
旱涝交替胁迫条件下粳稻株高生长模拟与分析
2016-03-23 03:53王振昌郭相平吴梦洋栾雅珺邱让建刘春伟操信春虞颖蕾河海大学水利水电学院南京0098南京信息工程大学江苏省农业气象重点实验室南京0044
中国农村水利水电 2016年9期
王振昌,郭相平,吴梦洋,栾雅珺,陈 盛,邱让建,刘春伟,操信春,虞颖蕾,曹 宇(.河海大学水利水电学院,南京 0098;.南京信息工程大学江苏省农业气象重点实验室,南京 0044)
株高是水稻重要生长指标之一,也是构成水稻理想株型的重要因素,适宜的株高是协调产量结构、群体光合效率和抗倒伏能力的重要性状[1,2]。株高除受遗传因素影响外,也受外界环境的影响。前人研究表明,施肥量、CO2浓度、O3浓度以及地温变化等因素均会影响水稻株高等生长指标[3-6]。除上述因素外,水稻不同生育阶段进行的灌溉处理,也能显著影响水稻株高等生长性状[4,7,8]。在水稻灌排实践中,人们主要关注各生育阶段的适宜耐淹深度和耐淹(旱)历时对水稻产量、品质以及环境效益的影响[9-12],然而对于不同灌溉方式,尤其是不同生育阶段旱涝交替胁迫处理下水稻的抗倒伏性能研究较少[13]。水稻随着株高以及地上鲜重的增大,基部茎节的弯曲力矩变大,水稻倒伏几率增大[13-16],严重影响水稻的产量、品质及收割效率[14]。研究不同生育阶段旱涝交替胁迫处理对水稻株高的动态变化,以及应用生长曲线模型对株高生长进行模拟,可为估算水稻生长后期的倒伏风险提供数据支持。近年来研究人员已构建了多种用于对植物生长曲线进行描述的数学模型, 并发现多数植物的生长均符合 Logistic, Gompertz 和 von Bertalanffy 等非线性模型,且上述模型多用于模拟不同品种、基质、氮素水平以及种植密度条件下植物的生长动态[16-21], 而对于单生育阶段的旱涝交替胁迫对水稻株高生长的模拟鲜见报道。前人研究表明,一定生育阶段适度的水分胁迫可使复水后短期内作物的生理生长得以改善[22-25]。然而,植物生长曲线模型是否适合模拟不同生育阶段旱涝交替胁迫及其补偿效应产生的株高生长动态变化,尚未见报道。因此本文通过采用盆栽试验,研究水稻株高在分蘖期、拔节期旱涝交替胁迫下的动态变化,并通过 Logistic, Gompertz与von Bertalanffy 3种典型的非线性生长曲线模型对常规的浅水勤灌方式及不同生育阶段旱涝交替胁迫下水稻生长规律进行生长曲线拟合,并经过分析不同灌溉处理的株高生长速度以及不同旱涝胁迫的生长差异,揭示水稻在不同灌溉方式下株高生长规律,优选出适宜旱涝交替胁迫处理下的株高生长模型,以期为进一步模拟不同水分调控方式对水稻株高影响,以及估算水稻生长后期的倒伏风险提供理论及数据参考。
1 模型选择与数据分析
1.1 模型选择
植物生长曲线法作为趋势外推法的一种重要方法,在描述及预测生物个体的生长发育等领域中已得到广泛应用[19,20]。常用的数学模型有Logistics、Gompertz 和von Bertaffany 等模型[21,22],其中Logistics和Gompertz 模型拐点固定,且均具有饱和增长特征,而von Bertalanffy模型的拐点可变[11,12,23]。Logistic, Gompertz与von Bertalanffy等3种非线性模型的表达式及特征参数如表1所示。

表1 非线性生长模型及特征参数Tab.1 Three types of nonlinear models and characteristic parameters for fitting the growth curve
注:t为水稻移栽后天数;Y是在时间t的株高;A为水稻最大株高;K为相对生长速率参数;B为常规参数。
1.2 试验设计及株高数据测定
本试验于2013年5-10月在河海大学南方地区高效灌排与农业水土环境教育部重点实验室玻璃温室内进行(南京, 31°57′N, 118° 50′E, 海拔14.4 m),试验水稻品种为当地高产品种“南粳44”(Oryza sativa L. Nanjing44)。试验桶高为25.5 cm,上部和底部直径分别为 24和18 cm。试验用土采自附近稻田耕作层,土壤类型为黏壤土,土壤中速效氮与速效磷含量分别为50.2 和 9.58 mg/kg。所用土壤经晒干、打碎、过筛和风干后,均匀施肥,施肥量为每千克风干土尿素[CO(NH2)2]0.22 g、硫酸钾(K2SO4)0.09 g、磷酸二氢钾(KH2PO4)0.25 g、有机肥1.67 g,其中有机肥氮素含量质量分数为3.75%,装桶后土壤容重为1.29 g/cm3,田间持水率(Field capacity,FC)为 33.0%(质量含水率)。试验于2013年5月8日育秧,6月21日选择大小基本一致的3叶1心的秧苗移栽,每桶3穴,每穴移栽秧苗2株。本试验共设置5个灌溉处理,分别为对照处理(CK),分蘖期轻旱-涝-轻旱处理(T-LD),分蘖期重旱-涝-重旱处理(T-HD),拔节期轻旱-涝-轻旱处理(J-LD)和拔节期重旱-涝-重旱处理(J-HD)。试验中对照处理(CK)为浅水勤灌,除黄熟期外,各生育阶段均保持5 cm水层不变。试验中干旱胁迫通过人工称重法实现:在试验期间,通过一天2次的频率(07∶00和18∶00 各称量1 次)称取试验桶重,当土壤含水量低于或接近于灌水下限时,人工灌水至灌水上限,维持桶内土壤含水量处于相应生育阶段的灌水上限和灌水下限之间;试验中的涝胁迫通过将水稻试验桶统一放入对应水位的水箱来实现。水分胁迫开始前,各处理均为浅水勤灌。水稻分蘖期旱胁迫于2013年7月21日开始,历时 5 d,于7月26日结束,随即由旱胁迫转入涝胁迫,并持续5 d,于8月7日再次转入旱胁迫,8月12日分蘖期胁迫结束,后续转入浅水勤灌;水稻拔节期旱胁迫于2013年8月20日开始,胁迫历时5 d;8月25日由旱胁迫转入涝胁迫,胁迫历时5 d;9月2日再次转入旱胁迫,9月7日拔节期胁迫结束后随即转入浅水勤灌,并于10月29日(DAT 130)收获水稻。试验水分处理旱胁迫分为重旱(土壤含水率上下限分别为 60%FC和70%FC)和轻旱(土壤含水率上下限分别为70%FC和80%FC),涝胁迫为轻涝(在分蘖期和拔节期分别保持桶中土面距水面10和15 cm)。水稻各水分控制方案如图1所示。上述各灌溉处理随机排列,每种灌溉处理设40个重复。水稻植株移栽后,分别于DAT 24、35、40、51、59、65、70、77、94、108、118和130,随机抽取6株水稻用直尺测定株高。水稻抽穗前株高为土面至每丛最高叶尖的高度,抽穗后为土面至最高穗顶(不连芒)的高度。
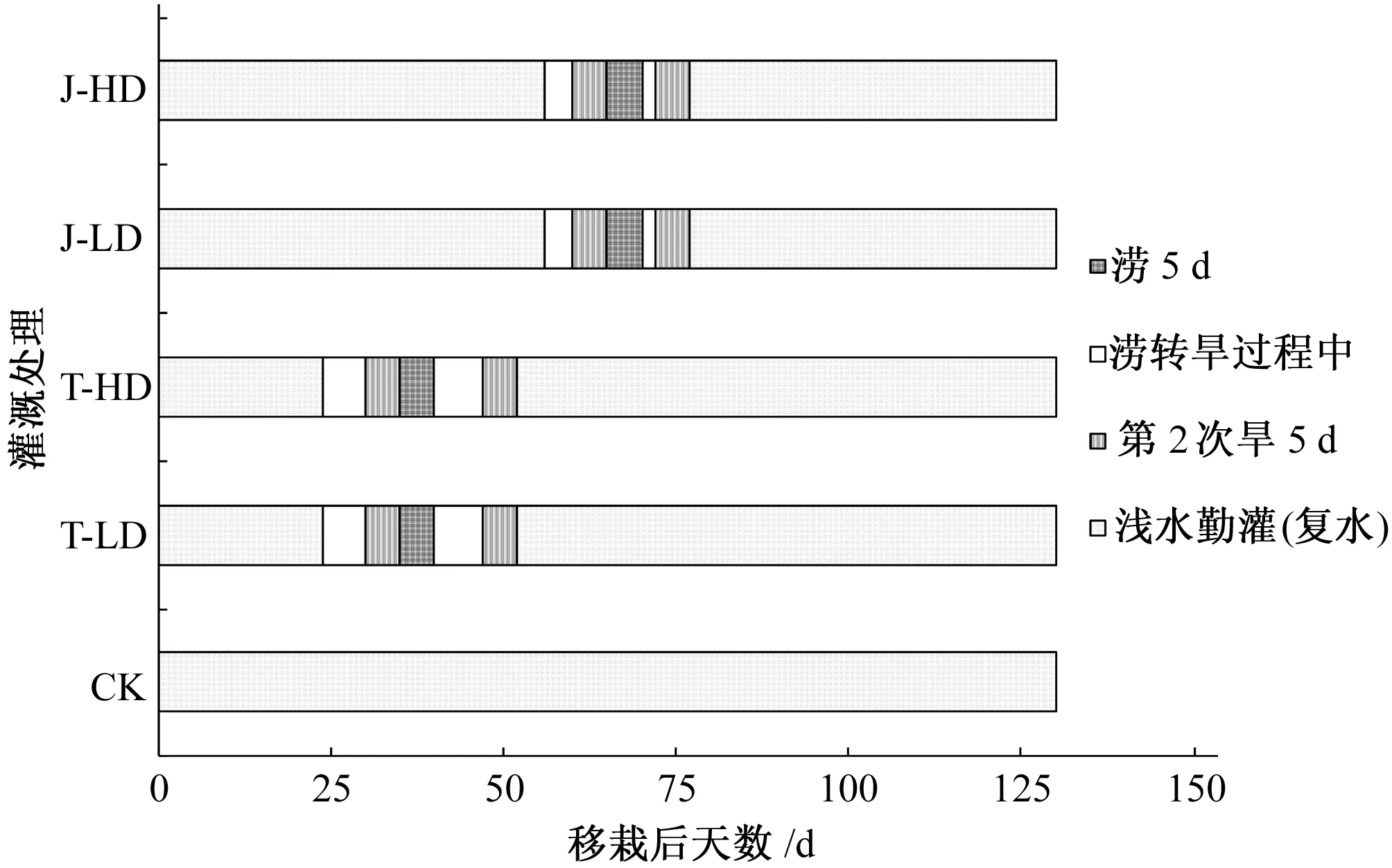
图1 灌溉处理示意图Fig.1 Diagram of experimental setup
1.3 数据统计与参数拟合
利用SPSS 13.0 (SPSS, Chicago, IL, USA) 进行方差分析与邓肯多重比较对不同灌溉处理的株高数据进行分析。选用表1中3种非线性增长模型,取各灌溉方式下65%的实际测定株高,利用SPSS 13.0 中的Nonlinear Regression功能,通过比较拟合优度的拟合优度最大估算法分别对不同灌溉处理水稻株高增长过程进行拟合,求出不同灌溉方式相应模型下的表达式及特征参数,建立不同生育阶段旱涝胁迫下水稻株高生长曲线模型。
1.4 模型评价标准
取65%的株高数据用于参数估计后,取剩下的35%的株高数据用于上述3种非直线模型实测值和模拟值的比较。修正效率系数E1、修正一致性指标d1和平均绝对误差MAE为3种模型评价指标[26]。E1反应模型预测值和实测值的符合程度,其值在-∞与1之间,数值越大表明符合程度越好,d1的值在0与1之间,数值越大,表明其一致性优越。上述3种模型评价指标的计算方法分如下:
(3)

2 结果分析与模型拟合
2.1 不同生育阶段旱涝交替胁迫下水稻株高动态变化分析
由图2知,在分蘖期第一次旱胁迫结束时(DAT 35),T-LD和T-HD的株高从40 cm(DAT24)分别增长到48.75和47.5 cm,其中干旱胁迫重的T-HD比胁迫轻的T-LD处理株高低了1.05 cm,经统计分析,两者无显著差异(p>0.05),但两处理均值均显著低于无干旱胁迫的CK处理(p<0.01);其后的涝胁迫处理 (水层深度持续保持10 cm),使得该时期T-HD的增长速度显著大于T-LD处理,但涝胁迫5 d后(DAT40)T-LD和T-HD的平均株高仍显著低于CK处理(p<0.01);随后的涝转旱过程以及其后5d的旱胁迫,使得T-HD和T-LD处理的株高直至分蘖期结束时,仍显著低于CK处理(p<0.01)。水稻拔节期旱胁迫于2013年8月20日开始(DAT 60,图1),胁迫历时5 d后(DAT 65),J-HD和J-LD处理的株高虽低于CK处理,但三者无显著差异(p<0.05),且均显著高于T-HD和T-LD处理(p<0.01);8月25日(DAT 65)J-HD和J-LD处理由旱胁迫转入涝胁迫,胁迫历时5 d,并于DAT73再次转入旱胁迫; J-HD和J-LD处理经过5 d持续的干旱胁迫后(DAT 78),J-HD株高显著低于其余4个处理(p<0.01),且T-LD处理株高已经赶超J-LD处理(74.8 cm vs 73.9 cm);拔节期胁迫结束后,随即转入浅水勤灌,DAT94测定的数据表明,T-HD(93.35 cm)和T-LD处理株高(92.35 cm)与CK(91.85 cm)无显著差异,且显著大于J-HD(83.05 cm)和J-LD处理(81.47 cm,p<0.01)。

图2 不同时期旱涝交替胁迫作用下水稻株高的变化动态Fig.2 Changes of rice plant heights under different irrigation regimes
2.2 植株生长模型的拟合与分析
利用Logistic, Gompertz和von Bertalanffy 模型对不同生育阶段旱涝交替胁迫的水稻株高进行拟合。从表2和图3可知,3种非直线生长模型的拟合度均较好(R2均大于0.83),说明3种模型均能较好地拟合各种灌溉条件下水稻生长曲线,其中全生育期浅水勤灌和分蘖期旱涝交替胁迫条件下的R2达到0.90以上,而对于拔节期的交替重旱-轻涝处理(J-HD),3种模型的R2均值却不足0.83。另外,从表2可知,3种模型预测各灌溉条件水稻株高生长过程线的最大株高、拐点高度、拐点时间均存在较大差异:Logistic 模型预测的拐点株高和拐点时间均显著大于Gompertz 和von Bertalanffy模型,而Gompertz和von Bertalanffy模型预测的水稻最大株高均显著大于Logistic 模型。结合该粳稻品种(Nanjing 44)的生长特性,其最大株高不应超过115 cm, Logistic模型预测值在此范围内,而Gompertz 和 von Bertalanffy模拟T-LD和T-HD处理下的最大株高已经超过128 cm,不符合水稻实际生长状况。

表2 2种拟合非线性模型参数估计值和拟合度Tab.2 Fitting degree and parameter evaluation ofthree fitting nonlinear models
2.3 实际观测值与拟合曲线估计值比较
在不同灌溉处理下,均用65%实测株高数据模拟出的模型预测株高,并用剩余35%的实测数据进行验证。用1∶1 作图法,比较模拟值与实测值在标准对称线两侧的分布,即分别以实测值为横坐标,以模拟值为纵坐标,作等间隔相关图,同时计算二者的修正效率系数E1、修正一致性指标d1和平均绝对误差MAE。从表3和图4可知,实测值与模拟值所建立的回归方程的决定系数达到0.85以上,且回归方程统计检验均达到极显著水平(p<0.01);经分析CK,T-LD和T-HD处理下的实测株高值和模拟株高值,可知 Logistic 模型条件下的E1、d1均大于Gompertz 和von Bertalanffy,而MAE小于Gompertz和von Bertalanffy模型,表明在CK,T-LD与T-HD灌溉方式下,用logistic模型模拟的株高生长状况与实际符合程度好,一致性优越;经分析J-LD和J-LD的实测株高和模拟株高在3种模型下E1、d1和MAE无差异,表明在J-LD和J-LD灌溉方式下3种模型模拟的生长状况无差异,但整体来看,各模型在模拟CK,T-LD与T-HD条件下株高生长状况比J-LD和J-HD处理的效果好(CK,T-LD与T-HD处理在各模型下的E1、d1均显著大于J-LD和J-HD,而MAE则小于J-LD和J-HD处理)。
2.4 不同灌溉方式下株高增长速率随时间变化状况
经实际观测值与拟合曲线估计值比较可知,上述3种非曲线模型中,Logistic 模型模拟5种灌溉方式下株高生长状态最好。通过对Logistic 模型求一阶导数,得出株高增长速率随移栽后天数变化的方程为:
(4)
式中:v(t)为株高增长速率随时间的变化方程;A为水稻最大株高;K为相对生长速率参数;B为常规参数。

图3 不同灌溉方式下水稻株高生长模型拟合Fig.3 Fitting nonliear models of rice height under different irrigation regimes
从图5可知,水稻秧苗移入大田后,由于苗期的旱涝交替胁迫,在DAT 40之前,CK,J-LD和J-HD的增长速率大于T-LD和T-HD处理;水稻进入拔节期以后,J-LD和J-HD由于受到旱涝交替胁迫,其株高增长速率明显低于CK,T-LD和T-HD处理,甚至在DAT 60以后,T-LD和T-HD处理的株高增长速率反超CK处理,表现出较强的补偿效应。
3 讨 论
生长曲线法是趋势外推法的一种重要方法,广泛应用于描述及预测生物个体的生长发育以及经济特性等领域中,其常用的数学模型有Logistics、Gompertz 和 von Bertaffany 等模型,其中Logistics和Gompertz 模型均具有饱和增长特征,在预测植物生长发育进程等方面应用最多[ 17-21]。前人研究表明Logistic生长曲线模型能较好地描述植物生长的模型[27,28],而马秋月等的研究发现von Bertalanffy模型拟合植物生长的效果比较好,并认为von Bertalanffy模型的拐点可变,而Logistic与Gompertz的拐点固定,是造成Logistic与Gompertz拟合效果差于von Bertalanffy的主要原因[19]。本研究运用Logistics、Gompertz和von Bertaffany 模型模拟了不同灌溉方式下的株高变化,结果表明,不同灌溉处理株高增长均符合渐增期-快增期-缓增期3个阶段的“S”形曲线生长的一般规律,说明株高作为生物积累量的重要指标可以反映植物的生长状况。比较水稻株高的拐点时间、拐点株高和最大株高的模型预测值与实测值的接近程度、综合预测值和实测值的线性相关性、修正效率系数E1、修正一致性指标d1以及平均绝对误差MAE,可知Logistics 模型能更好地模拟不同灌溉方式下水稻株高变化动态,这与Wu等[29]的研究结果相一致,而不同于马秋月等的研究成果[19]。
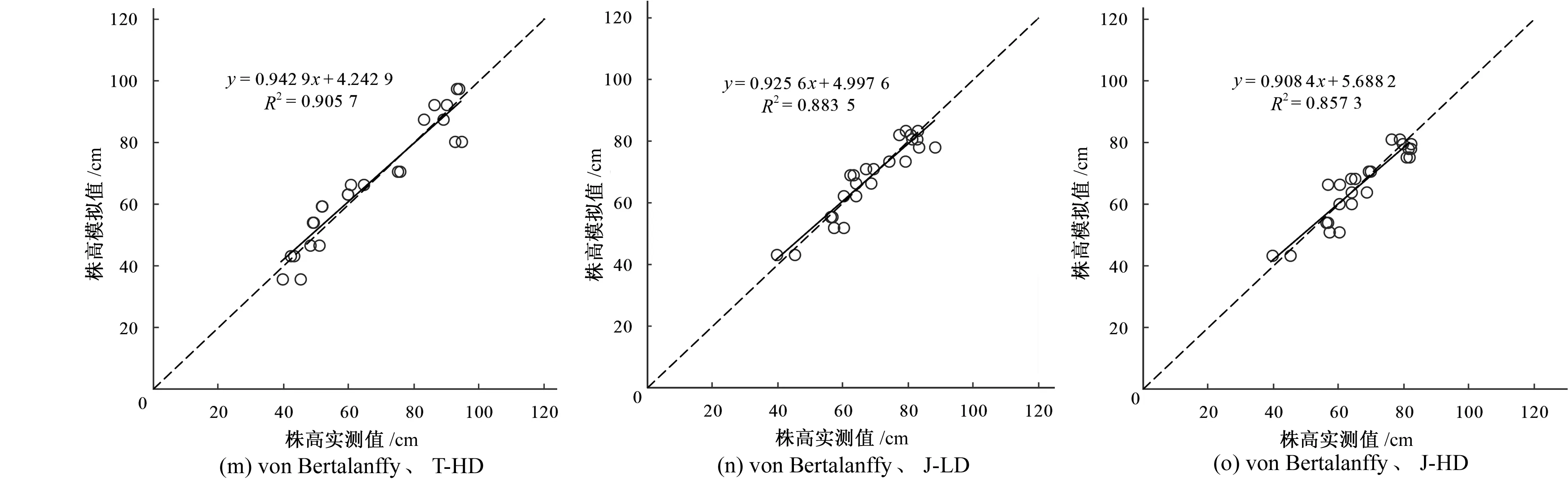
图4 不同灌溉方式下水稻株高模拟值与实测值的比较Fig.4 Comparison of measured rice height and estimated rice height
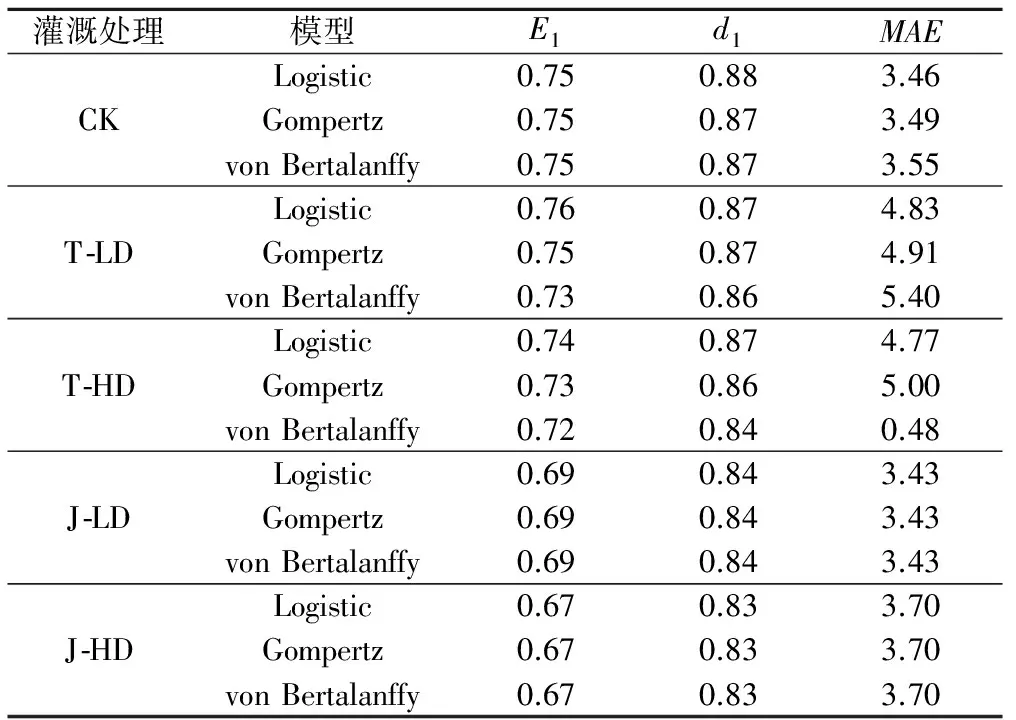
表3 3种非线性模型实测值和模拟值相关性比较Tab.3 Correlation between simulated values and observed values under three different nonlinear growth models
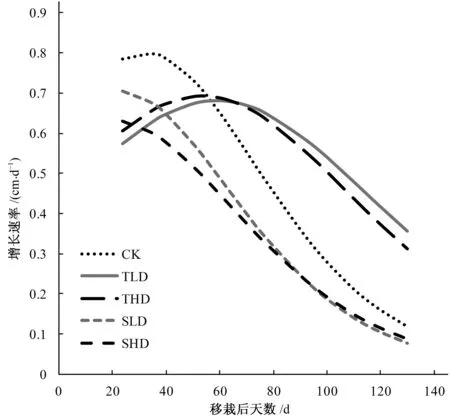
图5 不同灌溉方式下株高增长速率图Fig.5 Growth rates of rice height under different irrigation regimes
根据Logistics模型曲线拟合不同灌溉方式下的株高生长的数据(表2、表3、图3和图4 ),可知Logistic模型曲线模拟常规灌溉(CK)以及分蘖期旱涝交替胁迫的灌溉处理(T-LD和T-HD处理)的效果优于拔节期旱涝交替胁迫的处理(J-LD和J-HD处理)。这可能解释为:分蘖期旱涝交替胁迫下的株高可以通过后期补偿效应来保持 Logistic 模型推导过程[17]相对生长率K和常规参数B在生育期前后保持协调的状况有关;而对于水稻拔节期旱涝胁迫,由于株高生长受限且后期的补偿效应较弱,不能保证上述参数的前后协调。
从表2和图3可知,水稻最大株高以及拐点时间受旱涝交替胁迫实施的时期影响较大,并且不同时期的旱涝交替胁迫下拐点株高存在显著差异,在分蘖期旱涝交替胁迫处理的拐点株高高于全生育期浅水勤灌的CK处理,而拔节期旱涝交替胁迫处理的值为最低,这说明不同时期的旱涝交替胁迫显著影响植物的生长状况。通过对Logistic 模型求一阶导数,得出株高增长速率随水稻移栽天数的方程:分蘖期水稻经历旱涝交替胁迫后,能在生育后期产生一定的补偿效应,促进株高的增长;拔节期的旱涝交替胁迫后进行复水,其株高反而小于整个生育期浅水勤灌的处理,这与前人的研究表明水稻分蘖期旱后复水的后效性大于拔节期的结论相一致[7,8,22,23]。近期我们的研究成果也表明,生育早期旱涝交替胁迫后水稻比常规灌溉较早形成通气组织、较大的导管面积和通气面积以及较大的光合速率是水稻出现明显补偿效应的原因[24,25]。
Logistics模型相对于其他2个生长模型可以较好地模拟出不同灌溉方式下植物的生长状况(表2、表3和图3、图4);通过对Logistics模型一阶求导得出的株高生长速率随时间变化的方程,也可以较好地模拟出分蘖期旱涝交替胁迫在生育后期对株高的补偿效应。然而通过图5可知,生育后期模型模拟出T-LD和T-HD生长速率与实际情况不符,因为在该时期,尤其是DAT 100以后,实际生产中株高的生长已经停止,而模型模拟出的株高生长速率还有较大的值,这表明Logistics模型在模拟不同阶段的旱涝交替胁迫对株高的影响时还存在一些缺陷,还需要进一步改进模型,使得该模型能够更准确地模拟不同旱涝交替胁迫下植株的生长动态,为估算水稻生长后期的倒伏风险等情况提供理论及数据参考。
□
[1] 袁隆平.杂交水稻超高产育种[J].杂交水稻, 1997,12(6):1-3.
[2] 杨守仁,张步龙,王进民, 等. 水稻理想株型育种的理论和方法初论[J]. 中国农业科学, 1984,(3):6-13.
[3] 赵轶鹏, 邵在胜, 王云霞, 等.大气 CO2和O3浓度升高对汕优63生长动态、物质生产和氮素吸收的影响[J]. 生态学报,2015,35(24):8 128-8 138.
[4] 杨长明, 杨林章,颜廷梅,等. 不同养分和水分管理模式对水稻抗倒伏能力的影响[J]. 应用生态学报, 2004,15(4):646-650.
[5] Shi G Y,Yang L X,Wang Y X,et al. Impact of elevated ozone concentration on yield formation of four Chinese rice cultivars under fully open-air field conditions[J]. Agriculture, Ecosystems and Environment, 2009,131(3 /4):178-184.
[6] Zhu C, Ziska L H, Sakai H, et al. Vulnerability of lodging risk to elevated CO2and increased soil temperature differs between rice cultivars[J]. Eur. J. Agron., 2013,46:20-24.
[7] 郝树荣,郭相平,王文娟.旱后复水对水稻生长的后效影响[J].农业机械学报 2010,41(7):76- 79.
[8] 肖梦华, 胡秀君, 褚琳琳. 水稻株高生长对旱涝交替胁迫的动态响应研究[J]. 节水灌溉,2015,(9):15-22.
[9] 吕 露, 冯常萍, 崔远来. 水稻不同节水灌溉模式的水分利用效率比较-以荆门、桂林为例[J]. 节水灌溉,2011,(11):15-18.
[10] 郭相平, 袁 静, 郭 枫, 等. 水稻蓄水-控灌技术初探[J].农业工程学报,2009,25(4):70-73.
[11] 郭相平,张展羽,殷国玺.稻田控制排水对减少氮磷损失的影响[J]. 上海交通大学学报:农业科学版,2006,24(3):307-310.
[12] 郭相平, 袁 静, 郭 枫, 等. 水稻蓄水-控灌技术初探[J]. 农业工程学报, 2009,(4):70-73.
[13] 郭相平, 甄 博, 王振昌. 旱涝交替胁迫增强水稻抗倒伏性能[J]. 农业工程学报, 2013,29(12): 130-135.
[14] 李小坤, 李云春, 鲁剑巍, 等. 强降雨致洪涝灾害下不同因素对水 稻倒伏的影响[J].自然灾害学报, 2012,21(6):99-103.
[15] 彭世彰, 张正良, 庞桂斌. 控制灌溉条件下寒区水稻茎秆抗倒伏力学评价及成因分析[J].农业工程学报,2009,25(1):6-10.
[16] Setter T L, Laureles E V, Mazaredo A M. Lodging reduces yield of rice by self-shading and reductions in canopy photosynthesis[J]. Field Crop Res, 1997,49:95-106.
[17] 莫惠栋. Logistic方程及其应用[J].江苏农学院学报, 1983,4(2):53-57.
[18] 钱永强,孙振元,韩 蕾,等. 基于Logistic 与Gompertz 非线性模型的野牛草克隆生长模拟与分析[J].中国农业科学, 2011,44(11):2 252-2 259.
[19] 马秋月, 陈赢男, 渠纪腾, 等. 簸箕柳种内杂交F1群体株高生长曲线的拟合[J]. 南京林业大学学报, 2013,37(4):13-16.
[20] 姚克敏,邹江石,买 苗,等.两系法杂交水稻株高变化规律及其与气象条件的关系[J]. 杂交水稻(增刊), 1999,14:49-51.
[21] 赵雨明,卢桂宾,贺义才.帅枣系品种枝条果实生长发育动态[J].东北林业大学学报,2011,39(9):45-48.
[22] 郝树荣,郭相平,王为木,等. 水稻分蘖期水分胁迫及复水对根系生长的影响[J].干旱地区农业研究, 2007,25(1):149-152.
[23] 郝树荣, 郭相平, 张展羽, 等. 水稻根冠功能对水分胁迫及复水的补偿响应[J]. 农业机械学报, 2010, 41(5):52-55.
[24] 甄 博, 郭相平, 陆红飞. 旱涝交替胁迫对水稻分蘖期根解剖结构的影响[J].农业工程学报, 2015,31(9):107-113.
[25] 陆红飞, 郭相平, 甄 博, 等. 旱涝交替胁迫条件下粳稻叶片光合特性[J]. 农业工程学报, 2016,32(8):105-112.
[26] Legates DR, McCabe G J. Evaluating the use of 'goodness-of-fit'measures in hydrologic and hydroclimatic model validation[J]. Water Resour Res, 1999,35:233-241.
[27] Niklas K L.Plant allometry:the scaling of form and process[M]. Chicago:University of Chicago Press,1994.
[28] West G B,Brown J H,Enquist B J.A general model for ontogenetic growth[J]. Nature, 2001,413:628-631.
[29] Wu R L,Ma C X,Yang M C K,et al.Quantitative trait loci for growth trajectories in Populus[J].Genetical Research, 2003,81(1):51-64.
猜你喜欢
水科学进展(2022年4期)2022-09-14
中国农业科学(2022年8期)2022-05-16
今日农业(2021年6期)2021-11-27
水利规划与设计(2020年1期)2020-05-25
人民长江(2019年12期)2020-01-02
吉林农业·下半月(2017年7期)2017-07-15
农民致富之友(2017年6期)2017-05-10
动物营养学报(2015年10期)2015-12-01
亚热带资源与环境学报(2015年1期)2015-01-22
江苏农业科学(2014年8期)2014-10-23
