
营养生长期水分胁迫和氮素对大豆干物质及产量的影响
2016-03-23 00:59褚丽丽
节水灌溉 2016年7期
褚 丽 丽
(黑龙江大学水利电力学院,哈尔滨 150030)
土壤水分与氮素营养是影响作物生长和产量构成的两个重要因素。近年来,基于对作物水分胁迫补偿效应技术研究表明[1-3],水分胁迫能够改变作物的光合产物在不同组织器官的分配,一定程度的水分亏缺在抑制作物生长的同时不对产量的形成构成威胁,但使作物产量产生补偿效应的水分胁迫受水分胁迫时期、胁迫强度、胁迫历时以及土壤肥力等多种因素的限制。Nowak M A、张志伟[4,5]等认为在作物营养生长期进行调控水分,可以促进根系生长,而水分胁迫时间和强度也会使根冠的相对累积速率发生变化,使根系的生长速率大于冠,水分胁迫对根系的促进作用,便于作物生殖生长期能够获得更多水分和养分,促进光合产物向籽粒调运,从而提高了作物的经济产量[6,7],这是作物对水分逆境的一种适应性反应。在干旱逆境下,氮素能够参与和调节作物的适应、伤害和修复等各种生理生化过程,通过积累渗透物质、减小气孔导度、提高ABA含量等变化适应干旱,减少水分胁迫对生长速率、叶面积及产量等的抑制效应[8,9],因此氮素对干旱复水后的作物后续生长能力的提高具有重要作用。
水分胁迫和氮素对作物生理生长都具有显著影响,因此有必要深入研究在水分胁迫和氮素共同调控下,作物干物质和产量的变化规律。目前,较少有人研究营养生长期水分胁迫和氮素交互作用对作物后续生长的影响,为此笔者进行了大豆营养生长期水分胁迫——复水和氮素施加对大豆干物质及产量影响试验,试验把水分胁迫强度、胁迫历时和氮素营养三者有机结合起来,研究大豆干物质和产量对营养生长期水分胁迫和氮素交互作用的响应机制,探讨水氮互作对干物质积累的后效性和对大豆产量的影响,并寻求获得大豆产量补偿的水分胁迫因子和氮素营养的合理阈值区域,为实现水、氮高效利用和大豆高产提供理论依据。
1 材料与方法
1.1 材料与种植
试验于2013年6-10月和2014年6-10月在黑龙江省庆安县试验基地进行。该地属于温带半干旱季风气候,多年平均降水量为454.5 mm,春季风大少雨,5、6月的蒸发量达409.4 mm,接近全年降水量。当地耕层土壤为黑钙土,0~20 cm土层土壤有机质量为45.2 g/kg、全氮为1.9 g/kg、全磷为0.6 g/kg、全钾为23.5 g/kg, pH值为7.06,田间持水率为23%~25%(重量含水率),土壤干密度为1.25 g/cm3。采用盆钵试验,试验设在移动式防雨棚内进行,供试盆钵采用聚乙烯塑料盆,盆钵顶部内径28 cm,盆底内径21 cm,高35 cm。供试土壤为黑钙土,经晒干、打碎、过筛,均匀施肥后放入盆中,每盆基肥用量:氯化钾2.2 g,磷酸二氢钙4.5 g。供试大豆品种为黑河35号。
1.2 试验设计
试验设3个因素:水分胁迫强度(轻度胁迫L,相对含水率为60%;中度胁迫M,相对含水率为50%;重度胁迫S,相对含水率为40%)、水分胁迫历时(7、14和21 d)和氮肥施用量(低肥N1,23 kg/hm2;中肥N2,92 kg/hm2;高肥N3,184 kg/hm2),氮肥为尿素。试验采用完全设计方法,共27个处理,3次重复,设计方案如表1所示。试验同时设充足水分对照处理,该处理在整个生育期土壤相对含水量都保持在75%~80%,施氮量为最高水平184 kg/hm2。
大豆种子于5月末种植在盆钵中,土壤水分采用感量为1 g的DY20K电子天平称重控制,每天17∶ 00称重灌水,使各处理稳定在设计土壤含水量范围内。大豆出苗后第7 d定苗,每盆留长势相近的幼苗3株开始进行控水处理,在规定控水时间结束后,恢复到正常的土壤含水量水平。
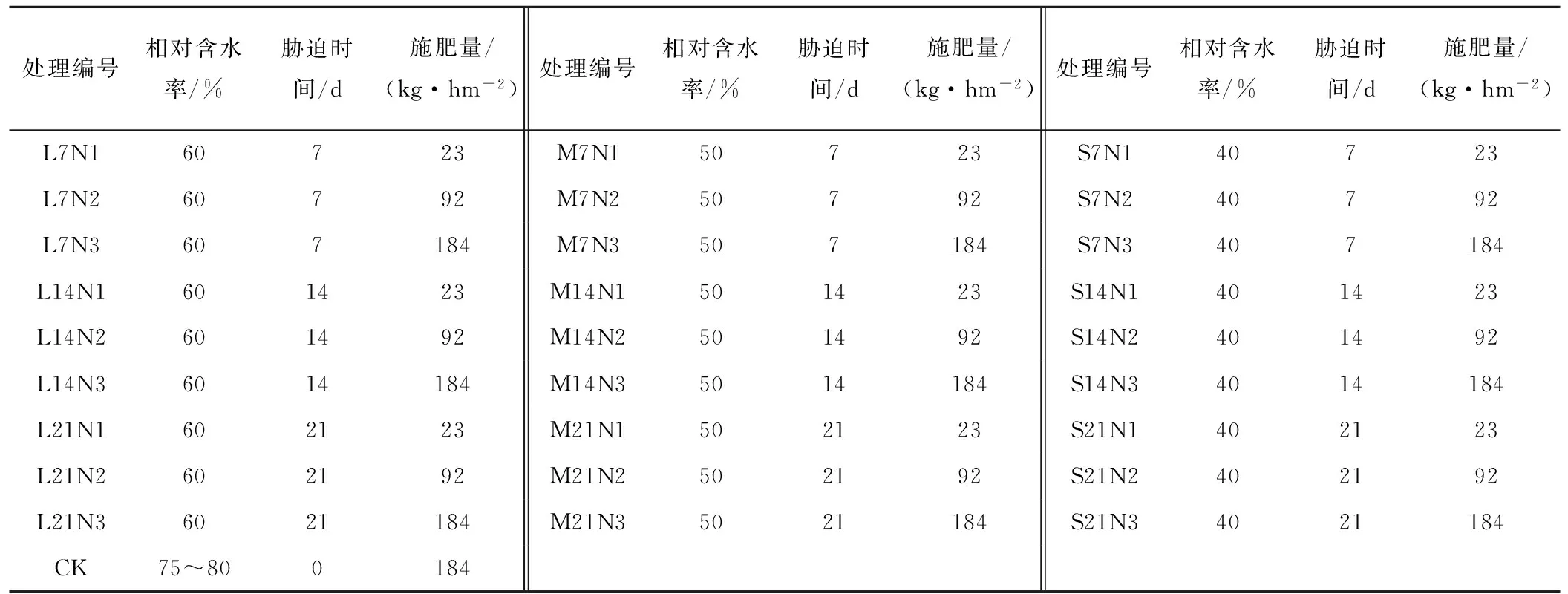
表1 试验设计方案Tab.1 Experimental design
1.3 测定项目及方法
大豆成熟后,剥出豆粒测产称重。同时用水将盆土充分浸泡,将土壤连同植株轻轻倒出,然后用流水慢慢冲净根上所有附泥,在子叶节处将植株剪断,分为根、茎两部分。用吸水纸吸干茎表面水分,然后用电子天平称量茎鲜重,采用同样方法测定根鲜重。根冠比=根鲜重/茎鲜重。
采用Excel 2007、SPSS 12.0进行数据计算、绘图及统计分析,图中数据均为各处理数据的平均值±标准差。
2 结果与分析
2.1 7 d水分胁迫和氮素对大豆干物质及产量的影响
图1可以看出在大豆苗期7 d干旱条件下,复水后同种氮素处理的不同强度的水分胁迫对大豆植株地下、地上干物质积累影响并不显著(P>0.05),干物质并未因水分胁迫强度的增强而产生反弹式增长。但同种水分条件下,不同氮素处理的地上干物质量差异却极为显著(P<0.05),随着施氮量的增加地上干物质量明显提高,轻、中和重度胁迫下的地上干物质量的变异系数分别为31%、18%和17%;与地上干物质显著增加的现象相反,各种氮素处理的地下干物质差异不显著(P>0.05)。由于地上和地下干物质增幅不同,导致根冠比产生较大变化,从图1可以看出根冠比随着施氮量的增加而明显减小(P<0.05),随着水分胁迫度的增加而略有增大,这一试验结果表明,在大豆苗期短时干旱对增加地下干物质贡献不大,但增加施氮量却有助于提高地上干物质的累积量。

图1 7 d水分胁迫复水后,胁迫强度和氮肥对大豆地上、地下干物质及R/S的影响Fig.1 Under the conditions of 7 d water stress re-watering, effects of nitrogen and water stress intensity on the ground, underground dry matter and root/shoot ratio of soybean
由于土壤水分条件和氮素的使用量改变了光合产物在根、冠间的分配比例,尤其是施氮量的增加提高了地上干物质的累积,因此也改变了大豆籽粒的产量。图2可以看出,水分胁迫对产量的影响不显著,但不同施氮量对产量影响明显,低氮、中氮处理也未出现补偿现象,只有高氮处理超过了对照水平。回归分析表明,大豆产量与水分胁迫强度呈负相关,而与施氮量和水氮互作效应成正相关,二次回归方程为:y=66.31-56.80x21+0.07x22+5.15x1x2(其中x1为土壤相对含水量,x2为施氮量)。方差分析得F拟合=13.191**[F拟合0.05(9,10)=3.2,F拟合0.01(9,10)=4.91](**表示达到F0.01显著水平),复相关系数R=0.942 2**,试验所建立的模型与实际值拟合程度较好。T显著性检验结果分别为Tx1=-0.35,Tx2=2.04,Tx1x2=1.29[T0.05(9)=2.26,T0.01(9)=3.02]。虽然试验中氮素对产量影响较为明显,高氮处理的产量都超过了对照水平,但统计分析表明水、氮及其互作效应对大豆产量的影响未达到显著水平,这说明7 d的水分胁迫对产量影响并不显著。

图2 7 d水分胁迫复水条件下,水分胁迫强度和氮对大豆产量的影响Fig.2 Under the conditions of 7 d water stress re-watering, the effects of nitrogen and water stress intensity on yield of soybean
2.2 14 d水分胁迫复水和氮素对大豆干物质及产量的影响
图3表明,当水分胁迫时间延续到14 d之后,在同一氮素水平下,地下和地上干物质随着水分胁迫强度的增强而明显增加,差异性均达到了显著水平(P<0.05)。其中N1、N2、N3处理的地上干物质的变异系数分别为7.7%、2.2%、8.3%,而地下干物质的变异系数更为显著10.0%、13.7%、17.0%,因此同一氮素水平下的根冠比呈现增长的趋势,差异显著(P<0.05),这说明干旱历时是作物干物质产生反弹的另一个关键因素,只有在水分胁迫强度和胁迫历时均达到一定程度后,干物质才产生较明显的补偿积累现象。

图3 14 d水分胁迫复水后,胁迫强度和氮肥对大豆地上、地下干物质及R/S的影响Fig.3 Under the conditions of 14 d water stress re-watering, effects of nitrogen and water stress intensity on the ground, underground dry matter and root/shoot ratio of soybean
从图3还可以看出在同一干旱程度下,不同氮素水平处理的地下和地上干物质量也都明显的提高(P<0.05),轻度、中度和重度水分胁迫处理的地上干物质的变异系数均超过了10%,分别为18.0%、18.0%和17.0%,而地下干物质的变异系数为9.0%、14.0%和13.7%,从根冠比的变化中可以看出,随着氮素施用量的增加,根冠比显著降低(P<0.05),这说明氮素的增加更有利于地上干物质的累积。此外水氮的交互作用明显提高了大豆干物质的累积量,S14N3的地上和地下干物质分别是L14N1的1.87和1.76倍,高于单一因子的贡献率(S14N3是L14N3的1.23和1.53倍,是S14N1的1.55和1.35倍)。
图4表明,当水分胁迫延至14 d时,随着水分胁迫强度的增强和施氮量的增加,大豆籽粒产量明显提高。高氮处理的产量产生了显著的补偿效应, S14N3的籽粒产量达到了最大值82.93 g/盆;M14N2和S14N2处理的产量也超过对照水平,产生了补偿现象;但低氮各水分处理的产量仍处于50~60 g/盆的区域。产量与水分胁迫强度和氮肥的二次回归方程为:y=61.58-43.18x21-0.09x22+6.60x1x2。方差分析得F拟合=21.89**,复相关系数R=0.964 0**,结果表明大豆产量与施氮量和水分胁迫强度的相关性进一步增强,R和F值明显增大。T显著性检验结果分别为Tx1=-2.69*,Tx2=-0.732,Tx1x2=2.85*(*表示达到T0.05显著水平)。结果表明,当水分胁迫时间延长至14 d后,水分胁迫强度对产量的影响达到了显著水平,这说明水分胁迫历时是产量产生补偿效应的主要因子之一;氮对产量影响仍不显著,但水氮互作效应对大豆籽粒产量影响的显著性要大于水分单因子,本试验数据也表明水氮交互作用对产量的显著性影响,如S14N3的籽粒产量是L14N1的1.62倍,高于水分胁迫(S14N3是L14N3的1.37倍)单一因子的贡献率。

图4 14 d水分胁迫复水条件下,水分胁迫强度和氮素对大豆产量的影响Fig.4 Under the conditions of 14 d water stress re-watering, the effects of nitrogen and water stress intensity on yield of soybean
2.3 21 d水分胁迫复水和氮素对大豆干物质及产量的影响
当把水分胁迫时间进一步延长至21 d时,图5表明,在低氮N1处理下随着水分胁迫强度的增加,地上和地下干物质都有所增加(P<0.05),变异系数分别为8.5%和13.4%,地下干物质增幅更大,这说明低氮水平下,加重水分胁迫还有利于干物质的积累。但是当施氮量增加到92 kg/hm2时,在轻度、中度水分胁迫下,地上和地下干物质的累积量还在增加,但在重度水分胁迫下,地下干物的累积量略有增加,但地上干物质的累积量已经开始下降,S21N2的地上干物质为M21N2的91%,这说明当水分胁迫时间和强度增强到一定程度时(胁迫时间延至21 d,相对含水率降至40%时),增施氮肥会增强土壤的生理干旱,不利于地上干物质的累积。

图5 21 d水分胁迫复水后,胁迫强度和氮肥对大豆地上、地下干物质及R/S的影响Fig.5 Under the conditions of 21 d water stress re-watering, effects of nitrogen and water stress intensity on the ground, underground dry matter and root/shoot ratio of soybean
当施氮量增加到184 kg/hm2时,轻、中度水分胁迫的地上和地下干物质增幅较小,差异不显著(P>0.05);而重度水分胁迫的地上和地下干物质都有了显著降低(P<0.05), S21N3处理的地上干物质仅为M21N3的0.69,降到了低氮处理的水平,而且地下干物质也降至M21N3的89%,降幅小于地上干物质,这进一步证明在高强度水分胁迫下,增施氮肥对干物质积累不利。

图6 21 d水分胁迫复水条件下,水分胁迫强度和氮素对大豆产量的影响Fig.6 Under the conditions of 21 d water stress re-watering, the effects of nitrogen and water stress intensity on yield of soybean
图6表明,当水分胁迫延至21 d时,低氮处理的产量变化较小;高氮的重度水分胁迫处理的产量显著降低,S21N3的产量降到58.73 g/盆,但高氮的轻、中度水分胁迫处理的产量都有所升高,超过对照水平,产生明显的补偿性,L21N3产量为74.18 g/盆,是S21N3产量的1.26倍。产量与水分胁迫强度和氮肥的二次回归方程为:y=51.18-22.36x21-0.36x22+11.65x1x2。方差分析得F拟合=25.56**,复相关系数R=0.968 9**,R和F值增大,模型与实际值拟合程度更好。T显著性检验结果分别为Tx1=-1.34,Tx2=-2.98*,Tx1x2=4.95**,检验结果表明,在长期水分胁迫下,水分胁迫强度对产量影响的显著性降低,氮素对产量产生显著的负效应,在重度水分胁迫下增施氮肥会降低大豆产量;水氮互作效应对产量产生显著的正效应,在中、轻度水分胁迫下,提高施氮量能提高水氮交互作用对籽粒的贡献率。
3 讨 论
较高的生物量是作物高产的前提,而物质的累积又是以养分的吸收为基础[10]。因此具有吸水吸肥功能的根系和具有光合功能的叶冠对作物产量形成具有重要影响。当环境条件对作物生长不利时,根、冠在生长过程中存在竞争关系[11],因而水分胁迫和氮肥亏缺会显著影响根冠干物质生产总量及其在根、冠间的分配比例。葛体达[12]等认为整个生育期相同程度水分胁迫条件下,根、冠生长曲线的高低、陡度会发生改变,但整体生长趋势不变,根冠比随着生长曲线的变化而改变,在干旱条件下同化物对根系的分配会有所增加,以恢复根系结构,维持其最低限度的功能发挥[13]。在本试验中,根系干物质随着水分胁迫强度的增强和历时的延长而明显增大,根冠比呈增大趋势,而氮肥的施用量并不会影响这一趋势。杨建昌、岳文俊[14,15]等认为无论水分充分与否,增施氮素对地上部分茎叶生长的促进作用都大于根系,使根冠比值减小,这与本研究相一致,在干旱条件下增施氮肥会降低根冠比,但在水分胁迫对根冠比的促进作用下,不同水分胁迫处理下的根冠比降低幅度不同,短历时轻度水分胁迫处理的根冠比值降幅较小,但随着水分胁迫的增强,增施氮肥反而会提高根冠比,这也许是水分胁迫对根系的正效应大于氮肥对叶冠的正效应。由此可见水分胁迫和氮肥对根冠比的正、负效应可以相互抵消,所以孙群等[16]等认为水分胁迫下氮素对根冠比的影响不明显。
作物干物质生产能力和同化物向经济器官运转的能力是作物产量形成的两个关键因素。陈晓远[17]等指出,当根、冠生长比例协调,结构与功能处于均衡状态时,资源利用效率较高。本试验营养生长期适度的水分胁迫促进根系生长,而氮肥增施又提高复水后茎叶的补偿生长量,使根冠处于平衡生长的态势;而且在干旱条件下,作物对水分逆境的适应性反应是更多地将贮存物质向穗部运转[18],有利于作物产量的提高。因此在本试验中,大豆产量随着营养生长期水分胁迫的增强和施氮量的增加而提高,但水分胁迫严重时,增施氮肥会抑制大豆的生长,干物质和产量均会降低。总之,作物营养生长期的水分胁迫和氮素的交互作用可以协调根、冠关系,只有当水分胁迫强度、历时和施氮量达到一定配合比例时,根、冠的生长才能达到一种最佳的平衡状态,为产量的提高提供物质基础。
4 结 语
(1)水分胁迫强度的增强和历时的增加可以促进根系干物质的积累,根冠比呈增大趋势,而且二者的交互作用明显高于单一因子对根系干物质累积的影响。增施氮肥更利于地上干物质的累积,根冠比减小,水、氮对根冠比的正负效应可以相互抵消,使根、冠均处于一种良好的平衡生长态势。
(2)水分胁迫强度、胁迫历时和氮素均是大豆产量产生补偿性的主要因子,三者的互作效应达到了显著水平,但符合报酬递减规律,高氮14 d重度水分胁迫使水肥的正效应达到最大值。
[1] 陈晓远,罗远培. 不同生育期复水对受旱冬小麦的补偿效应研究[J]. 中国农业生态学报, 2002,10(1):35-37.
[2] 闫永銮,郝卫平,梅旭荣,等. 拔节期水分胁迫-复水对冬小麦干物质积累和水分利用效率的影响[J]. 中国农业气象, 2011,32(2):190-195.
[3] 孟凡超,刘 明,于吉琳,等. 水分胁迫对玉米产量及干物质分配的影响[J]. 江苏农业科学, 2011,39(3):96-97,98.
[4] 张志伟,王志强,王 莹,等. 上部根区水分胁迫对玉米幼苗根系及地上部生长的影响[J]. 河南农业科学, 2013,42(5):29-56.
[5] Nowak M A, Boer list MC, Cooke J, et al. Evolution of genetic redundancy[J]. Nature, 1997,388(8):16-21.
[6] 蔡焕杰,康绍忠,张振华,等.作物调亏灌溉的适宜时间与调亏程度的研究[J]. 农业工程学报, 2000,16(3):24-27.
[7] 杨贵羽, 罗远培, 李保国,等. 水分胁迫持续时间对冬小麦根冠生物量累积的影响[J]. 干旱地区农业研究, 2006,24(4):94-98.
[8] 王林林,陈 炜,徐 莹,等. 氮素营养对小麦干物质积累与转运的影响[J].西北农业学报, 2013,22(10):85-89.
[9] 刘占军,谢佳贵,张 宽,等. 不同氮肥管理对吉林春玉米生长发育和养分吸收的影响[J]. 植物营养与肥料学报,2011,17(1):38-47.
[10] Watt M S, Clinton P W, Whitehead D, et al. Above-ground biomass accumulation and nitrogen fixation of broom(Cytisus scoparius L.) growing with juvenile Pinus radiata on adryland site [J]. Forest Ecology and Management, 2003,184(1-3):93-104.
[11] Zerihun A, Gutschick V P, Bassirirad H. Compensatory roles of nitrogen uptake and photosynthetic N-use efficiency in determining plant growth response to elevated CO2: evaluation using a functional balance model[J]. Annals of Botany, 2000,86:723-730.
[12] 葛体达,隋方功,李金政,等. 干旱对夏玉米根冠生长的影响[J].中国农学通报, 2005,21(1):103-109.
[13] Thornley J H M. Modeling shoot: root relations: the only way forwards[J]. Annals of Botany, 1998,81:165-171.
[14] 杨建昌,王志琴,朱庆森.不同土壤水分状况下氮素营养对水稻产量的影响及其生理机制的研究[J].中国农业科学,1996,29(4):58-66.
[15] 岳文俊,张富仓,李志军,等. 返青期水分胁迫、复水和施肥对冬小麦生长及产量的影响[J].西北农林科技大学学报(自然科学版), 2012,40(2):57-63.
[16] 孙 群,李学俊,达 娃.氮素对水分胁迫下玉米苗期生长和某些生理特性的影响[M]. 北京:中国农业科学技术出版社, 2001:128-130.
[17] 陈晓远, 高志红, 罗远培. 植物根冠关系[J]. 植物生理学通讯, 2005,41(5):555-562.
[18] Equiza M A, Mirave J P, Tognetti J A. Morphological, anatomical and physiological responses related to differential shootvs. root growth inhibition at low temperature in spring and winter wheat[J]. Annals of Botany, 2001,87:67-76.
猜你喜欢
中国科技纵横(2018年14期)2018-08-30
浙江农林大学学报(2018年4期)2018-08-22
现代农业科技(2017年16期)2017-09-22
江苏农业科学(2014年1期)2014-07-18
植物营养与肥料学报(2014年1期)2014-03-11
植物营养与肥料学报(2012年2期)2012-10-26
植物营养与肥料学报(2012年1期)2012-10-26
植物营养与肥料学报(2011年5期)2011-11-06
植物营养与肥料学报(2011年2期)2011-10-26
植物营养与肥料学报(2011年3期)2011-10-24
