
A new species of the genus Scutiger (Anura: Megophryidae) from Medog of southeastern Tibet, China
2016-03-22 03:35KeJIANGKaiWANGDaHuZOUFangYANPiPengLIJingCHEStateKeyLaboratoryofGeneticResourcesandEvolutionKunmingInstituteofZoologyChineseAcademyofSciencesKunmingYunnan650ChinaSamNobleOklahomaMuseumofNaturalHistoryandDepartmento
Zoological Research 2016年1期
Ke JIANG, Kai WANG, Da-Hu ZOU, Fang YAN, Pi-Peng LI, Jing CHE, *State Key Laboratory of Genetic Resources and Evolution, Kunming Institute of Zoology, Chinese Academy of Sciences, Kunming Yunnan 650, ChinaSam Noble Oklahoma Museum of Natural History and Department of Biology, University of Oklahoma, Norman OK 707-709, U.S.A.Tibet University, Lhasa Tibet 850000, ChinaInstitute of Herpetology, Shenyang Normal University, Shenyang Liaoning 00, China
A new species of the genus Scutiger (Anura: Megophryidae) from Medog of southeastern Tibet, China
Ke JIANG1,#, Kai WANG1,2,#, Da-Hu ZOU3,1, Fang YAN1, Pi-Peng LI4, Jing CHE1, *1State Key Laboratory of Genetic Resources and Evolution, Kunming Institute of Zoology, Chinese Academy of Sciences, Kunming Yunnan 650223, China
2Sam Noble Oklahoma Museum of Natural History and Department of Biology, University of Oklahoma, Norman OK 73072-7029, U.S.A.3Tibet University, Lhasa Tibet 850000, China
4Institute of Herpetology, Shenyang Normal University, Shenyang Liaoning 110034, China
Received: 17 October 2015; Accepted: 20 December 2015
Foundation items: This study was supported by the "Light of West China" Program of the Chinese Academy of sciences (CAS), the Ministry of Science and Technology of China (2014FY210200, 2011FY120200), and the Animal Branch of the Germplasm Bank of Wild Species of Chinese Academy of Sciences (the Large Research Infrastructure Funding)
#Authors contributed equally to this work
ABSTRACT
A new species of Scutiger Theobald, 1868 is described from Medog, southeastern Tibet, China, based on morphological and molecular data. The new species was previously identified as Scutiger nyingchiensis, but it can be differentiated from the latter and all other congeners by the following combination of characters: (1) medium adult body size, SVL 50.5-55.6 mm in males and 53.8-57.2 mm in females; (2) maxillary teeth absent; (3) web rudimentary between toes; (4) prominent, conical-shaped tubercles on dorsal and lateral surfaces of body and limbs; (5) tubercles covered by black spines in both sexes in breeding condition; (6) a pair of pectoral glands and a pair of axillary glands present and covered by black spines in males in breeding condition, width of axillary gland less than 50% of pectoral gland; (7) nuptial spines present on dorsal surface of first and second fingers, and inner side of third finger in males in breeding condition; (8) spines absent on the abdominal region; (9) vocal sac absent. In addition, the distribution and conservation status of the new species are also discussed. Keywords: Eastern Himalayas; Tibet; Scutiger nyingchiensis; Scutiger spinosus sp. nov.; DNA barcoding
INTRODUCTION
The Tibet-Hengduan Mountain area is one of the 35 biodiversity hotspots of the world (Mittermeier, 2004) and supports an abundance of organisms, many of which are endemic to the region. Among the endemic fauna, the majority of species in the megophryid frog genus Scutiger Theobald, 1868 are known only from the montane habitats at high elevations between 1 900 m and 5 100 m in this region. Currently, there are 20 species recognized in the genus (Frost, 2015), of which six species are known from Tibet (Jiang et al., 2012), including S. boulengeri (Bedriaga, 1898), S. maculatus (Liu, 1950), S. mammatus (Güther, 1896), S. nyingchiensis Fei, 1977, S. wuguanfui Jiang et al., 2012, and S. sikkimensis (Blyth, 1854). During our fieldwork from 2011 to 2013 in southeastern Tibet, we collected 16 specimens of megophryid frogs that were initially identified as Scutiger nyingchiensis from 62K, Medog (=Motuo). After morphological comparisons and genetic analysis with respect to topotype specimens of S. nyingchiensis, we were able to distinguish however, the Scutiger specimens from 62K, Medog can be readily distinguished from S. nyingchiensis by a suit of morphological characters and a significant genetic distance. Therefore, we describe the 62K population of Scutiger as a new species.
MATERIALS AND METHODS1
From June 2011 to June 2013, a total of 16 individuals (12 males and four females) were collected from 62K, Medog, southeastern Tibet, China. Following euthanasia, liver tissues were taken from each individual and preserved in 95% ethanol, and all specimens were then fixed in 10% formalin solution and transferred to 75% ethanol after returning from the field. All specimens were designated as the type series, andwere deposited in Kunming Institute of Zoology, Chinese Academy of Sciences.
Morphological analysis
Morphological characters used and their measurement methods followed Fei et al. (2009). Measurements were taken with dial calipers and recorded to the nearest 0.1 mm.
The morphological characters and their abbreviations as: SVL, snout-vent length; HL, head length; HW, head width; SL, snout length; INS, internarial distance; IOS, interorbital distance; EHD, eye horizontal diameter; UEW, maximum width of upper eyelid; FAHL, forearm and hand length; FAW, maximum width of forearm; HAL, hand length; FML, femur (thigh) length; TBL, tibia (shank) length; TFL, length of tarsus and foot; FOL, foot length.
The new species was compared to museum specimens (Appendix) as well as descriptions in the literature: Scutiger adungenis (Dubois, 1979), S. bhutanensis (Delorme & Dubois, 2001), S. brevipes (Liu, 1950), S. nepalensis (Dubois, 1973), and S. chintingensis, S. glandulatus, S. gongshanensis, S. jiulongensis, S. liupanensis, S. maculatus, S. muliensis, S. ningshanensis, S. pingwuensis, S. tuberculatus and S. wanglangensis from Fei et al. (2009). The following museum abbreviations were used: CIB-Chengdu Institute of Biology, Chinese Academy of Sciences, Chengdu, China. KIZ-Kunming Institute of Zoology, Chinese Academy of Sciences, Kunming, China.
Molecular analysis
Total DNA was extracted from 15 specimens of seven Tibetan Scutiger species (Table 1), using a standard three-step phenolchloroform method (Sambrook et al., 1989). A 561 base pair DNA fragment of the mitochondrial gene Cytochrome Oxidase Subunit I (COI) was amplified and sequenced for all samples using primers Chmf4 and Chmr4 (Che et al., 2012). Protocols for PCR and sequencing followed Che et al. (2012). All novel sequences were deposited in GenBank (KU243053-KU243067, Table 1). Sequences for three additional species of Tibetan Scutiger were downloaded from Genbank as well outgroup taxa including Xenophrys, Brachytarsophrys and Leptolalax (Table 1).
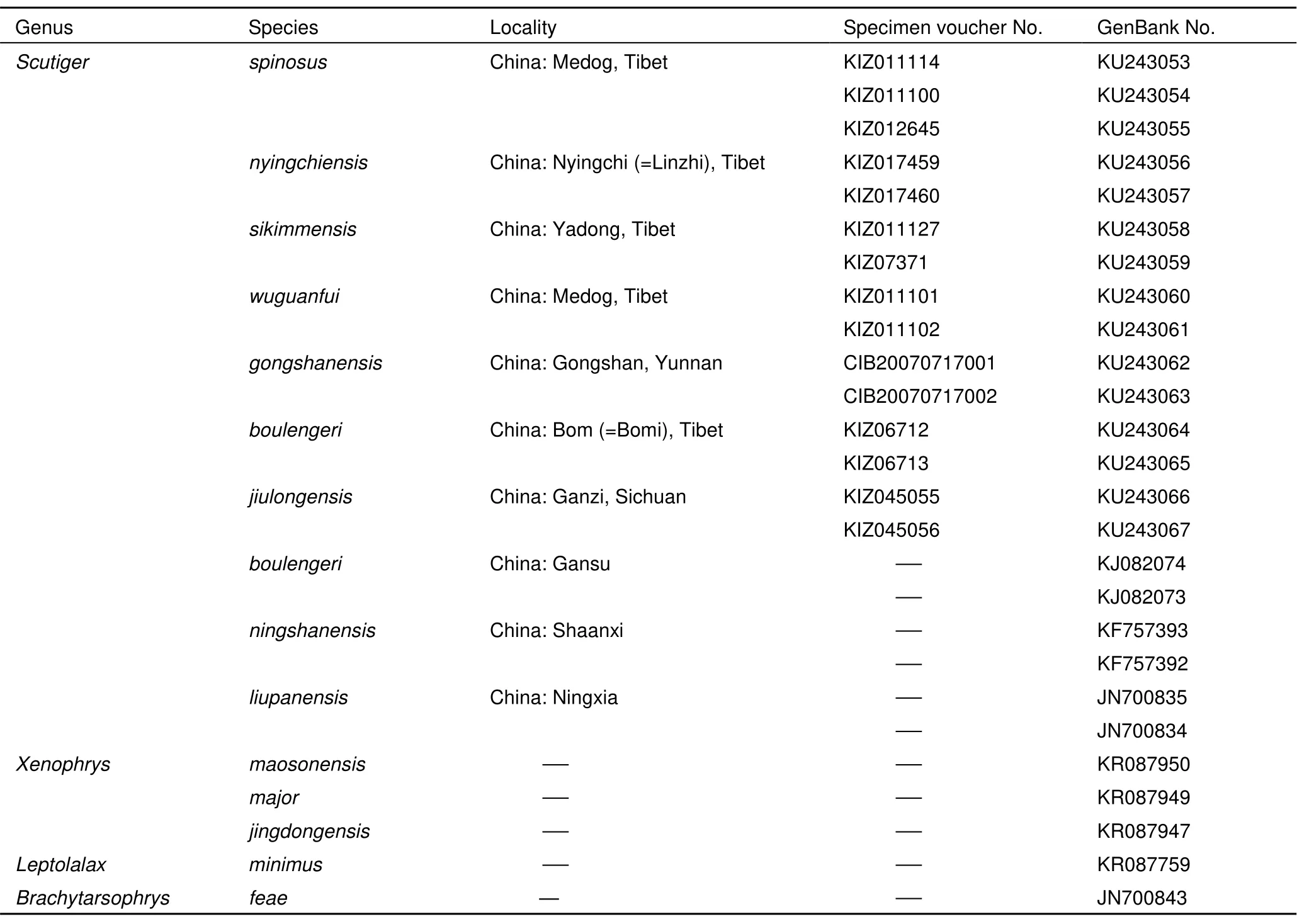
Table 1 Information of samples used in molecular analysis
All sequences were aligned and edited in MEGA 5 (Tamura et al., 2011). The best model of nucleotide substitution for each codon position was SYM+I, F81 and GTR+G determined using jModeltest v1.0.1 (Posada, 1998). Bayesian inference (BI) was used to generate a phylogenetic relationship using MrBayes 3.1.2 (Ronquist & Huelsenbeck, 2003). Using different modelfor each codon position, the Markov chains were estimated for 10 million generations, and every 100 generations were sampled. The genetic distance between species was calculated using MEGA 5 with Kimura 2-parameter model (Che et al., 2012).
RESULTS
Morphological comparison
The results of the morphological comparisons between the Scutiger population from 62K, Medog and its congeners are summarized in Table 2. The morphological comparisons reveal that the Scutiger population at 62K, Medog can be readily distinguished from morphologically similar S. nyingchiensis and other congeners by a suite of diagnostic characters, including a light brown triangular pattern of pigmentation on the dorsal surface of the head, rudimentary webbing between toes, and prominent and conical shaped tubercles on the dorsal and lateral surfaces of the body and limbs.
Phylogenetic relationships
The 561 base pairs (bp) of COI sequences for 15 individuals were sequenced (Table 1). There are 155 and 202 potentially parsimony informative sites within genus Scutiger and between the ingroup and outgroup, respectively. The BI analysis supports the genus Scutiger as a monophyletic group. Five lineages are identified based on available data for Scituger (Clade A-E, Figure 1). Species distributed in Tibet are recovered in Clade A, C, D, and E. The 62K population of Scutiger, along with S nyingchiensis from Nyingchi, Tibet and S. gongshanensis from Gongshan, Yunnan, forms the linage D. The 62K population of Scutiger differs from the phenotypically similar S. nyingchiensis and from S. gongshanensis by a genetic distance of 10.6% and 10.9% respectively. Scutiger nyingchiensis and S. gongshanensis are sister species, with a genetic distance of 8.5%.
Furthermore, the populations of S. boulengeri from Gansu and Tibet do not form a monophyletic lineage, and the genetic distance between the two populations is 8.1%.
Given that both morphological and molecular results support the Scutiger population from 62K, Medog as a distinct evolutionary lineage distinct from all other Tibetan Scutiger, therefore, following the species concept by and concordant evidence confirm species status (Wu & Murphy, 2015), herein we describe the 62K population of Scutiger as a new species.
Scutiger spinosus sp. nov. Jiang, Wang, Li and Che (Figures 2-4)
Synonyms
Scutiger nynchiensis: Li et al., 2010; Jiang et al., 2012
Holotype: KIZ 011114 (Figure 2), an adult male from 62K, Medog, Tibet, China (N29°42′33.6″, E95°34′56.0″, elevation 2 705 m). Collected by Ke JIANG on 07 June 2011.
Paratypes: a total of 15 specimens (11 males and four females) from the same locality as holotype, including one adult male (KIZ 011113) and two adult females (KIZ 011093, KIZ 011100), collected by Ke JIANG and Pi-Peng LI as same date as holotype; three adult males (KIZ 012645, 012647-48) and one adult female (KIZ 012646) collected by Ke JIANG, Kai WANG and Jiang XIE on 15 July 2012; and seven adult males (KIZ 013862-64, 013866-69) and one adult female (KIZ 013865), collected by Ke JIANG and Kai WANG on 17 June 2013.
Diagnosis: Scutiger spinosus sp. nov. is identified to the genus Scutiger by its molecular phylogenetic position and the following morphological characters: (1) maxillary teeth absent or indistinct; (2) vomerine teeth absent; (3) tympanum and tympanic ring entirely absent; (4) pupil vertical, elliptical; (5) pectoral and axillary gland present in males, and covered by black spines in breeding condition; (6) inner three fingers of males with black nuptial spines in breeding condition.
Scutiger spinosus sp. nov. is distinguished from all other congeners by the following combination of morphological characters: (1) medium adult body size, SVL 50.5-55.6 mm in males and 53.8-57.2 mm in females; (2) maxillary teeth absent; (3) web rudimentary between toes; (4) prominent, conical-shaped tubercles on dorsal and lateral surfaces of body and limbs; (5) tubercles covered by black spines in both sexes in breeding condition; (6) a pair of pectoral glands and a pair of axillary glands present and covered by black spines in males in breeding condition, width of axillary gland less than 50% of pectoral gland; (7) nuptial spines on dorsal surface of first and second fingers, and inner side of third finger in males in breeding condition; (8) spines absent on the abdominal region; and (9) vocal sac absent.
Description of the holotype: SVL 51.7 mm; body slightly compressed, moderately slender at waist; head large and flat, width nearly equal to length (HW/HL=1.04); snout rounded, slightly projecting beyond jaw; canthus rostralis obtuse, loreal region oblique and slightly concave; nostril midway between the tip of snout and eye; eye relatively large (EHD/HL=0.35); pupil vertical; interorbital space flat, interorbital distance less than upper eyelid width (IOS/UEW=0.90); tympanum and tympanic rim absent; a small tooth-like projection on anteromedial edge of mandible; jaws without teeth; tongue oval, slightly emarginate behind, papillae absent; choanae located against anterior border of palate, visible when viewed from below; vomerine teeth absent; vocal sac absent; supratympanic fold from posterior corner of orbit to shoulder, distinctly developed.
Forelimbs long; forearm enlarged; fingers slender, free of web or dermal fringes; relative length of fingers: I=II<IV<III; tips of fingers rounded, not dilated; subarticular tubercles absent; inner metacarpal tubercle almost equal to outer metacarpal tubercle, both large and flat; nuptial spines on dorsal surface of first and second fingers, and on inner side of third finger.
Hindlimbs moderately short, tibiotarsal articulation reaching the corner of mouth when adpressed, heels do not touch when folded at right angles to the body (FML/TBL=1.07); foot longer than shank (TFL/TBL=1.26); tips of toes round; dermal fringesfeeble; relative length of toes: I<II<III<V<IV; subarticular tubercles absent; ridges on undersurfaces of toes absent; inner metatarsal tubercle elliptical and prominent, outer metatarsal tubercle absent.
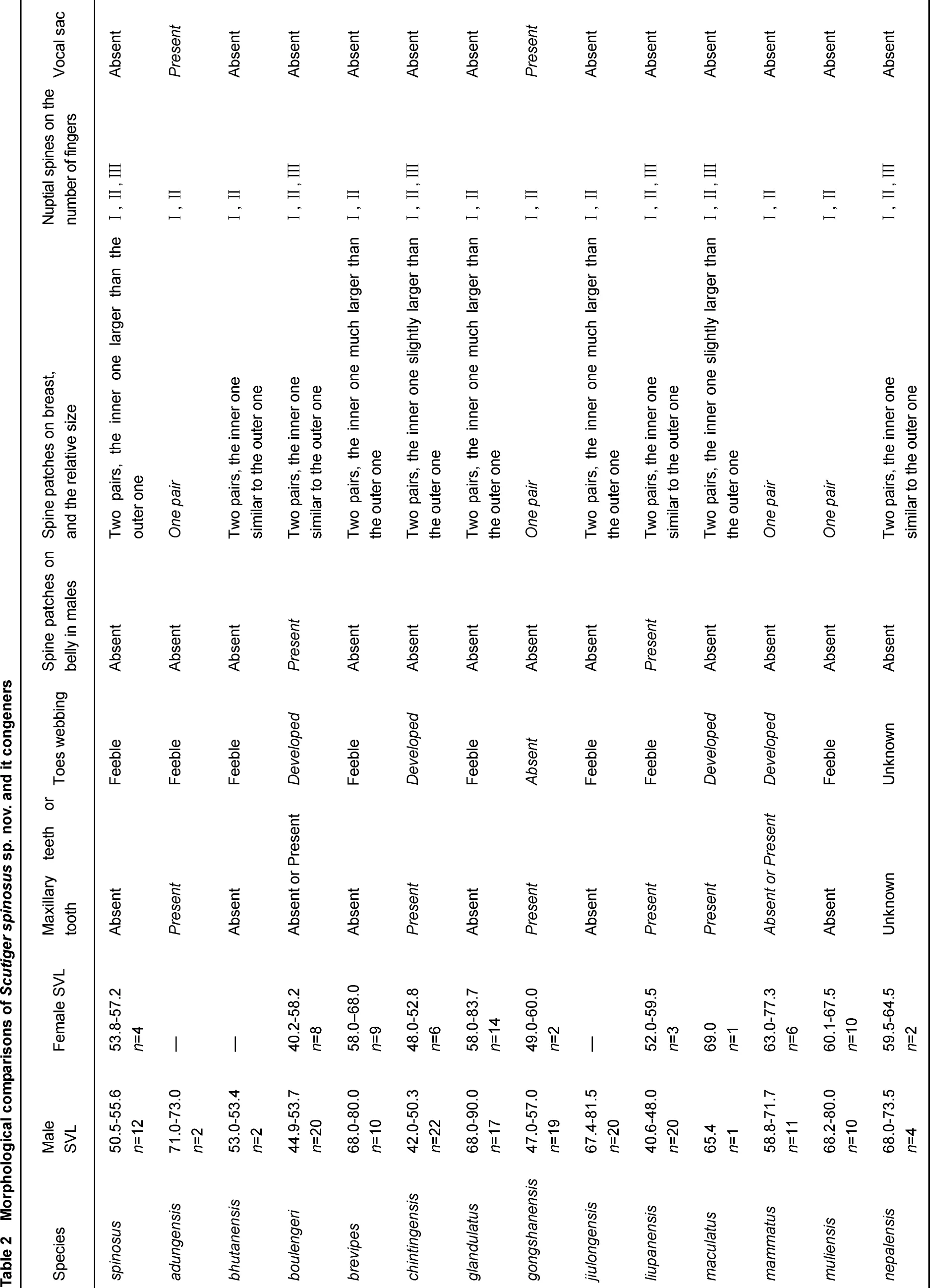


Figure 1 Bayesian inference tree based on barcoding COI data of the Tibetan congeners of the genus Scutiger

Figure 2 Scutiger spinosus sp. nov. holotype (KIZ 011114) in preservative (Photos by Ke JIANG)
Skin of dorsal side extremely rough. Forehead and upper lip relatively smooth, scatted small tubercles without spine; small tubercles present below supratympanic fold, each tubercle bearing one black spine; supratympanic fold, top of head, dorsal sides of body and limbs, and flanks covered with large, prominent, conical-shaped tubercles, all bearing black spines except on dorsal side of forearm; small tubercles scatted among the larger tubercles, mostly bearing black spine. Skin on ventral surface of body mostly smooth; numerous small black spines present on margin of lower jaw, small patches of black spines present near armpit. A pair of pectoral glands and a pair of axillary glands present on chest, pectoral twice longer than axillary, both covered by dense black spines.

Figure 3 Scutiger spinosus sp. nov. paratype (KIZ 013867), adult male in life (Photos by Kai WANG)

Figure 4 Scutiger spinosus sp. nov. paratype (KIZ 013865) adult female in life (Photos by Kai WANG)
Coloration in life: A light brown, triangular pattern of pig-mentation is observed on the dorsal surface of the anterior portion of the head, the apex is at the tip of snout and the base is between eyes. Several dark spots overlay the triangular pattern. Lateral surfaces of the head are dark brown; upper lip is creamy white, with irregular shaped brown mottling. The coloration of the remaining dorsal surfaces of the head, body, lower forelimbs, and hind limbs are dark brownish gray. The flanks are light brown and gradually fade into light brownish yellow ventrally. Dorsal surfaces of the upper forelimbs are light brownishyellow, with few transverse stripes of darker brown; dorsal surfaces of the lower forelimbs are dark gray, with few light gray transverse bands. Light mottling patterns of pigmentations are also observed on the dorsal surfaces of the hind limbs. Ventral surfaces of the head, body, and limbs are uniform light yellow.
The coloration in preservative closely resembles the coloration in life, except that the off-white coloration of the lips and the light brownish yellow of the venter fade and become light gray.
Variation: Measurements of type series are shown in Table 3. Individuals of type series are generally uniform in external characters, except for the characters related to sexual dimorphism in the males.
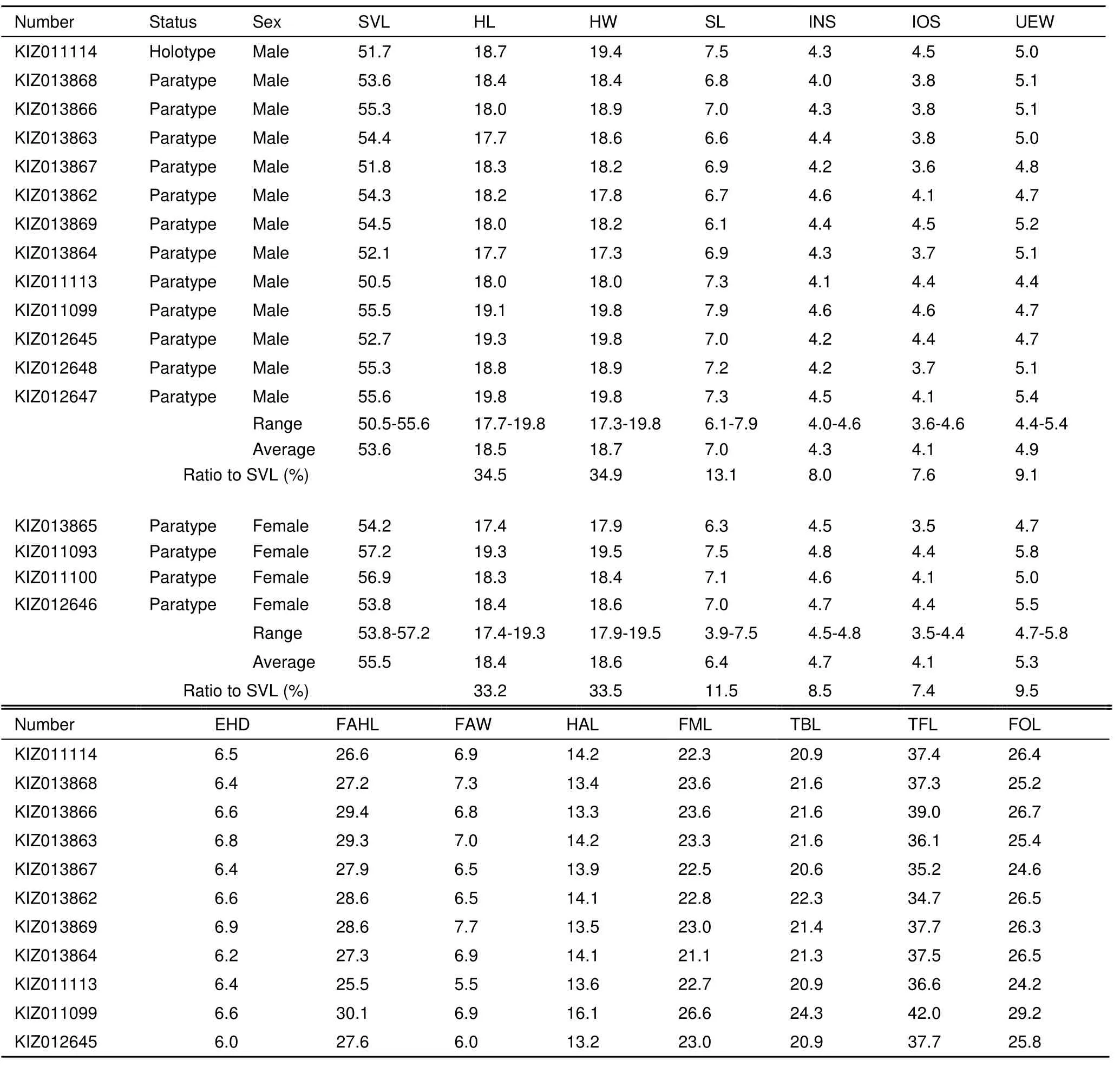
Table 3 Morphological measurements (mm) of the type series of Scutiger spinosus sp. nov.
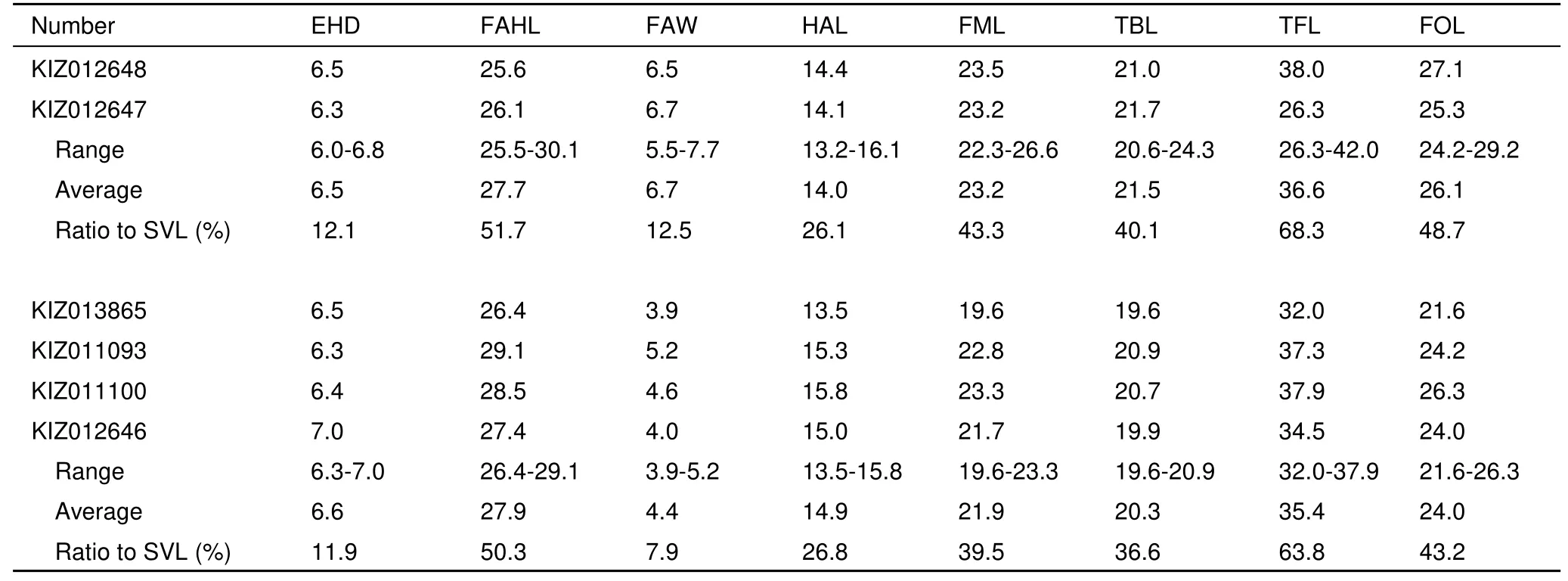
Continued
Sexual dimorphism: In males, small patches of black spines are present on the armpit, and a pair of pectoral glands and a pair of axillary glands are present on the chest, with the former ones much larger than the latter ones. Both pectoral and axillary glands are covered by black spines in breeding condition. Nuptial black spines are present on the dorsal surfaces of the first and second finger and the inner side of the third finger with in breeding condition. Forearms of males are much stronger than females (FAW male/female=1.5). In breeding condition, spines on the dorsal and lateral surfaces of males are more distinct and denser than females. Vocal sac and lineae masculinae are absent in males.
Ecological notes: The new species inhabits the mixed forest at high elevation, sympatric with Scutiger wuguanfui, Nanorana medogensis (Fei and Ye, 1999), and Liurana xizangensis (Hu, 1977). Breeding pairs were found under logs during the day and in the streams and ponds at night from the beginning to the end of June. The breeding habitats include slow-flowing streams and small to medium permanent ponds, with brownish, slightly acidic water. Males display inguinal amplexus, where the males grasp the females at the waist (Figure 5). Eggs are laid in donut-shaped masses and are attached to the under surfaces of logs and rocks (Figure 6). There is no evidence of nuptial spines on the fingers and pectoral and axillary glands in mid July. Tadpoles overwinter and take more than one year to develop into metamorphic juveniles, which is a common adaptation for high-elevation amphibians.

Figure 5 Breeding pair of the Scutiger spinosus sp. nov., showing the inguinal amplexus (Photos by Kai WANG)

Figure 6 Breeding habitat (A) and eggs (B) of Scutiger spinosus sp. nov. at 62K of Medog, southeastern Tibet, China (Photos by Kai WANG and Pi-Peng LI)
Etymology: The specific epithet spinosus, is derived from the Latin word spina, meaning spine and combined with ōsus to form an adjective indicating an abundance of the noun, spina. Spinosus is in reference to the conical-shaped tubercles and the numerous spines on the dorsal surfaces of this species.Based on the Latin name we suggest the English common name to be “Spiny Lazy Toad”, the Chinese name as “Ci You Chi Tu Chan” (刺疣齿突蟾).
Morphological Comparisons: Scutiger spinosus sp. nov. is morphologically most similar to S. nyingchiensis and S. boulengeri, in which both species have similar body size, brownish gray dorsal coloration, and distinct tubercles on the dorsal surface of the body. However, the new species can be distinguished readily from S. nyingchiensis (Figure 7) by having rudimentary webbings on toes (v.s. well developed), prominent, conical-shaped tubercles on the dorsal and lateral surfaces of the body and limbs (v.s. tubercles elongated, relatively low, and not conical in shape), as well as by the presence of a distinct, light brown, triangular pattern on the dorsal surface of the head (v.s. absent), presence of large tubercles on the dorsal surfaces of thigh (v.s. absent), presence of spines on the upper arm (v.s. absent), presence of numerous, spine-bearing, small tubercles scatted among the large tubercles on the dorsal and lateral surfaces of the body and limbs (v.s. absent), and the presence of spines on the tubercles on the dorsal surface of the body and limbs in females (v.s. absent); and from S. boulengeri by having rudimentary webbings on toes (v.s. well developed), prominent, conical shaped, spine-bearing tubercles on the dorsal and lateral surfaces of the body (v.s. relatively flat, not conical shaped, with smaller spines), and by the absence of spines on the abdominal region in breeding condition in males (v.s. present).

Figure 7 Male and female of Scutiger nyingchiensis (Photos by Da-Hu ZOU, Bao-Lin ZHANG and Ke JIANG).
Scutiger spinosus sp. nov. is closely related to S. gongshanensis based on the molecular phylogeny, but the new species differs from the latter by the absence of vocal sacs in males (v.s. presence), presence of axillary glands (v.s. absent) and presence of black spines on dorsal tubercles (v.s. absent). Additionally, the new species differs from the sympatric S. wuguanfui, by having a distinct dorsal coloration (brownish gray v.s. dark purplish brown), a smaller adult body size (SVL 50.5-55.6 mm in males, 53.8-57.2 mm in females v.s. 77.5-83.8 mm in males, 107.4-116.7 mm in females), prominent, conical, spine-bearing tubercles on the dorsal and lateral surfaces of body and limbs (v.s. flat protuberances that are relatively smooth), as well as by the absence of vocal sacs in males (v.s. presence).
DISCUSSION
Our molecular data shows that all known Scutiger species form a monophyletic group (Figure 1). Surprisingly, despite the closer geographic proximity and similar external morphology between S. nyingchiensis and S. spinosus, S. nyingchiensis is more distantly related to S. spinosus than S. gongshanensis. Scutiger gongshanesis is distributed much farther away in the Hengduan Mountain Range on the east and has quite different external morphology. Additionally, Scutiger boulengeri is paraphyletic based on our molecular phylogeny, which might be the results of mitochondrial gene introgression with other sympatric Scutiger species (Chen et al., 2009), existence of cryptic species, or mis-identification of congeners. Further phylogenetic and phylogeographic studies are needed to gain a better understanding of this interesting relationship.
Recently, Sarania et al. (2015) reported S. nyingchiensis from the China-India Disputed Region based on photographic evidence without vouchered specimens. However, based on the morphological characters described and shown in their photos, such as the rudimentary webbings on toes, the presence of spines on tubercles in females, and an inverted triangular mark of pigmentation from the upper eyelid to the snout, these individuals possess diagnostic characters of the new species, and thus are likely to be S. spinosus instead of S. nyingchiensis. Further examinations of vouchered specimens from the region are needed to ensure the taxonomic status of this population.
Although the new species is locally abundant, its pristine habitats are currently under great threats from rapid developments of roads in southeastern Tibet. Several breeding pools of the new species were eliminated entirely during the sampling period of our study, and significant deforestations were observed along the roads. Because the new species and other amphibians from the same habitats, namely S. wuguanfui, Nanorana medogensis and Liurana xizangensis are believed to be restricted to the mixed forests in this region, such deforestation and destructions of breeding habitat could be devastating to the long-term survival of these endemic amphibians. We recommend immediate attention to the conservation and protection of habitats in this area including the type locality of the new species. Given the recent human incroachment into these areas, in-depth assessments of the ecology and populationstatus of the endemic amphibians including the new species Scutiger spinosus are needed now more than ever.
ACKNOWLEDGEMENTS
We thank to Mr. Bao-Lin ZHANG (KIZ) and Mr. Jiang XIE (North China Electric Power University Science and Technology College), and the volunteers, Mr. Tao LIANG, Mr. Duan YOU, Mr. Ya-Di HUANG and Mr. Ya-Qiang SUN, who helped with fieldwork in Tibet, Dr. Yun-Ke WU (Cornell University) and Dr. Yu ZENG (University of California, Berkeley) for providing some literatures.
REFERENCES
Che J, Chen HM, Yang JX, Jin JQ, Jiang K, Yang ZY, Murphy RW, Zhang YP. 2012. Universal COI primers for DNA barcoding amphibians. Molecular Ecology Resources, 12 (2): 247–258.
Chen W, Bi K, Fu JZ. 2009. Frequent mitochondrial gene introgression among high elevation Tibetan megophryid frogs revealed by conflicting gene genealogies. Molecular Ecology, 18 (13): 2856–2876.
Delorme M, Dubois A. 2001. Une nouvelle espèce de Scutiger du Bhutan, et quelques remarques sur la classification subgénérique du genre Scutiger (Megophryidae, Leptobrachiinae). Alytes, 19: 141–153.
Dubois A. 1974 “1973”.. Diagnoses de trois espèce nouvelles d’amphibiens du Népal. Bulletin de la Société Zoologique de France, 98: 495–497.
Dubois A. 1979. Une espèce nouvelle de Scutiger (Amphibiens, Anoures) du nord de la Birmanie. Revue Suisse de Zoologie, 86: 631–640.
Fei L, Hu SQ, Ye CY, Huang YZ. 2009. Fauna Sinica, Amphibia, Vol. 2. Beijing: Science Press, 1–957. (in Chinese)
Frost DR. 2015. Amphibian Species of the World: an Online Reference. Version 6.0 (Nov 12, 2015). Electronic Database accessible at http: //
research.amnh.org/herpetology/amphibia/index.html. American Museum of Natural History, New York, USA.
Jiang K, Rao DQ, Yuan SQ, Wang JS, Li PP, Hou M, Chen MH, Che J. 2012. A new species of the genus Scutiger (Anura: Megophryidae) from southeastern Tibet, China. Zootaxa, 3388: 29–40.
Li PP, Zhao EM, Dong BJ. 2010. Amphibians and Reptiles of Tibet. Beijing: Science Press, 1–251. (in Chinese)
Liu CC. 1950. Amphibians of Western China. Chicago Natural History Museum, 2: 1–397.
Mittermeier RA, Gil PR, Hoffmann M, Pilgrim JD, Brooks TB, Mittermeier CG, Lamoreux JL, Fonseca GAB. 2004. Hotspots Revisited: Earth’s Biologically Richest and Most Endangered Ecoregions. Mexico City: CEMEX, 1–390.
Posada D, Crandall KA. 1998. MODELTEST: testing the model of DNA substitution. Bioinformatics, 14 (9): 817–818.
Ronquist F, Huelsenbeck JP. 2003. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics, 19 (12): 1572–1574.
Sambrook J, Fritsch E, Maniatis T. 1989. Molecular Cloning: A Laboratory Manual (2nd edition). New York: Cold Spring Harbor Laboratory Press, .
Sarania B, Devi A, Kumar A, Wang K, Rakshit K. 2015. A record of Scutiger nyingchiensis Fei, 1977 (Amphibia: Anuran: Megophryidae) in the Eastern Himalaya, North East India. Current Science, 109 (3): 413–414.
Tamura K, Peterson D, Peterson N, Stecher G. 2011. MEGA5: molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Molecular Biology and Evolution, 28 (10): 2731–2739.
Wu YK, Murphy RW. 2015. Concordant species delimitation from multiple independent evidence: A case study with the Pachytriton brevipes complex (Caudata: Salamandridae). Molecular Phylogenetic and Evolution, 92: 108–117.
APPENDIX
The following specimens were examined:
Scutiger boulengeri (n=28): KIZ 012573, KIZ 012581, KIZ 012583-84 (4♂♂), Nyalam (=Nielamu), Tibet; KIZ 012605 (1♂), KIZ 012602-04 (3♀♀), Tingri (=Dingri), Tibet; KIZ 014935-39, KIZ 014941-42, KIZ 014944-46 (9♂♂), KIZ 014948, KIZ 014950 (2♀♀), Qamdo (=Changdu), Tibet; KIZ 012606-07, KIZ 012609-11, KIZ 012614 (6♂♂), KIZ 012616-18 (3♀♀), Ndong (=Naidong), Tibet.
Scutiger glandulatus (n=7, unmeasured): KIZ 016477-80, KIZ 016482 (5♂♂), KIZ 016481, KIZ 016483 (2♀♀), Ganzi, Sichuan.
Scutiger mammatus (n=17): KIZ 016469-70, KIZ 017293-95, KIZ 017447-48, KIZ 017454-56, KIZ 017458 (11♂♂), KIZ 016471, KIZ 017288-89, KIZ 017449, KIZ 017457 (5♀♀), Jomda (=Jiangda), Tibet; KIZ 014035 (1♀), Baxoi (=Basu), Tibet.
Scutiger nyingchiensis (n=6): KIZ 017459-60 (2♂♂, Nyingchi); KIZ 09416, KIZ 019415-16 (3♂♂), KIZ 016217 (1♀), Gongbo'gyamda (=Gongbu’jiangda), Tibet. Scutiger sikimmensis (n=10): KIZ 011127, KIZ 013983-88, KIZ 013990 (8♂♂), KIZ 013982, KIZ 013989 (2♀♀), Yadong, Tibet.
Scutiger wuguanfui (n=8): KIZ 01101–02, KIZ 011109-11, KIZ 012649 (6♂♂), KIZ 011116, KIZ 012650 (2♀♀), Medog, Tibet.
DOI:10.13918/j.issn.2095-8137.2016.1.21
*Corresponding author, E-mail: chej@mail.kiz.ac.cn

- Zoological Research的其它文章
- New Year address from Zoological Research
- Advances in herpetological research emanating from China
- The Australasian frog family Ceratobatrachidae in China, Myanmar and Thailand: discovery of a new Himalayan forest frog clade
- A new genus and species of treefrog from Medog, southeastern Tibet, China (Anura, Rhacophoridae)
- A new species of the genus Amolops (Amphibia: Ranidae) from southeastern Tibet, China
- Two new species of Japalura (Squamata: Agamidae) from the Hengduan Mountain Range, China