Zhang F C, 2008. The Chinese Fossil Reptiles and Their Kin. Beijing: Science Press. 1-473
Longrich N R, Bhullar B-A S, Gauthier J A, 2012. Mass extinction of lizards and snakes at the Cretaceous-Paleogene boundary. Proc Natl Acad Sci, 109: 21396-21401
Montero R, Gans C, 1999. The head skeleton ofAmphisbaena albaLinneaus. Ann Carnegie Mus, 68(1): 15-80
Müller J, Hipsley C A, Head J J et al., 2011. Eocene lizard from Germany reveals amphisbaenian origins. Nature, 473: 364-367
Pianka E R, King D, King R A, 2004. Varanoid Lizards of the World. Bloomington & Indianapolis: Indiana University Press. 1-588
Qiu Z X, Li C K, Huang X S et al., 1977. Continental Paleocene stratigraphy of Qianshan and Xuancheng basins. Vert PalAsiat, 15(2): 85-93
Rage J-C, 1991. Squamate reptiles from the Early Paleocene of the Tiupampa area (Santa Lucía Formation), Bolivia. In:Suárez-Soruco R ed. Fósiles y facies de Bolivia, Vol. I, Vertebrados. Santa Cruz: Revista Técnica de YPFB. 503-508
Rage J-C, Augé M, 2015. Valbro: a new site of vertebrates from the Early Oligocene (MP 22) of France (Quercy). III -Amphibians and squamates. Ann Paleont, 101(1): 29-41
Rage J-C, Métais G, Bartolini A et al., 2014. First report of the giant snakeGigantophis(Madtsoiidae) from the Paleocene of Pakistan: paleobiogeographic implications. Geobios, 47(3): 147-153
Rieppel O, 1981. The hyobranchial skeleton in some little known lizards and snakes. J Herpet, 15(4): 433-440
Rieppel O, 1984. The cranial morphology of the fossorial lizard genusDibamuswith a consideration of its phylogenetic relationships. J Zool, 204(3): 289-327
Rieppel O, Grande L, 2007. The anatomy of the fossil varanid lizardSaniwa ensidensLeidy, 1870, based on a newly discovered complete skeleton. J Paleont, 81(4): 643-665
Sibert E C, Norris R D, 2015. New age of fi shes initiated by the Cretaceous-Paleogene mass extinction. Proc Natl Acad Sci, 112: 8537-8542
Smith K T, 2011. The evolution of mid-latitude faunas during the Eocene: Late Eocene lizards of the Medicine Pole Hills reconsidered. Bull Peabody Mus Nat Hist, 52(1): 3-105
Smith K T, Schaal S F K, Sun W et al., 2011. Acrodont iguanians (Squmata) from the Middle Eocene of the Huadian Basin of Jilin Province, China, with a critique of the taxon “Tinosaurus”. Vert PalAsiat, 49(1): 69-84
Stephenson N G, 1962. The comparative morphology of the head skeleton, girdles and hind limbs in the Pygopodidae. Zool J Linn Soc, 44: 627-644
Sullivan R M, 1982. Fossil lizards from Swain Quarry “Fort Union Formation,” Middle Paleocene (Torrejonian), Carbon County, Wyoming. J Paleont, 56(4): 996-1010
Sullivan R M, 1991. Paleocene Caudata and Squamata from Gidley and Silberling quarries, Montana. J Vert Paleont, 11(3):293-301
Sullivan R M, Lucas S G, 1996.Palaeoscincosaurus middletoni, new genus and species (Squamata: ?Scincidae) from the Early Paleocene (Puercan) Denver Formation, Colorado. J Vert Paleont, 16(4): 666-672
Van Itterbeeck J, Missiaen P, Folie A et al., 2007. Woodland in a fl uvio-lacustrine environment on the dry Mongolian Plateau during the Late Paleocene: evidence from the mammal bearing Subeng section (Inner Mongolia, P. R. China). Palaeogeogr Palaeoclimatol Palaeoecol, 243(1-2): 55-78
Wang Y Q, Li C K, Li Q et al., 2016. A synopsis of Paleocene stratigraphy and vertebrate paleontology in the Qianshan Basin, Anhui, China. Vert PalAsiat, 54(2): 89-120
Wu X C, Brinkman D B, Russell A P et al., 1993. Oldest known amphisbaenian from the Upper Cretaceous of Chinese Inner Mongolia. Nature, 366: 57-59
Xiong Y, Wang Y, 2006. Studies on skeletal system ofPhrynocephalus vlangalii hongyuanensisZhao, Jiang and Huang, 1980. Acta Zootaxon Sin, 31(2): 231-238
Zangerl R, 1944. Contributions to the osteology of the skull of the Amphisbaenidae. Am Midl Nat, 31(2): 417-454
安徽潜山古新世蜥蜴化石的分类学厘定
董丽萍1Susan E. EVANS2王 原1
(1 中国科学院古脊椎动物与古人类研究所,中国科学院脊椎动物演化与人类起源重点实验室 北京100044)
(2 英国伦敦大学学院细胞与发育生物系 伦敦 WC1E 6BT)
相对于中国和蒙古晚白垩世较知名的蜥蜴动物群,该地区古新世蜥蜴的研究非常有限。安徽潜山盆地曾报道了古新统望虎墩组和痘母组中的6种蜥蜴:中国飞蜥(Agama sinensisHou, 1974), 淮南安徽蜥(Anhuisaurus huainanensisHou, 1974), 短头安庆蜥(Anqingosaurus brevicephalusHou, 1976), 华南长江蜥(Changjiangosaurus huananensisHou, 1976), 黄铺潜山蜥(Qianshanosaurus huangpuensisHou, 1974)和痘母响蜥(Tinosaurus doumuensisHou, 1974), 但化石材料均较破碎。本文使用CT扫描技术,在最新的有鳞类分类框架下对这些属种进行了重新研究,修订了上述属种的形态学特征及其分类位置。潜山盆地古新世蜥蜴动物群具有比原认为更高的形态学和生态学分异度,如巨蜥类材料(IVPP V 22767)的发现和对安庆蜥可能为穴居型的新解释。
安徽潜山,古新世,蜥蜴,系统发育学,分类厘定,CT扫描
Q915.864
A
1000-3118(2016)03-0243-26
2016-01-13
Dong L P, Evans S E, Wang Y, 2016. Taxonomic revision of lizards from the Paleocene deposits of the Qianshan Basin, Anhui, China. Vertebrata PalAsiatica, 54(3): 243-268
国家自然科学基金(批准号:41472018)和国家重点基础研究发展计划项目(编号:2012CB821903)资助。
猜你喜欢
今日农业(2021年21期)2021-11-26 05:07:00
疯狂英语·新悦读(2021年5期)2021-06-08 01:54:10
云南画报(2020年12期)2021-01-18 07:19:24
动漫星空(兴趣百科)(2020年10期)2020-10-26 06:43:16
华南地质(2020年1期)2020-07-07 07:15:02
小哥白尼·野生动物画报(2019年12期)2019-02-28 11:46:11
化石(2018年4期)2018-12-19 06:01:48
环球时报(2018-11-14)2018-11-14 04:16:49
生物学通报(2017年3期)2017-05-12 06:22:40
当代化工研究(2016年9期)2016-03-20 16:22:17
