
四川米仓山自然保护区台湾水青冈群落学特征及多样性研究
2016-03-23 02:29李大东董廷发史清茂何显湘章世鹏吴定军
西北植物学报 2016年1期
关键词:物种多样性
李大东,董廷发,陈 坚,史清茂,何显湘,章世鹏,吴定军,胥 晓*
(1 西南野生动植物资源保护教育部重点实验室,四川南充 637009;2 四川省旺苍县米仓山自然保护区管理局,四川旺苍 628200)
四川米仓山自然保护区台湾水青冈群落学特征及多样性研究
李大东1,董廷发1,陈坚2,史清茂2,何显湘2,章世鹏2,吴定军2,胥晓1*
(1 西南野生动植物资源保护教育部重点实验室,四川南充 637009;2 四川省旺苍县米仓山自然保护区管理局,四川旺苍 628200)
摘要:以四川米仓山自然保护区内的台湾水青冈(Fagus hayatae)群落为研究对象,采用20 m×30 m典型样地调查的方法,对群落的物种组成、区系特征、生活型谱、重要值、物种多样性、优势种群年龄结构及群落结构的相似性等方面进行分析。结果表明:(1)米仓山自然保护区内台湾水青冈群落中共有维管束植物129种,隶属于48科83属,其中蔷薇科为优势科;植物区系以北温带、泛热带及东亚和北美洲间断分布为主,具有南北区系的特点;生活谱型主要以高位芽植物(61.16%)和地下芽植物(23.14%)为主,其它生活型较少,这与米仓山台湾水青冈群落分布区所在的地理区位相吻合。(2)乔木层中台湾水青冈种群的重要值达到42.37%,为群落的单优势种。群落中灌木层的Shannon-Wiener指数为0.97,显著高于乔木层和草本层,反映出灌木层物种丰富的特点。(3)在台湾水青冈种群年龄结构中,Ⅰ级和Ⅱ级个体所占比例为24.05%,Ⅳ级和Ⅴ级个体所占比例为54.81%,反映出种群具有不良的自然更新特点。(4)群落相似性分析表明,台湾水青冈群落可分为4个类群,类群间在物种组成方面具有显著差异。研究认为,米仓山自然保护区内的台湾水青冈的群落特征和多样性水平因地处内陆已具有明显的地域特点。
关键词:台湾水青冈;群落学特征;物种多样性;米仓山自然保护区
台湾水青冈(Fagushayatae)属壳斗科(Fagaceae)水青冈属(Fagus)的落叶乔木,树高可达25 m以上,是中国特有树种也是国家二级重点保护植物[1]。该物种现主要分布于中国台湾岛的北部山区,在湖北、甘肃、四川和陕西的大巴山脉,以及浙江临安的清凉峰、永嘉四海山和庆元等地也有分布[2]。从现有资料来看,针对台湾水青冈的研究侧重点主要集中在种群更新动态与生境的关系、群落学特征、物种多样性、密度制约效应及ISSR-PCR体系的优化等方面,如郭瑞等对清凉峰台湾水青冈种群更新动态与其生境关系进行了研究[2];何俊等研究了七姊妹山和清凉峰等地台湾水青冈的群落学特征[3-4];翁东明等[5]研究了清凉峰台湾水青冈群落物种多样性;丁文勇等分别对清凉峰台湾水青冈群落优势种群的密度制约效应以及ISSR-PCR体系的优化进行了研究[6-7]。
由于近年来的全球气候变化、人类活动的长期干扰以及种群自身较弱的更新能力,台湾水青冈的生存和发展面临着种子散布困难[8-9],人为破坏严重[10],老树毁坏范围大[11]等困境;故深入了解台湾水青冈群落的特征,掌握其生物多样性状况,揭示其生存现状对该类植物的保护和发展显得尤为重要。四川米仓山国家级自然保护区位于米仓山-大巴山山脉西段南坡,地处北亚热带和北温带的交界区域,也是中国内陆地区首次发现台湾水青冈种群分布的区域所在地。目前国内外学者对台湾水青冈研究的区域主要集中在台湾拉拉山、浙江清凉峰、湖北七姊妹山等地[3,5-6,12],而针对内陆地区台湾水青冈群落的研究工作并不多见。米仓山山脉与台湾岛相距近1 700 km,中间又有台湾海峡相隔,二者在气候、土壤环境方面具有显著差异。因此推测分布于米仓山山脉的台湾水青冈群落特征和多样性水平可能具有明显的地域特点。本实验以四川米仓山自然保护区为研究区域,针对不同样地的台湾水青冈群落进行研究,研究结果为进一步了解和保护台湾水青冈种群提供理论参考。
1材料和方法
1.1研究区概况
米仓山自然保护区位于四川盆地北缘、米仓山南麓和旺苍县东北部,地理坐标为32°29′~32°41′N,106°24′~106°39′E,总面积23 400 hm2,该区域属亚热带湿润性季风气候,降雨充沛,湿度大。年均温13.5~16.5 ℃,年降水量达1 100~1 400 mm。土壤类型多样且呈垂直地带性分布,主要为黄壤(1 200 m以下)、黄棕壤(1 200~2 000 m)和山地棕壤(2 000 m以上)[13]。植被类型以亚热带常绿阔叶林为主,兼有落叶阔叶林分布。保护区内共有维管束植物195科949属2 597种,分别占全国维管束植物科属种的55.24%、29.86%和9.57%,根据陈坚[10]对米仓山自然保护区水青冈属资源的调查,结果表明该区域内广泛分布有水青冈(Faguslongipetiolata)、亮叶水青冈(F.lucida)、台湾水青冈和米心水青冈(F.engleriana)等4种水青冈属植物。
1.2研究方法
1.2.1样地设置和调查方法2014年7月对四川米仓山自然保护区内的台湾水青冈群落进行野外调查。鉴于保护区内台湾水青冈群落呈斑块状分布,故在每个斑块中选取代表性区域布设样地,每个样地与相邻样地至少相距2 km,样方大小为20 m×30 m,共设18个样方,记录样地的坡向、坡度、郁闭度、海拔及经纬度等信息(表1)。
采用常规群落调查方法,对样地内株高在5.0 m以上的乔木进行每木调查,记录下树种名称、胸径、树高、物候、枝下高及冠幅;并在每个样方的四角和中心分别设置1个5 m×5 m的灌木样方和1个1 m×1 m的草本样方。记录灌木和草本样方中的物种名称、高度、盖度、株丛数及基径。
1.2.2区系及生活型谱分析参照吴征镒[14]对中国种子植物属的分布区类型的划分标准,对样地内台湾水青冈群落中的所有属进行区系划分;根据Raunkiaer对植物生活型的划分方法[15],对保护区内的台湾水青冈群落的生活型谱进行统计分析。
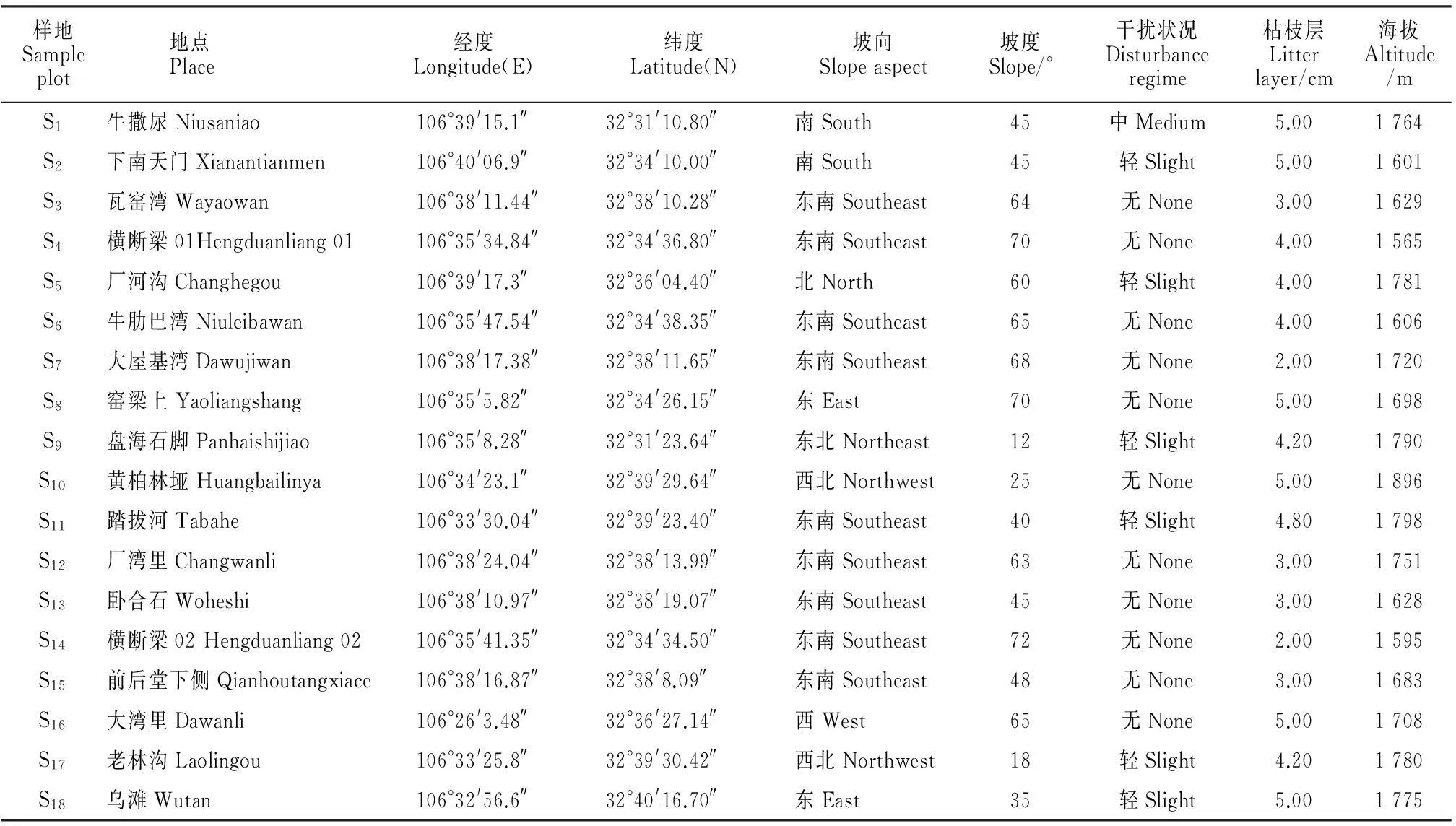
表1 四川米仓山自然保护区台湾水青冈群落样地信息表
1.2.3物种重要值和多样性分析乔木层物种的重要值参考马克平[16]及马克平和刘玉明[17]生物群落多样性的测度方法进行。公式如下:
乔木层重要值(IV)=[相对多度(RA)+相对显著度(RP)+相对频度(RF)]/3
物种多样性分析参考马克平的方法[18],分别选取了Shannon-Wiener指数(H)、Margalef丰富度指数(DM)、Pielou指数(J)、Simpson指数(DS)作为多样性的评价指标。公式如下:
Shannon-Wiener指数:H=-ΣPilnPi
Margalef丰富度指数:DM=(S-1)/lnN
Pielou指数:J=-ΣPilnPi/lnS
式中,S物种数目;N为所有物种个体数总和;Pi表示第i个物种的个体数与群落总个体数之比。
1.2.4种群年龄结构分析根据植物生长发育的情况并按照曲仲湘等[19]对立木的划分标准为依据,将样地内的台湾水青冈种群进行5级划分;Ⅰ级:高度<33 cm的幼苗;Ⅱ级:高度>33 cm胸径<2.5 cm的苗木;Ⅲ级:胸径在2.5~7.5 cm的幼树;Ⅳ级:胸径在7.5~22.5 cm者;Ⅴ级:胸径>22.5 cm者。并根据分级的结果采用Excel进行作图统计。
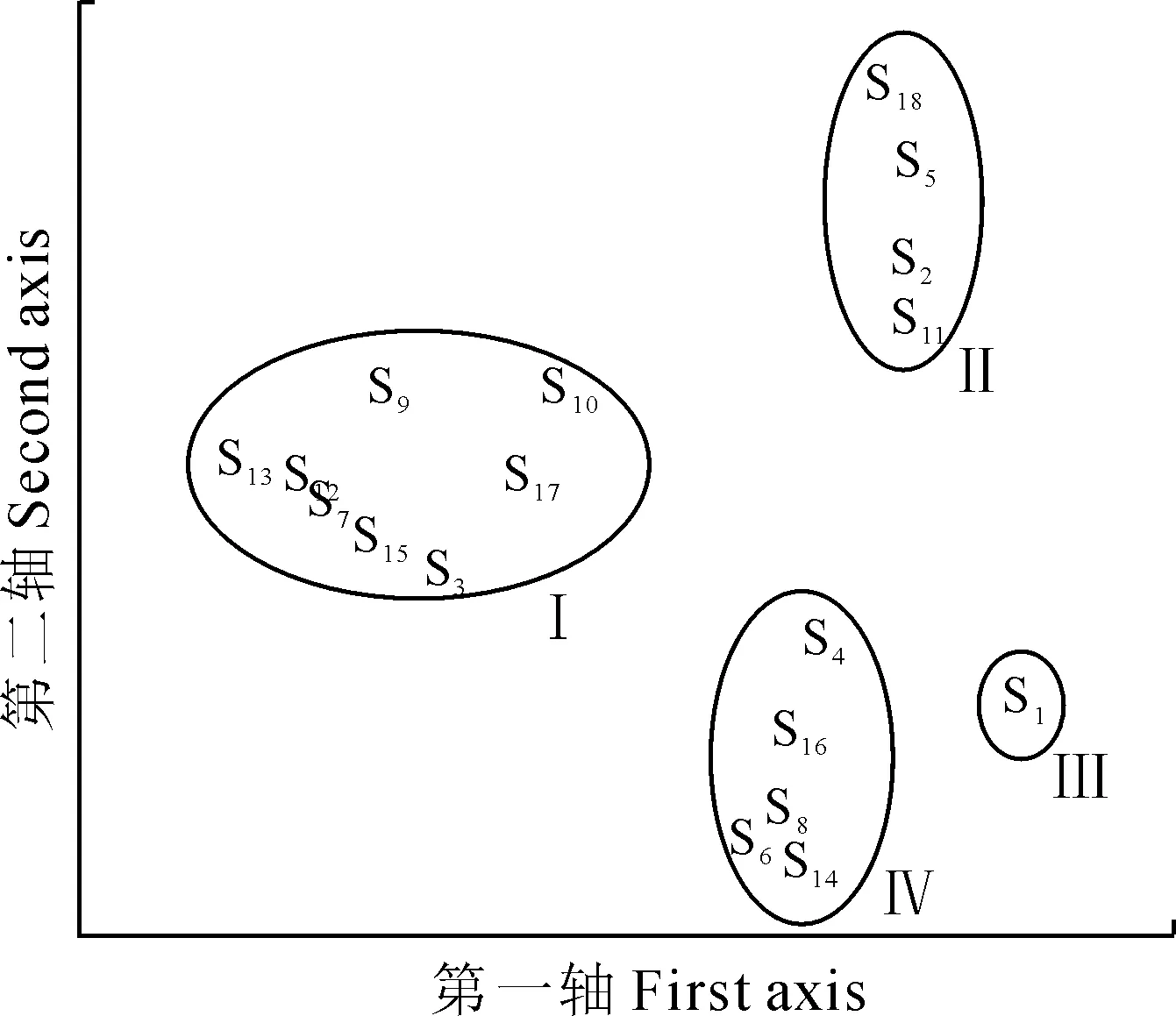
1.2.5群落结构的相似性分析运用英国普利茅斯海洋实验室开发的Primer 5.0软件进行群落结构的相似性分析。首先将所调查的18个台湾水青冈样地按照布设线路分为3个组;其次以18个样地为样本,各样地中的物种及株数为变量组成原始数据矩阵;再次采用Bray-Curtis Similarity系数及非加权的方法建立相似性矩阵并在此基础上运用组平均连接法建立等级聚类分析图;然后建立非度量多维标度排序(nonmetric multidimensional scaling,简称MDS)图分析米仓山自然保护区内台湾水青冈群落的结构特征[20-21]。最后用相似性分析和胁强系数来检验不同矩阵间的显著性差异,样本间的距离表示相似性的高低,相似性越高,距离越近。
2结果与分析
2.1群落的物种组成
样方调查结果显示,米仓山台湾水青冈群落中的植物共有129种,隶属48科83属(表2)。优势科是蔷薇科(Rosaceae,8属11种);其次是百合科(Liliaceae,7属10种)、壳斗科(Fagaceae,5属15种)和禾本科(Gramineae,5属6种);其中蕨类植物共有5科6属6种,占总种数的4.7%;裸子植物共有2科3属5种,占总种数的3.9%;被子植物共有41科74属118种,占总种数的91.5%。另有3个科超过10个种,它们分别为壳斗科15种,蔷薇科11种,百合科10种(表2)。

表2 四川米仓山自然保护区台湾水青冈
2.2植物区系组成
由表3可知,米仓山自然保护区内台湾水青冈群落由15个区系分布类型组成,主要以北温带、泛热带、东亚和北美洲间断分布及世界分布为主,分别占总属数的33.73%、18.07%、9.64%和9.64%。其中北温带分布类型有28属,主要位于群落的乔木层和草本层,如水青冈属、栎属(Quercus)、山茱萸属(Cornus)和绣线菊属(Spiraea)等;泛热带分布类型有15属,主要位于群落的灌木层和草本层,如卫矛属(Euonymus)、冬青属(Ilex)、紫金牛属(Ardisia)和菝葜属(Smilax)等;东亚和北美洲间断分布类型有8属,以灌木层和乔木层为主,如胡枝子属(Lespedeza)、十大功劳属(Mahonia)、石楠属(Photinia)和柯属(Lithocarpus)等;世界分布类型有8属,以草本层和灌木层为主,如灯心草属(Juncus)、堇菜属(Viola)、远志属(Polygala)和悬钩子属(Rubus)等。
2.3群落生活型谱
米仓山自然保护区内台湾水青冈群落的生活型谱(表4),在129种植物中,除去少量的藤本植物,种类最多的为高位芽植物,有74种,占总种数的61.16%;地下芽植物有28种,占总种数的23.14%;地上芽植物10种,占总种数的8.26%;种类较少的是地面芽(8种)和一年生植物(1种),分别占总种数的6.61%和0.83%。在高位芽植物中,大高位芽最少(6种);乔灌层物种丰富,小高位芽(31种),中高位芽(28种)植物较多,而矮高位芽数量较少(9种)。
2.4乔木层物种重要值
由表5和图1可见,在乔木层中台湾水青冈共有477株占据主导地位,具有较大的相对多度,径级结构主要以第Ⅴ级为主,其中Ⅳ(186株)、Ⅴ(190株)级的台湾水青冈占乔木层总株数的35.98%,相对显著度大,为68.13%,其重要值为42.37%;锐齿槲栎(Quercusalienavar.acuteserrataMaxim.)株数较多,相对多度较大,径级主要以第Ⅳ级为主,第Ⅴ级也具有一定比例,其重要值为7.70%位居第二;米心水青冈株树虽少于台湾水青冈但多于锐齿槲栎,其径级结构主要以第Ⅳ级为主,重要值为5.88%。其他伴生种如:猫儿刺(IlexpernyiFranch.)、四川杜鹃(RhododendronsutchuenenseFranch.)和柃木(EuryajaponicaThunb.)等重要值低不超过5.76%。
2.5物种多样性
物种多样性分析的结果表明(表6),在台湾水青冈群落中乔木层的物种数为38,灌木层为88,草本层为34;灌木层的Margalef指数(8.65)大于乔木层(5.32)和草本层(4.98);而在多样性指数和均匀度指数方面乔木层和草本层均大于灌木层。具体为:乔木层的Simpson指数(0.77)大于灌木层(0.33)和草本层(0.72);乔木层的Shannon-Wiener指数(2.27)大于灌木层(0.97)和草本层(1.98);乔木层的Pielou指数(0.62)大于灌木层(0.22)和草本层(0.56)。
2.6种群年龄结构
根据18个样地面积的统计结果表明,台湾水青冈种群处于Ⅰ级的个体有89株,Ⅱ级有76株,Ⅲ级有145株,Ⅳ级有186株,Ⅴ级有190株。Ⅳ、Ⅴ所占比例较大且接近,分别为27.11%和27.70%,Ⅲ级所占比例为21.14%,Ⅱ级所占比例最小,为11.08%,其次Ⅰ级所占比例为12.97%(图1)。

表3 四川米仓山自然保护区台湾水青冈群落维管束植物属的分布区类型
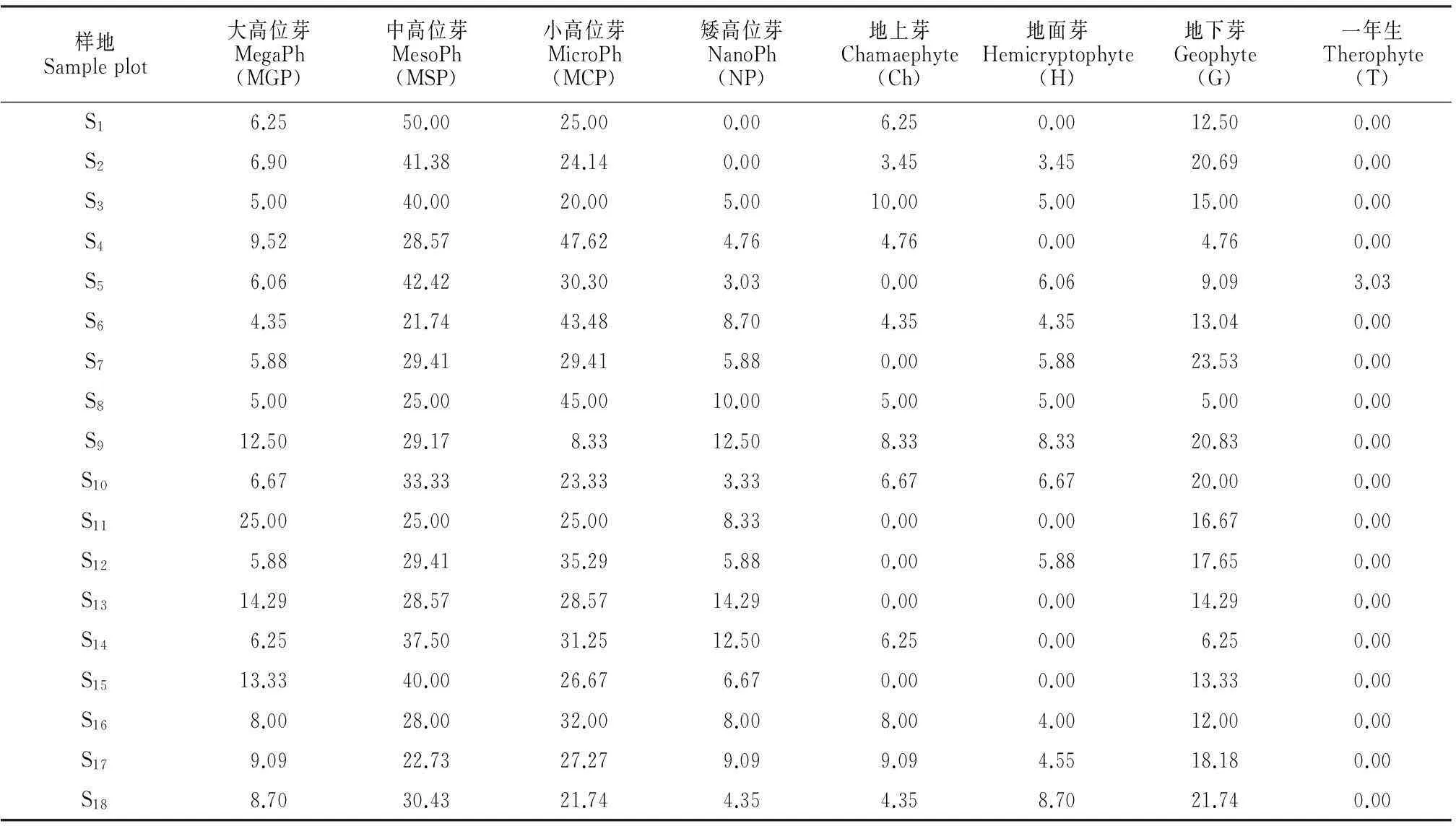
表4 四川米仓山自然保护区台湾水青冈群落植物生活型谱
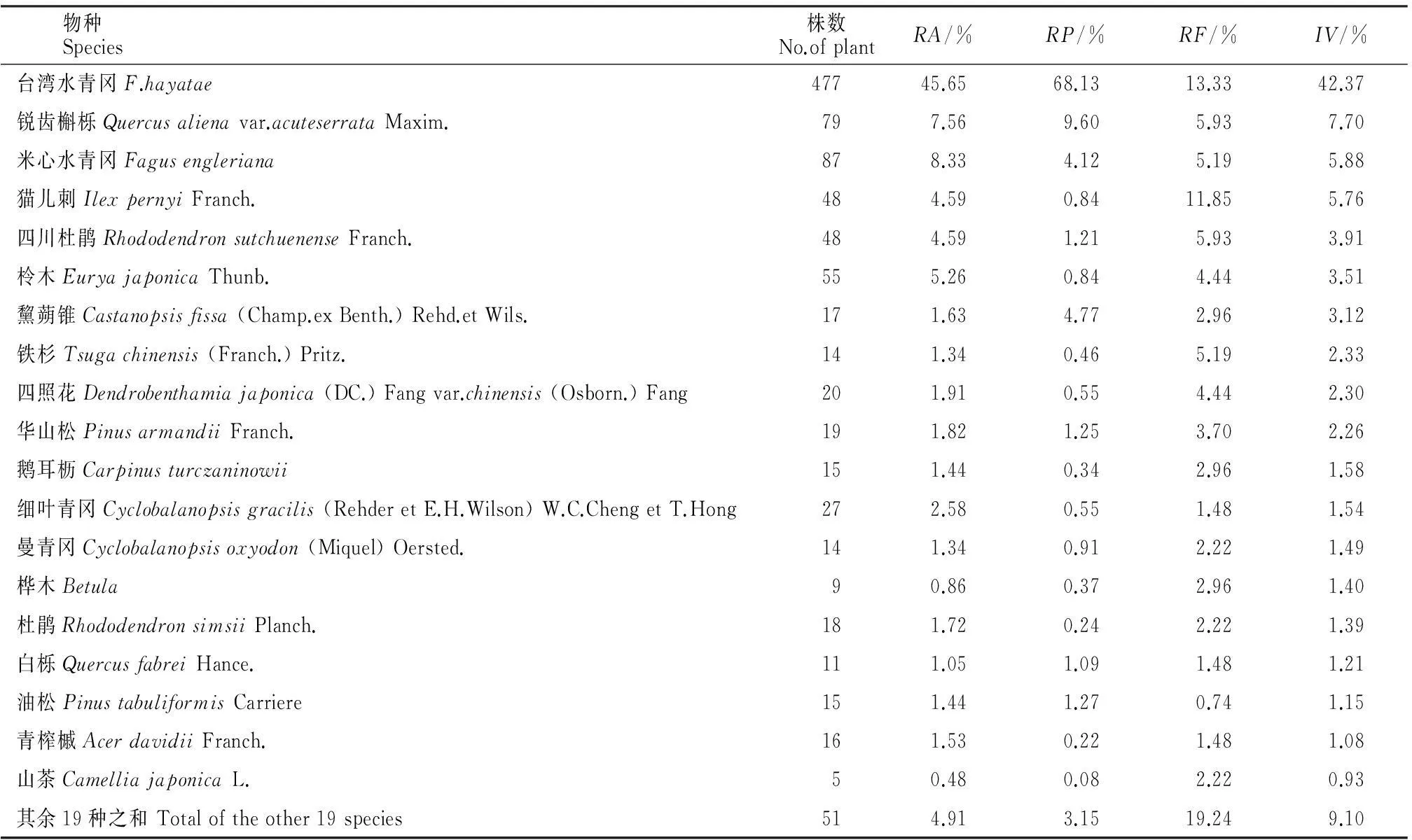
表5 四川米仓山自然保护区台湾水青冈群落乔木层物种重要值
注:RA.相对多度;RP.相对显著度;RF.相对频度;IV.重要值。
Notes:RA.Relative abundance;RP.Relative prominence;RF.Relative frequency;IV.Importance values.
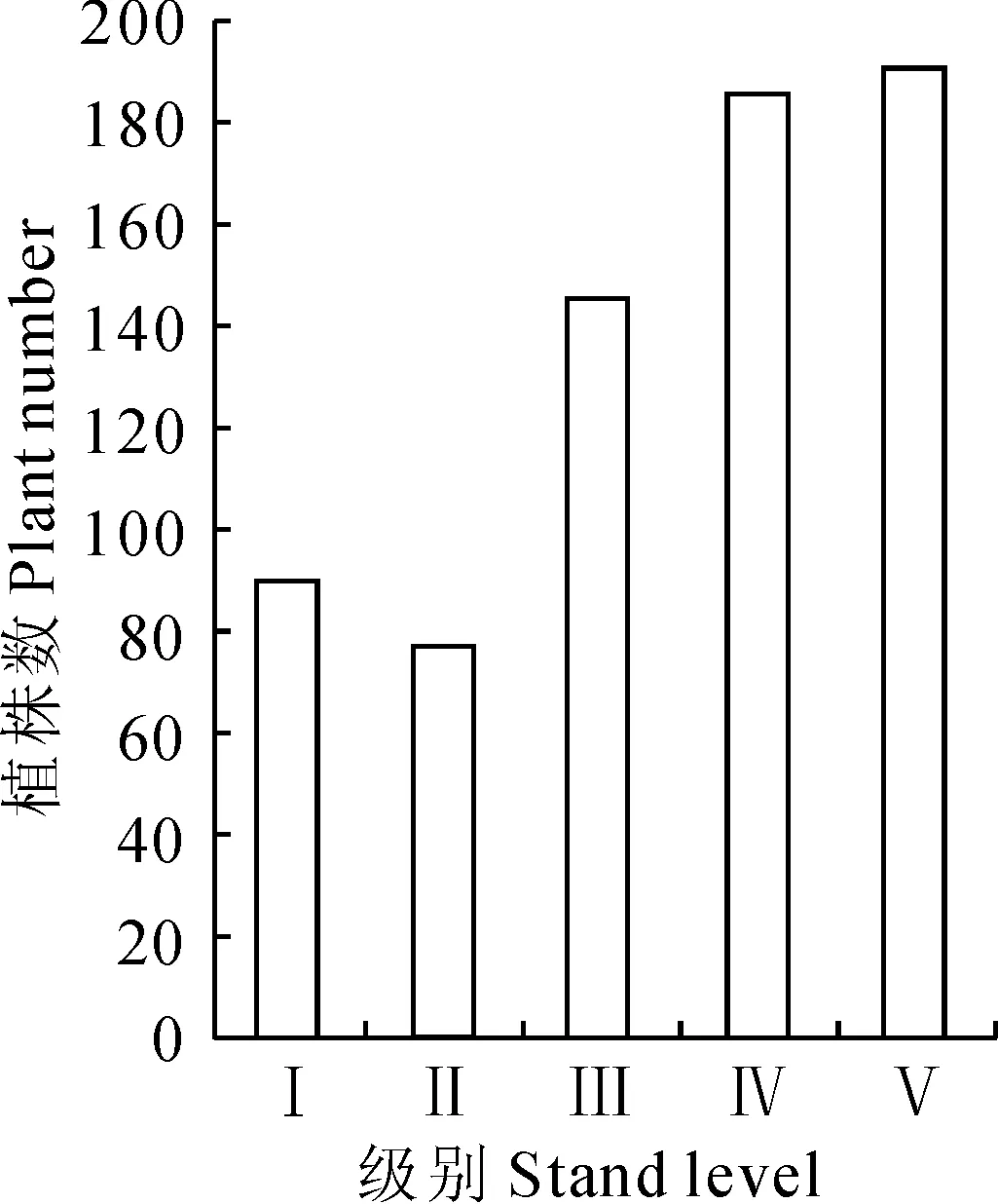
Ⅰ.高度<33 cm;Ⅱ.高度>33 cm、胸径(DBH)<2.5 cm;
2.7群落结构的相似性分析
根据Cluster聚类分析可知,在相似性系数为38%的水平上可将米仓山18个台湾水青冈群落的样地分为4个类群:类群I包括盘海石脚、瓦窑湾、前后堂下侧、卧合石、大屋基湾、厂湾里、黄柏林垭、老林沟;类群Ⅱ包括踏拔河、下南天门、厂河沟、乌滩;类群Ⅲ包括牛撒尿;类群Ⅳ包括横断梁01、横断梁02、大湾里、窑梁上、牛肋巴湾(图2和图3)。
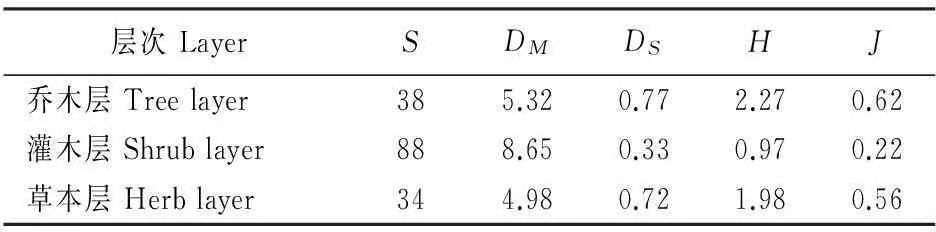
表6 四川米仓山自然保护区台湾水青冈群落物种多样性
注:S.总物种数;DM.Margalef指数;DS.Simpson指数;H.Shannon-Wiener指数;J.Pielou指数。
Notes:S.Number of species;DM.Margalef index;DS.Simpson index;H.Shannon-Wiener index;J.Pielou index.
3讨论
植物群落是生长在某一地区的植物经过长期历史发展的产物,因此植物群落的特征往往和所处的地理位置密切相关。米仓山自然保护区地处北温带与亚热带的过渡区,区内的植物群落在物种组成、区系特征和生活型谱等方面也很可能独具特点。我们发现台湾水青冈群落中共有维管束植物129种隶属48科83属;其中壳斗科、蔷薇科和百合科为优势科在群落中占据重要地位,且台湾水青冈群落区系中温带性质属(T)所占比例为60.24%,热带性质属(R)所占比例为28.92%。根据方全等[22]的研究,利用区系分析结果中温带属和热带属的比值(温热比)可以准确揭示植物区系的性质和特点。采用该方法,我们获得台湾水青冈群落中的温热比(T/R)为2.08,反映出该区温带属在整个植物区系中占主导地位。这与应俊生[23]对秦岭植物区系性质的研究结果基本一致。此外,我们还发现尽管台湾水青冈群落处于北温带和亚热带的过渡区,但其区系中缺失热带亚洲至热带大洋洲,温带亚洲,地中海区西亚至中亚和中亚及其变型等分布类型,一方面这可能是本研究只考虑了一种植物群落(台湾水青冈群落),而没有考虑其他群落所引起的统计误差。另一方面,也可能是因为台湾水青冈群落分布在海拔1 500~1 900 m的区域内,造成某些热带区系的植物分布受到限制,从而导致其区系成分的缺失。另外,我们还发现群落的生活型谱以高位芽植物为主,而地面芽和一年生植物相对较贫乏,其中高位芽尤以小高位芽和中高位芽占主导地位,大高位芽植物较少。根据江洪和于顺利等的研究[24-25],生活型谱的组成成分往往会受到群落所在地海拔、纬度或水热条件的影响。为此,对不同海拔段的高位芽植物比例进行了统计,结果发现随着海拔的增高其比例逐步下降,低海拔(1 565~1 629 m)样地的高位芽植物平均占比(80.73%)大于中海拔(1 683~1 764 m)样地(79.33%)和高海拔(1 775~1 896 m)样地(71.28%),表现出大高位芽植物的分布主要受海拔影响的特征,尽管该现象与前人对卧龙植物生活型垂直分布规律及对喀喇昆仑山-昆仑山地区植物生活型组成的研究结果相一致[26-27],但由于本研究样地海拔范围较小(1 565~1 896 m),且影响生活型谱的除了海拔还有纬度、地形、物种年龄等因子,所以该结果还有待进一步完善。另一方面,群落聚类分析可以直观地反映出群落间的相似性。采用Primer 5.0软件对米仓山自然保护区内的18个台湾水青冈群落进行聚类。结果表明,该区域的台湾水青冈群落可分为4个类群,且ANOSM和胁强系数的显著性检验均表明这4个类群间具有明显差异(P=2.0%,P<0.05;0.1 S1~S18代表样地编号,同表1;下同 图3 四川米仓山自然保护区台湾水青冈 种群年龄结构是种群的一个重要数量特征[32],可以反映出种群的更新状况。米仓山自然保护区台湾水青冈种群中幼苗和苗木等幼龄个体较少,大树和壮树等老年个体相对较多枯落层薄干扰程度轻,表明该区域内的台湾水青冈种群的自然更新不良,若一旦受到外界较大的干扰,种群可能出现较大的波动甚至其年龄结构也将受到影响。从野外调查的结果来看,台湾水青冈群落在自然演替中,其郁闭度增加,林内光照减少,导致台湾水青冈幼苗与邻近个体、灌木及下层乔木间对光环境的竞争加剧,幼苗的生长不良,死亡率增高。另一方面台湾水青冈本身自我更新能力相对较弱,扩散能力也很差所致[8-9]。因此,台湾水青冈自我的扩散能力较差和幼苗的种间竞争低下是引起其种群更新不良的主要原因。 通过与台湾拉拉山、浙江清凉峰及永嘉四海山等沿海地区的台湾水青冈群落相比[4-5,12],发现米仓山自然保护区台湾水青冈群落具有物种少(仅129种83属48科)、多样性低的特征,另一方面与永嘉四海山森林公园和清凉峰自然保护区台湾水青冈幼苗、幼树数量显著多于大树不同,米仓山自然保护区的台湾水青冈Ⅰ、Ⅱ级个体数量少(24.05%),Ⅳ、Ⅴ级个体数量大(54.81%),呈现出种群天然更新不良的特点;此外,在台湾水青冈群落分布高度方面,米仓山自然保护区与沿海地区差异也较大,浙江四海山和清凉峰台湾水青冈群落的分布海拔在650和1 000 m左右,而米仓山自然保护区台湾水青冈群落则主要分布于1 500~1 900 m。因此认为米仓山自然保护区内的台湾水青冈的群落特征和多样性水平因内陆其独特的气候和水热环境与沿海地区的台湾水青冈群落相比已具有明显的地域特点。 参考文献: [1]陈子英,谢长富,毛俊杰,等.冰河孑遗的夏绿林-台湾水青冈[M].台北:行政院农业委员会林务局,2011:1-26. [2]郭瑞,翁东明,金毅,等.浙江清凉峰台湾水青冈种群2006-2011年更新动态及其与生境的关系[J].广西植物,2014,34(4):478-483. GUO R,WENG D M,JIN Y,etal.Topography related regeneration dynamics ofFagushayataeduring 2006-2011 at Qingliangfeng Nature Reserve in Zhejiang Province[J].Guihaia,2014,34(4):478-483. [3]何俊,汪正祥,雷耘,等.七姊妹山自然保护区台湾水青冈林群落学特征研究[J].华中师范大学学报,2008,42(2):272-277. HE J,WANG Z X,LEI Y,etal.The study on coenological characteristics ofFagushayataecommunity in Qizimei Mountain Natural Reserve[J].JournalofHuazhongNormalUniversity,2008,42(2):272-277. [4]张方钢.浙江清凉峰台湾水青冈林的群落学特征[J].浙江大学学报,2001,27(4):403-406. ZHANG F G.The community characteristics ofFagushayataeforest of Qingliangfeng Mountain in Zhejiang[J].JournalofZhejiangUniversity,2001,27(4):403-406. [5]翁东明,张 磊,陈晓栋,等.清凉峰自然保护区台湾水青冈群落物种多样性研究[J].浙江林业科技,2009,29(4):1-6. WENG D M,ZHANG L,CHEN X D,etal.Species diversity ofFagushayataecommunity in Qingliangfeng National Nature Reserve[J].JournalofZhejiangForestryScienceandTechnology,2009,29(4):1-6. [6]丁文勇,翁东明,金毅,等.清凉峰自然保护区台湾水青冈群落优势种群密度制约效应分析[J].浙江大学学报,2014,41(5):583-592. DING W Y,WENG D M,JIN Y,etal.Density-dependent effects on dominant tree survival in anFagushayataecommunity located at Qingliangfeng National Nature Reserve[J].JournalofZhejiangUniversity,2014,41(5):583-592. [7]宋文静,金则新,李钧敏,等.台湾水青冈ISSR-PCR体系的优化[J].福建林业科技,2009,36(2):18-22. SONG W J,JIN Z X,LI J M,etal.Optimization of ISSR-PCR System ofFagushayatae[J].JournalofFujianForestryScienceandTechnology,2009,36(2):18-22. [8]肖宜安,何平,李晓红,等.濒危植物长柄双花木自然种群数量动态[J].植物生态学报,2004,28(2):252-257. XIAO Y A,HE P,LI X H,etal.Study on numeric dynamics of natural populations of the endangered speciesDisanthuscercidifoliusvar.longipes[J].ActaPhytoecologicaSinica,2004,28(2):252-257. [9]张志祥,刘鹏,蔡妙珍,等.九龙山珍稀濒危植物南方铁杉种群数量动态[J].植物生态学报,2008,32(5):1 146-1 156. ZHANG Z X,LIU P,CAI M Z,etal.Population quantitative characteristics and dynamics of rare and endangeredTsugatchekiangensisin Jiulongshan Nature Reserve of China[J].JournalofPlantEcology,2008,32(5):1 146-1 156. [10]陈坚.米仓山自然保护区水青冈属(Fagus)资源调查报告[J].中国野生植物资源,2014,33(2):47-52. CHEN J.Report of investigation onFagusof Micangshan Nature Reserve[J].ChineseWildPlantResources,2014,33(2):47-52. [11]POCS T.The epiphytic biomass and its effect on the water balance of two rain forest types in the Uluguru Mountains[J].ActaBotanicaAcademiaeScientiarumHungaricae,1980,26:143-167. [12]王献溥,李俊清.广西水青冈林的分类研究[J].植物研究,1996,16(4):369-404. WANG X P,LI J Q.The study of beech forest classification in Guangxi[J].BulletinofBotanicalResearch,1996,16(4):369-404. [13]何毓蓉.米仓山林区土壤的肥力特征及保护研究[J].水土保持学报,1991,5(4):73-79. HE Y R.Study on protective fertility and properties of soil fertility in forest region of the Micangshan[J].JournalofSoilandWaterConservation,1991,5(4):73-79. [14]吴征镒.中国种子植物属的分布区类型[J].云南植物研究,1991,(S):1-139. WU Z Y.The areal-types of Chinese genera of seed plants[J].ActaBotanicaYunnanica,1991,(S):1-139. [15]曲仲湘.植物生态学[M].北京:高等教育出版社,1990:142-147. [16]马克平.生物群落多样性的测度方法——α多样性的测度方法(上)[J].生物多样性,1994,2(3):162-168. [17]马克平,刘玉明.生物群落多样性的测度方法-α多样性的测度方法(下)[J].生物多样性,1994,2(4):231-239. [18]马克平.生物群落多样性测度方法[M]//钱迎倩,马克平.生物多样性的原理与方法.北京:中国科学技术出版社,1994:141-165. [19]曲仲湘,文振旺.琅琊山林木现况的分析[J].植物学报,1953,2(3):349-370. QU Z X,WEN Z W.An ecological observation on the Langyashan woods,Anhui Province[J].JournalofIntegrativePlantBiology,1953,2(3):349-370. [20]周红,张志南.大型多元统计软件PRIMER的方法原理及其在底栖群落生态学中的应用[J].青岛海洋大学学报,2003,33(1):58-64. ZHOU H,ZHANG Z N.Rationale of the multivariate statistical software PRIMER and its application in benthic community ecology[J].JournalofOceanUniversityofQingdao,2003,33(1):58-64. [21]杨少荣,高欣,马宝珊,等.三峡库区木洞江段鱼类群落结构的季节变化[J].应用与环境生物学报,2010,16(4):555-560. YANG S R,GAO X,MA B S,etal.Seasonal dynamics of fish community in Mudong section of the three gorges reservoir of the Yangtze River,China[J].ChineseJournalofAppliedandEnvironmentalBiology,2010,16(4):555-560. [22]方全,刘以珍,林朝晖,等.云居山栓皮栎群落特征及多样性研究[J].植物科学学报,2015,33(3):311-319. FANG Q,LIU Y Z,LIN Z H,etal.Research onQuercusvariabiliscommunity characteristics and diversity of Yunju Mountain[J].PlantScienceJournal,2015,33(3):311-319. [23]应俊生.秦岭植物区系的性质,特点和起源[J].植物分类学报,1994,32(5):389-410. YING J S.An analysis of the flora of Qinling Mountain range:its nature,characteristics,and origins[J].ActaPhytotaxonomicaSinica,1994,32(5):389-410. [24]江洪.东灵山植物群落生活型谱的比较研究[J].植物学报,1994,36(11):884-894. JIANG H.Study on life-form spectrum of plant community in Dongling Mountain[J].ActaBotanicaSinica,1994,36(11):884-894. [25]于顺利,陈灵芝,马克平.东北地区蒙古栎群落生活型谱比较[J].林业科学,2000,36(3):118-121. YU S L,CHEN L Z,MA K P.Life-form spectrum of vascular plants ofQuercusmongolicacommunities in North East China[J].ScientiaSilvaeSinicae,2000,36(3):118-121. [26]蔡绪慎,黄加福.卧龙植物生活型垂直分布规律初探[J].西南林学院学报,1990,10(1):31-40. CAI X S,HUANG J F.An approach to the vertical distribution pattern of plant life forms in Wolong Natural Reserve[J].JournalofSouthwestForestryCollege,1990,10(1):31-40. [27]郭柯,郑度,李渤生.喀喇昆仑山-昆仑山地区植物的生活型组成[J].植物生态学报,1998,22(1):51-59. GUO K,ZHENG D,LI B S.The characteristics of plant life form spectra in the Karakorum-Kunlun Mountains[J].ActaPhytoecologicaSinica,1998,22(1):51-59. [28]CLARKE K R.Non-parametric multivariate analyses of changes in community structure[J].AustralianJournalofEcology,1993,18:117-143. [29]CLARKE K R,GORLEY R N.Primer 5.0:user manual/tutorial.Plymouth,UK:Primer-E,2001. [30]张峰,张金屯,张峰.历山自然保护区猪尾沟森林群落植被格局及环境解释[J].生态学报,2003,23(3):421-427. ZHANG F,ZHANG J T,ZHANG F.Pattern of forest vegetation and its environmental interpretation in Zhuweigou,Lishan Mountain Nature Reserve[J].ActaEcologicaSinica,2003,23(3):421-427. [31]张文江,宁吉才,宋克超,等.岷江上游植被覆被对水热条件的响应[J].山地学报,2013,31(3):280-286. ZHANG W J,NING J C,SONG K C,etal.The response of vegetation cover to the variation of heat and water conditions in upper Minjiang watershed,China [J].JournalofMountainScience,2013,31(3):280-286. [32]冷平生.园林生态学[M].北京:中国农业出版社,2003:155-156. (编辑:潘新社) Characteristics ofFagushayataeCommunity and Species Diversity in Micangshan Nature Reserve,Sichuan LI Dadong1,DONG Tingfa1,CHEN Jian2,SHI Qingmao2,HE Xianxiang2,ZHANG Shipeng2,WU Dingjun2,XU Xiao1* (1 Key Laboratory of Southwest China Wildlife Resources Conservation,Ministry of Education,Nanchong,Sichuan 637009,China;2 Micangshan Nature Reserve,Wangcang,Sichuan 628200,China) Abstract:The community characteristics of Fagus hayatae and species diversity in Micangshan Nature Reserve (32°29′-32°41′N,106°24′-106°39′E) of Sichuan were studied on the 20 m×30 m forest plots.The results indicated that:(1)there were 129 vascular plant species belonging to 83 genera and 48 families in the F.hayatae community and Rosaceae was the dominant family.The north temperate,pantropic and east Asia-north America disjuncted distribution were the dominant floristic distribution types,which had the characteristics of the north and south.The main life-form spectrum distribution was Phanerophytes (61.16%) and Geophyta (23.14%),with relatively fewer other life-form spectrum,which was consistent with the geographieal location of F.hayatae community distributed in Micangshan Mountain.(2)F.hayatae was the single dominant species,and its important value was 42.37%.The Shannon-Wiener index of the shrub layer in the community was 0.97,significantly higher than that of tree layer and herb layer,which reflected the abundance of species in shrub layer.(3)In age structure of F.hayatae population,the individual of Ⅰ and Ⅱ occupying 24.05%,Ⅳ and Ⅴ accounting for 54.81%,which showed the poor natural regeneration in the population.(4)The community structure similarity analyses showed that F.hayatae communities could be divided into 4 groups with significant differences in species composition.The results showed that the characteristics of F.hayatae community and species diversity in Micangshan Nature Reserve have obvious regional features due to the inland. Key words:Fagus hayatae;community characteristics;species diversity;Micangshan Nature Reserve 中图分类号:Q948.15+6 文献标志码:A 作者简介:李大东(1989-),男,硕士,主要从事植物生态研究。E-mail:2240126200@qq.com*通信作者:胥晓,博士,教授,硕士生导师,主要从事植物生理生态研究。E-mail:xuxiao_cwnu@163.com 基金项目:四川米仓山自然保护区管理局委托项目(2014-02) 收稿日期:2015-0-0;修改稿收到日期:2015-12-23 文章编号:1000-4025(2016)01-0174-09 doi:10.7606/j.issn.1000-4025.2016.01.0174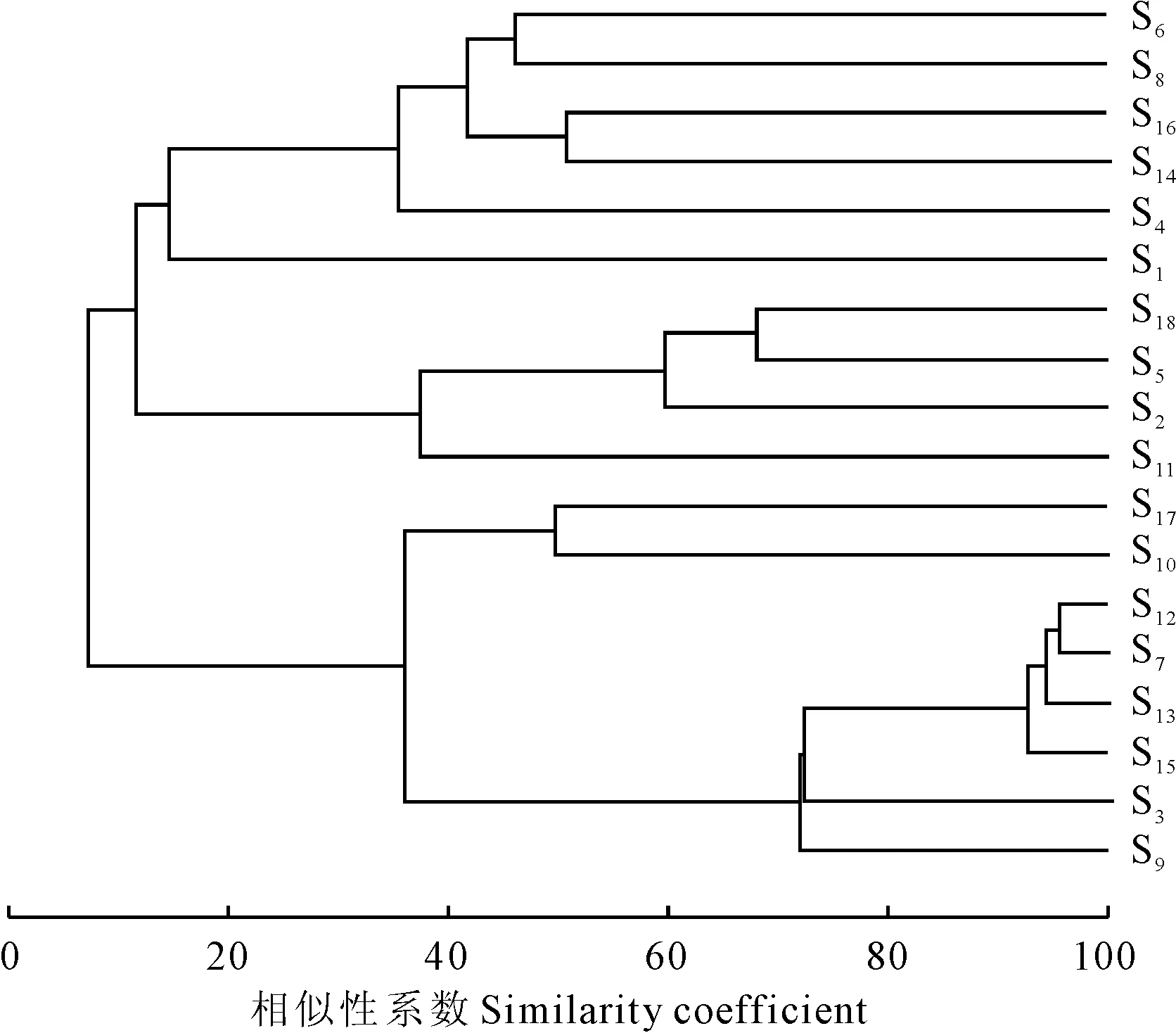
猜你喜欢
湖北农业科学(2016年21期)2017-03-18
山东工业技术(2017年5期)2017-03-16
山东农业科学(2017年2期)2017-03-15
吉林农业·下半月(2017年2期)2017-03-10
绿色科技(2016年21期)2016-12-27
现代农业科技(2016年20期)2016-12-20
科技创新导报(2016年21期)2016-12-17
现代经济信息(2016年13期)2016-06-17
绿色科技(2015年6期)2015-08-05
江苏农业科学(2015年1期)2015-04-17
