
空间信息技术在河口潮间带栖息地分类研究中的应用
2016-03-04 08:31:56张婷婷侯俊利
海洋渔业 2016年4期
张婷婷,赵 峰,侯俊利,高 宇,庄 平
(中国水产科学研究院东海水产研究所,农业部东海与远洋渔业资源开发利用重点实验室,农业部东海与长江口渔业资源环境科学观测实验站,上海 200090)
·综述·
空间信息技术在河口潮间带栖息地分类研究中的应用
张婷婷,赵 峰,侯俊利,高 宇,庄 平
(中国水产科学研究院东海水产研究所,农业部东海与远洋渔业资源开发利用重点实验室,农业部东海与长江口渔业资源环境科学观测实验站,上海 200090)
研究河口潮间带底栖生境(intertidal benthic habitat)变化,能为河口湿地生物的保护、底栖环境质量的评价、受损生境的生态恢复提供的科学依据和管理建议。本文从关注空间和时间尺度的研究出发,对河口潮间带栖息地监测、分类与评价方面的研究进展进行了归纳总结,分为以下六个方面进行论述:河口潮间带底栖生境的时空新特征、河口潮间带底栖生境特征参数、河口潮间带底栖生境特征获取方式、数据处理、河口潮间带底栖生境分类模型选择、多时空尺度分析。河口潮间带底栖生境特征的分类方式由于考虑生态过程和格局中所涉及的空间生态学问题(边界分割、尺度效应、分类精度、动态变化等),模拟底栖物种及群落分布更为便捷,最终使物种/群落栖息地分类结果更加精确、真实、全面。最后,本文提出利用SIT对河口潮间带底栖生境进行分类将对底栖生态系统评价及研究生态功能机制带来新的视角,同时也需注意一些问题。例如,潮间带生境特征参数需要选择合适的统计方法在不同的尺度上进行甄别和筛选,以期有效提高指示参数在研究尺度上的解释度。
空间信息技术;河口潮间带;底栖生境;分类研究
潮间带栖息地(intertidal habitat)是河口生态系统的重要组成部分,其与陆地、海洋生态系统的营养物质交换频繁、环境复杂多变,具有支持植被、提供动物栖息所,维持生物多样性的生态功能[1]。在全球气候变化、上游水沙变化和外来物种入侵等自然因素以及促淤圈围、深水航道工程、水库建设等人为因素的共同作用下,高度敏感、脆弱的河口潮间带栖息地是受污染物和人类活动影响最早和最直接的生态区,近几十年来发生了巨大变化,其生态系统的功能和服务遭到严重破坏[2]。作为与河口潮间带理化环境密切相关的底栖动物群落[3],其分布能有效反映河口生态系统的健康状况[4],是指示河口潮间带生境变化最直接的指示因子。研究和监测河口潮间带底栖生境(intertidal benthic habitat)变化,能为河口湿地生物的保护、底栖环境质量的评价、受损生境的生态恢复提供科学依据和管理建议。
随着空间信息技术(spatial information technologies,SIT)的发展,SIT技术逐渐囊括地理信息系统技术(geographic information system,GIS)、遥感(remote sensing,RS)、空间模型(spatial modeling)、空间统计学(spatial statistics)[5],利用SIT技术能获取栖息地的空间信息及动态的时间信息,具有多样性与多维性,使科学家们能得到更为精确、全面、系统的河口潮间带底栖生境监测与评价结果[6]。
本文从关注空间和时间尺度的研究出发,以满足宏观河口管理的需求为目标,在现行SIT技术快速发展的背景下,对河口潮间带栖息地监测、分类与评价方面的研究进展进行了归纳总结,分为以下六个方面进行论述:河口潮间带底栖生境的时空新特征、河口潮间带底栖生境特征参数、河口潮间带底栖生境特征获取方式、数据处理、河口潮间带底栖生境分类模型选择、多时空尺度分析。以期构建大尺度上河口底栖物理生境分类指标体系和分类方法,为河口底栖生境分类和基于生境的生态保护及水生态修复措施等管理策略的制订提供依据。
1 河口潮间带底栖生境的时空新特征
近20年来,广大科研工作者已经在河口潮间带生境变化与底栖动物群落的结构、数量动态、时空分布格局、次级生产力、食性和营养级、重金属富集毒理等方面的关系,进行了广泛而深入的研究。在这些研究中,研究者面临的首要问题是对潮间带底栖生境进行划分和描述。
通常,“生境”或“栖息地”(habitat)的定义为维持特定物种或群落生存所需的空间和其中全部物理、化学、其它生物因子的总和[7-8]。对栖息地定义的理解,导致划分栖息地类型的研究大多直接依据环境中的生物或非生物因子。研究者对河口潮间带底栖生物栖息地的划分,往往也是根据不同的生物及非生物因子分类规则,进行比较研究,分类规则大体分为三类[9]:一、根据某一主要生态或环境因子分类,如盐沼植物生境(海草、红树林)[10-11]、潮沟生境[12-13]、盐度梯度生境[14]、潮上中下带生境[15];二、综合一组河口潮滩湿地中差异性最大的环境因子[16],或者一组对生物群落影响最大的环境因子进行分类,如构成生境适宜性指数(habitat suitability index,HSI)[17]、环境指数(index of environmental condition,IEC)[18]、生态整合度指数(index of ecological integrity,IEI)[19]进行生境分类;三、划分为人工干扰生境与自然生境,如大型航道工程生境[20]、土方工程生境[21]、围垦生境[22]。
然而,这三种分类方式都有一定的局限性。在小区域范围内进行短期静态研究时,或影响目标物种生存的栖息地特征因子较为简单的情况下,以上三种方法是合适的[23]。但是河口是河流与海洋的结合地段,受河流终段和近海水域相互影响,因此,河口潮间带是一类空间高度异质化的复杂体,包括光滩、潮汐、盐沼植被等,其生物和非生物因子随着不同的生境有着很大的变化,如植物、水文、地形、土壤、盐度、浮游生物等。另外,受到潮汐、风暴及人类活动的影响,这些物理生化环境因子会在很小的空间范围内(有时小于100 m)和时间尺度上发生变化[24]。因此,对如此诸多、动态变化的环境因子进行大规模采集存在人力、物力的限制,传统的栖息地定义及随之产生的分类规则亦不能全面满足监测和评价的需求,亟需从时空角度上考虑栖息地新特征,确定新的分类规则和采样手段,对复杂多变的河口潮间带底栖生境进行分类,用以满足长期、便捷、更加精确监测其动态变化的需求[8]。
近年来,随着SIT技术的发展以及对空间生态学概念理解的加深,可以利用GIS对栖息地类型特征或空间单元(斑块)赋予空间变量特征,记录其在范围、面积、形状、斑块位置、与邻近斑块空间关系的变化等空间结构和纹理信息[8]。因此,基于栖息地斑块的内在属性和空间性质[25],可以将栖息地特征重新划分为三类:(1)属性特征(typology):指生物和非生物因子特征,如地形、地貌、植物群落组成、植物优势度、植物演替阶段、其它动物群落组成等。通过栖息地斑块属性特征的相似性,或基于此运算得到的栖息地指数确定不同栖息地类型(以往的三类分类研究属于基于该类特征)。(2)纹理特征(texture):关注每一种栖息地类型的斑块数目、大小及其频度分布。当每种栖息地类型中斑块数量和面积大小计算出来时,即可对纹理特征进行量化。(3)结构特征(structure):由栖息地斑块的空间结构(范围、面积、形状)、布局特征以及斑块之间的空间地理关系(斑块位置、与邻近斑块空间关系的变化)确定。空间结构可以由多种空间统计方法或空间指数描述(如破碎化指数、景观指数等)。栖息地斑块空间性质、纹理和结构特征能反映多种环境因子下不同时空尺度上生境的综合状况。通过描述栖息地的纹理和结构特征这类新属性,研究其与生物种群、群落之间的关系,能从与栖息地水平相关的多尺度条件下,描述和理解生物多样性状态和变化机制。
2 河口潮间带底栖生境特征参数
一般河口潮间带底栖生境的状态是通过对上述的属性特征(物理-化学性质、物种组成和/或种群相对丰度)以及纹理、结构空间特征(斑块面积、分布、数目、破碎化程度等)进行采集而评估的。除了生境斑块的内在特征外,特征信息采集通常还涉及一些额外信息,如水文、生物群落生理生化指标、以及潜在的驱动因子和人类胁迫因子(土地利用方式等)。表1根据以往文献总结了这些生境特征数据组及额外信息的类型以及获取方式[1,26-28]。
3 河口潮间带底栖生境特征获取方式
根据主要的数据来源,河口潮间带底栖生境特征获取方式主要分为野外采样和辅以地面验证数据的遥感监测方式。
3.1 野外采集生物及底泥信息
野外采样是指基于野外调查和测量,通常采用固定的样点或样条,在小尺度上探查栖息地中物种组成或相对丰度。在未使用SIT技术之前,一般通过对某些区域实施抽样采样的调查方法,通过先验的随机法或后验的空间模型法(外推法),推测其它区域物种组成或相对丰度。野外采样能有效记录局部和区域尺度上底栖物种空间分布结构以及其环境决定因子空间变化。然而,野外采样的方法十分耗时耗力,同时还需专业的分类学家参与。因此,野外采样制图,很少能覆盖完整的研究区域,并且小尺度上的异质性格局通常被忽略。单单通过野外采样,不可能设计出栖息地高分辨率的采样方案,更不能实现对栖息地进行长期监测的目标[1]。
随着遥感技术的发展,潮间带栖息地的大多环境属性特征能得到有效地大范围、连续精确监测[8](见3.2),但其底栖环境的大部分生物特征和底泥特征不能直接被遥感技术测量。因此,为了阐明底栖生物群落及其生境之间的关系,利用原位采样的方法,采集底栖环境的生物和底泥特征必不可少(表1)。
3.2 遥感监测生境环境属性特征
利用遥感技术能对河口潮间带的多种环境因子进行识别,从而实现对栖息地多种环境因子大规模连续监测的目标。近几十年来,已有研究者利用多种遥感源对河口栖息地环境特征进行监测[1],例如,航拍图像、卫星影像(潮间带最常用的卫星是Quickbird、IKONOS、SPOT、Landsat TM/ETM),高光谱图像、声学遥感。河口潮间带的多个主要环境特征已得到了成功地提取和反演,如高程[29]、植被[30]、土地利用[31]、潮沟[32]、土壤[33](表1)。
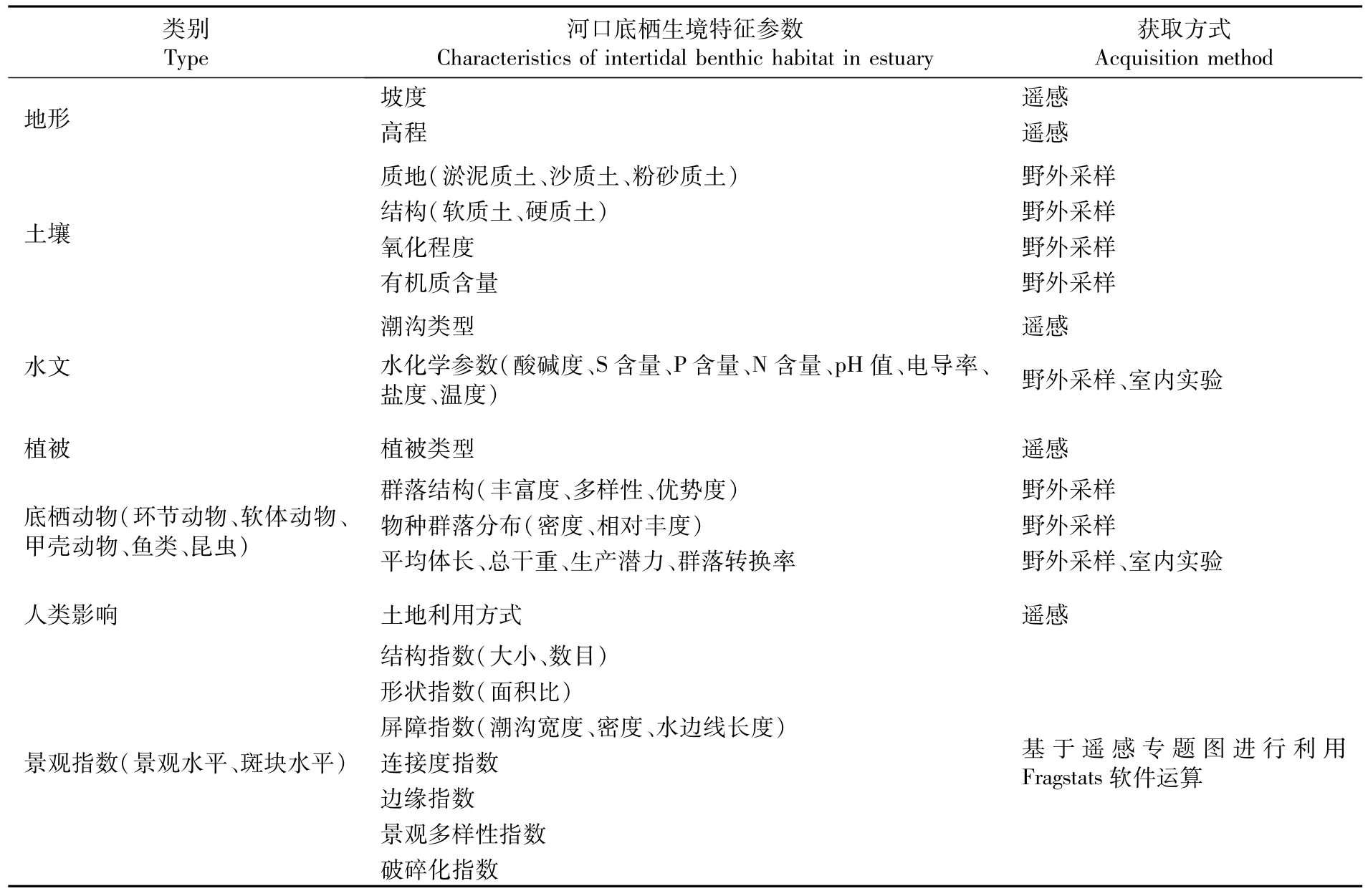
表1 河口潮间带底栖生境特征参数及其主要获取方式Tab.1 Characteristics of intertidal benthic habitat in estuary and their acquisition methods
与野外采样相比,遥感有明显的优势。例如避免对河口潮间带造成物理侵入或人为的干扰、不需要太多的生态专业知识[8]、可以得到连续覆盖的大尺度环境数据层,尤其是高分辨率数据,能在更小的尺度上辨识和监测高度破碎化的潮间带底栖生境系统[34]。
在大部分的研究中,河口潮间带底栖生境图是通过遥感环境数据和原位采样数据之间的关系推译出来的。在这种方式下,河口潮间带底栖生境分布估测图最终是由完整的环境数据作为指示因子所绘制成图。
3.3 Fragstats软件提取生境空间特征
栖息地斑块的空间特征与种群和群落的多个生态过程和分布格局密切相关,如物种关系、繁殖、分散、干扰等。ZIV[35]认为栖息地类型越多越复杂,即多样性(diversity)程度高,将为更多的生物种群创造更多的生存机会及更大的栖息地选择权;栖息地斑块大小(patch size)是决定种群数量及灭绝速率的重要因子,斑块面积越大,可支持的种群数量越多、灭绝速率越低;栖息地斑块性(patchiness)越大,即同一栖息地类型中斑块数量越多,一个群落中个体在整个区域生存机会越大,灭绝速率越低,种群的稳定性越高。
遥感易于辨识各种栖息地斑块的空间特征,如纹理和结构特征,然后通过简单的运算得到景观指数(landscape metrics)进行量化。利用Fragstats软件工具可以便捷地通过遥感专题图计算河口潮间带生境景观指数,以代表生境在不同尺度上的景观结构、形状、屏障、连接度、边缘、多样性等生境特征(表1)。通过量化的景观指数,对栖息地进行分类,例如分为破碎性生境、连续性生境、边缘锐化生境、边缘平滑生境、多屏障生境、多廊道生境。目前,研究者已经就应用景观特征描述河口潮间带生境格局和动态变化方面进行了相关研究[36-38],并通过景观指数定量分析景观特征对大型底栖动物群落结构和分布的影响,证实了河口潮间带生境斑块大小、斑块数目、斑块组成和斑块连接度对大型底栖动物物种多样性、物种优势度、物种丰度有显著影响[39-41]。例如,DE JUAN等[40]通过提取新西兰八大河口区六大生境类型的生境破碎化指数、斑块大小和数目等景观指数,将其与物种多样性指数进行相关分析。结果表明,斑块大小对河口大型底栖动物物种多样性有显著影响。BEATRIZ等[41]深入研究了巴西圣保罗北部海岸生境破碎化后,边缘景观、斑块大小、斑块组成对潮下带底栖动物群落的影响。研究表明,生境破碎化后尽管小斑块的边缘效应增加了物种的丰度,但物种优势度和多样性大大降低。THRUSH等[42]通过研究生境斑块数目、斑块大小、斑块连接度等指数与大型底栖动物群落丰度、总密度和种群组成的关系。结果表明,物理环境上的景观连接度对大型底栖动物群落结构的恢复起了重要作用。
4 数据处理
随着SIT技术的发展,一方面关于栖息地状态的数据来源更加丰富,另一方面,处理这些数据的手段也更为多样。因此,研究者可以更加全面考虑生态过程和格局中所涉及的空间生态学问题,最终使物种/群落栖息地分类结果更加精确、真实、全面。
一个全面的数据处理程序应该包括以下步骤[43]:一、整合相关数据,使之可用于同一分析过程中,包括多源遥感数据融合、遥感数据与野外采样数据整合;二、评价数据的充分性(数据精确性、全面性、完整性),包括生物数据的零值处理、遥感专题图分类精度优化、生态特征变量代表层次选择;三、处理高度相关的环境变量——空间自相关分析;四、栖息地连续特征变量的边界分割。
4.1 数据整合
由于栖息地特征获取方式多样,数据来源多元化,在栖息地分类制图前,需要对元数据进行数据整合。通常有二类数据整合方式:一、不同来源的遥感数据融合。不同分辨率的原始遥感数据代表的尺度不同,例如小尺度数据无法简单代表大尺度数据[8],因此,由空间、光谱分辨率差异导致的尺度问题对栖息地分类有较大影响。已有大量文献对不同遥感来源的数据融合方法进行了综述[1,44],主要有象元(pixel)、特征(feature)和解译(interpretation)水平的融合;二、遥感数据与野外采样数据整合[44]。这类整合的优势为能将栖息地特征进行互补,利用高精度的野外采样数据用以支持遥感数据或对遥感解译图像进行验证,或提供遥感所不能获取的底栖环境额外信息,如土壤信息等。另外,遥感能用作野外采样的空间信息的补充数据或校正依据,提供在野外采样中难以探查的栖息地空间格局信息,如破碎化、连接度等。但野外采样数据大多为位点上的离散数据,需要在GIS平台上对其进行栅格化或矢量化后,与连续的遥感数据进行整合。
4.2 零值处理
在对底栖种群或群落的野外采样中常常面临大量的零值数据,影响随后的科学分析。零值数据的来源主要有三类[45]:一、真实零值,表示研究区内真的不存在某一物种;二、假零值,也称为伪零值,这类零值是对物种较低的采样概率所导致;三、采样干扰零值,已知某一物种常出现在某地,但由于侵入式的采样手段导致其惊吓躲避从而无法记录采样。目前已有很多方法用于处理这三类采样时产生的零值[46]。最简单的方法是在数据处理前剔除零值。但简单地去除零值常常会对结果产生影响,使分析结果出现偏差[46-47]。另一种方法是把所有的观测值加一个正值的常量,接着进行对数函数(Log)转化。这一方法要求人为选择一个常量,这个常量的选择不当将造成估计值偏差[47]。有研究对此方法进行改进,利用观测值的标准分布概率函数(Delta对数正态或Gamma函数)得到一个二次型分布正值代替该常数。此法称为Delta法[48],由于这一方法中常量是非零值观测值的产物,大大减少了估值偏差。
4.3 遥感专题图分类精度优化
遥感解译得到土地利用/土地覆盖专题图是运算景观指数的基础,遥感影像专题图分辨率(由土地覆盖分类策略决定)代表地理空间信息的详细程度,对景观元素(斑块等)分类及景观指数分类有重要影响[49]。改变分类图像的专题分辨率同样将导致景观类别数目和空间格局的变化,从而导致景观指数的变化,许多研究结果证明了专题图分辨率对大多数景观指数有显著的影响[49-51]。解决这一问题的方法是在允许的条件下,尽可能使用高空间分辨率的遥感影像进行解译得到土地利用/土地覆盖专题图。高空间分辨率影像具有丰富的空间信息。因此在高分辨率影像上,同一地物表现出更大的差异,得到的景观特征更加丰富。但传统的基于像元光谱信息的分类方法会由于同一地物的光谱异质性造成严重的“椒盐效应”(在一个小区域内,同种地物被分类成不同的类别,原本均一的地块被“打碎”的效应),目前取而代之的是面向对象分类方法对高空间分辨率遥感影像进行分类。该分类法能够结合形状、纹理、拓扑、层次结构等空间信息,较其它空间分辨率遥感影像,可将河口潮间带植被分类精度提高10%~25%[30]。
4.4 生境特征变量层级选择
随着SIT技术和空间生态学的发展,生态学家已经认识到物种和群落的分布依赖于尺度变化和在特定尺度上的栖息地异质性。在不同的尺度上,影响生物种群或群落结构和分布格局的关键栖息地特征不尽相同,往往从大尺度到小尺度上,这些关键性的栖息地特征之间具有层级性(hierarchical)关系[52]。MACEDO等[52]列举了在河口流域尺度和局域尺度影响底栖动物群落结构与分布的栖息地特征因子,如局域尺度上重要栖息地特征为潮沟、底泥、水流、水化学和物理指标、水生植被、人为捕捞因子;而在流域尺度上的重要栖息地特征为地形、地貌、气候、自然植被、草地、农用地、城市、人口密度、道路。因此,在选择和处理影响生物种群或群落结构和分布的栖息地所有潜在特征因子时,需要根据不同的研究目的,选择代表不同尺度下合适等级的生境特征类别。或者预先通过尺度效应分析,明确不同尺度下影响生物种群或群落结构和分布的特定栖息地特征,再行选择分析。
4.5 空间自相关分析
生物数据(如丰富度)以及生境特征数据在地理空间上具有相关性,即空间格局本身不独立,存在由相邻个体变量的相互作用而产生的空间依赖性,或称为空间自相关性[53-54]。若不能处理好数据的这一特征,将对模型参数估值及其不确定性造成影响,如栖息地模型的结果将产生空间相关的残差、有偏差的误差变量、错误的显著水平和过高的回归系数[55],最终导致不正确的统计推断。因此,在选择栖息地模型进行分类研究时,需要选择能处理空间自相关性的模型。目前较为常用的、整合考虑了空间自相关性的种群栖息地模型包括[43]:自回归方法、基于克里金插值(Kriging)的地统计法、广义线性混合模型,广义评估函数、地理权重回归法等。
4.6 栖息地连续特征变量的边界分割
随着SIT技术的发展,高度异质性的潮间带底栖生境的大部分环境特征能被遥感所探查,所解译的连续环境数据层能在GIS系统中进行空间分析和处理。为了能清晰地表示生物及其栖息地的空间分布格局,在对栖息地不同类型进行划分时需要确定相邻类别之间的边界,这意味着需要对连续的环境数据层进行分割(segmentation)。以往,一般相邻的栖息地类型之间都是由人为划分的清晰边界来分割。然而,在现实情况下,河口栖息地特征之间的边界并非总是清晰明确的(例如岩礁和周围软泥的相交地带),尤其是底栖生境的环境特征经常呈梯度式变化。栖息地特征之间的分割边界可以用更为真实的梯度方式来表达。新的空间统计方法,可以实现利用空间连续的环境数据组以连续或离散的方式描述和预测河口潮间带底栖生物格局。这些方法更容易构建潮间带底栖群落形成的真实情况,并避免了以往僵化固定的离散边界。常用的边界分割方法[7]有专家解译、主成分分析、神经网络法、贝叶斯决策法则、监督分类等。
5 河口潮间带底栖生境分类模型选择
针对不同研究对象,需要选择合适的模型,对栖息地生境特征与生物分布数据进行拟合并模拟分类。已有的河口潮间带底栖生境分类研究可以分为两类[7]:单一底栖物种栖息地分类(single benthic species habitatmapping),底栖群落栖息地分类(benthic communitymapping)。
5.1 单一底栖物种栖息地分类
单一底栖物种栖息地分类所得到的栖息地类型特征必须本质上与目标物种支持环境状况相匹配。因此理论上,生成底栖生境地图最简单的方法是将环境数据组作为指示因子与物种分布数据进行模拟,确定目标物种的栖息地。这是一种先整合、再预测的非监督分类策略,采取的是一种从上至下的方法,在现有的研究中使用最为广泛。
随着GIS和RS的快速发展,研究人员更易获取更高质量和详细的环境数据层,这些环境数据层能够在质量上、空间范围或分辨率上满足目标物种的估测。在应用非监督分类策略时,这些连续的环境数据层预先通过专家解译或目标分类的方法分割(见4.6)为栖息地“空间单元”,然后再利用采样点上获取的物种的生物数据(正常值/缺省值)进行建模分析。建模过程通常利用的是最简单的相关统计方法。根据生物与环境数据组之间的相关关系,对生物物种的出现值(分布值)进行外推(图1-A)。

图1 底栖栖息地分类策略Fig.1 Strategies of benthic habitatm apping
另一种分类策略是先预测,再整合的监督分类策略,这是一种从下至上的方法,较前一种方法更加复杂和有针对性。在此,地面采样所获得的目标物种生物数据(出现值)将作为环境数据预测值的函数,通过物种分布模型(species distribution modelling,SDM)[43],并生成一张物种分布预测图。在物种分布预测图的基础上,原位采样所获得的生物/地理地面验证数据被用作“空间单元”(此处的空间单元为栖息地类型)分割的标准,将物种分布预测图划分物种栖息地分类图(图1-B)。SDM是一类能联系物种分布数据(在已知地点的出现值/丰富度)和这些地点环境或/及空间特征信息的数学模型。相比传统的多元线性回归模型等[43],SDM能在时间和空间上进行外推,利用不充足的数据,实质上能等同表达物种所处环境的物种拟合模型,SDM模型能提供有用的生态学解释和显示强有力的预测能力。
一般情况下SDM方法都是类似回归的,假设一个物种的出现值或丰富度可以通过环境因子的加性结合模拟,或通过表示环境因子之间关系的参数所模拟。SDM模型最常用的模型方法为广义加性模型(generalized addictive model,GAM)[43]。由于其能描述非线性的生态响应(二次方、幂方或其它参数转换),在拟合生态过程中提供了除线性关系以外更为真实的灵活度。另外,利用机器学习和数据挖掘的模拟方法[43],也是近来 SDM发展的热点。如神经网络法(artificial neural networks,ANNs)、多变量适应回归曲线法(multivariate adaptive regression splines)、分类和回归树(classification and regression trees)、最大熵值模型(maximum entropy models)等。这些模型中有一些能提供很好控制的变量选择和系数估计,另一些能自动探测和拟合预测因子之间的关系。为评价模型的模拟结果及避免模型偏差,一般使用CCI值、kappa系数、AIC(akaike information criterion,AIC)值用于表示模拟结果(生物群落分布、栖息地类型)和实测数据之间的拟合程度。已有研究表明,SDM可以成功地用于单一底栖物种栖息地分类研究[57-58],它们预测行为的最终结果能优于大部分传统的方法[56]。
5.2 底栖群落栖息地分类
群落栖息地分类步骤与单一物种栖息地分类类似,同样有两种策略(图1),如非监督分类策略中,群落生物数据通常以组别的形式存在(由多元统计、聚类分析等方法得到),进而与预先分割的环境空间单位进行比较分析,并对群落类别进行外推。
监督分类策略中,进行群落栖息地分类时,理论上可以通过每次生成一种物种栖息地分布图,经过多次后最终将这些单一物种栖息地分布图整合叠加为群落栖息地分布图。
另一种监督分类常用的方法是,首先根据多元统计分析、专家判断、或者已有的生物区系分类系统,对原位采样生物数据进行群落分类。然后,利用这些群落类别以监督分类的形式对连续覆盖的环境数据层进行分割,构成不同的群落栖息地类型。这类群落栖息地分类还可以引入一个群落生境(biotope)的概念,即“一个非生物环境组成的栖息地及其相关的生物群落的结合”[59]。这一术语能在空间上有效表达群落信息。从近年来大量栖息地分类研究来看,亟需发展基于群落生境概念的栖息地分类策略[43],这样的栖息地分类规则具有层级结构,如高层级由大范围的非生物因子(地理、地质因子)构成,低层级由在生物区系水平的物种信息构成,这样所形成的栖息地分类系统既不会太粗略,又不会太过于细致(过多种群的细节信息),不用太过考虑集群结构的季节性变化,更有效地达到和实施保护目的[59]。
在潮间带区域,栖息地以梯度式变化环境特征为主,并且在其非生物环境因子剧烈变化(潮水波动)、小尺度环境异质性极高的区域,群落格局容易被掩盖,这一分类策略由于可以采用群落生境进行群落栖息地分类,其层级结构可以实现在不同空间尺度上识别底栖群落格局,但目前在潮间带的研究还鲜有报道。
6 多时空尺度分析
随着对物种和群落的栖息分布存在尺度效应这一认识的发展,对于复杂多变的河口潮间带生态系统,在单一空间尺度分析栖息地特征与底栖生物群落分布之间的关系,已经不能满足生态管理和保护的要求,空间层次法(spatial hierarchical approach)中多空间尺度分析(multiple scales analysis)可以用于解决这一问题[60]。
多空间尺度分析是根据系统的层级组织特征,将大尺度由上自下分为多个层级(level)小尺度的方式,在各个层级的空间尺度下,测试变量敏感性的方法[60]。采用多空间尺度的分析方法避免了人为随意确定研究物种或群落区域尺度的问题。可量化、多层次的多个空间尺度范围使分析群落与环境之间的关系具有可比性,可以明确群落在不同空间尺度上格局与过程中的变化和不确定性,得到更为全面和精确的结果和评价。
尤其是利用景观指数作为栖息地空间特征的指示参数时,必须进行尺度效应分析。已有大量研究围绕景观指数在不同尺度上与大型底栖动物群落的关系展开,证明了在不同尺度上,潮间带底栖动物群落对生境景观特征的响应程度不同。THRUSH等[61]分别在样点和景观尺度上分析了新西兰北岛卡沃湾的景观特征与底栖生物群落多样性的相关关系。结果表明,在样点尺度上,局部的景观连接度对物种多样性影响显著,但影响程度较弱;在更广的景观尺度上,生境的连接度对物种多样性增加效果更为显著且有力。POTTER等[62]分别在流域尺度和河岸区域尺度上对北卡罗来纳州73个站点提取景观特征,并研究其与底栖动物群落的关系。结果表明,流域尺度上提取的景观特征(解释度为69.5%~75.4%)比河岸区域尺度上提取的景观特征(解释度为57.4%~65.2%)能更好地解释大型底栖动物群落结构的变化。综上所述,量化景观指数,分析栖息地空间特征与底栖动物群落的关系必须在多时空尺度下进行。
另一个方面,大多重要的大型底栖物种为运动型(如蟹类、虾类、鱼类),分布范围广,且在不同的生活史阶段(如觅食、避难、繁殖和育幼等)以不同的方式利用潮间带栖息地资源。为了生成合理、准确的栖息地地图,亟需深刻理解这些重要的底栖物种生活史阶段栖息地。这些地图需要进行多时间尺度上(月变化、季节变化、年变化)的动态分析,用于反映由季节或生物个体自身遗传引起的栖息地偏好性在生活史各个阶段的时空动态变化。然而,已有一些研究率先开展了对潮间带底栖生物(蟹[63]、虾[64]等)的长期动态监测,探索其栖息地时空动态选择及变化规律。
7 小结与展望
SIT技术和空间生态学的发展,使获取栖息地的空间信息及动态的时间信息具有多样性与多维性。例如,栖息地类型特征或空间单元(斑块)被赋予空间变量特征;通过遥感技术,大多潮间带环境属性特征能得到有效地大范围、连续精确监测,河口潮间带底栖生境特征能得到更为全面、系统、精确地描述。另一方面,数据来源丰富多样,数据处理手段更为多样,分类方式可以更加全面考虑生态过程和格局中所涉及的空间生态学问题(边界分割、尺度效应、分类精度、动态变化等问题)。最后,空间模型和群落生境概念的发展使得底栖物种及群落分布的模拟更为便捷,最终使物种/群落栖息地分类结果更加精确、真实、全面。
潮间带底栖栖息地分类对潮间带底栖生态系统进行生态评价十分重要。例如,在对潮间带底栖栖息地进行精确分类的基础上,可以选择与大型底栖生物群落栖息地质量密切相关的主要环境因子作为评价指标。应用栖息地质量量化模型对影响因子进行比较并赋予权重和分值以建立潮间带底栖栖息地质量评估体系。
另外,在潮间带底栖栖息地分类的基础上,还可以结合潮间带栖息地多样性、底栖生物多样性与食物网结构等关系,研究河口潮间带底栖栖息地随着环境梯度、地理空间上的变化,在局部和区域尺度上其生态功能的变化机制。
显而易见,利用SIT对河口潮间带底栖生境进行分类将对底栖生态系统评价及研究生态功能机制带来新的视角。然而,利用SIT技术目前也存在一些问题。例如,利用景观指数作为栖息地空间特征的指示参数时,不是所有的参数都具备显著性的指示作用,需要选择合适的统计方法在不同的尺度上进行甄别和筛选,才能有效提高指示参数在研究尺度上的解释度。另外,由于新发展的物种分布模型等空间统计方法较传统的数学方法更为复杂,现阶段在数据处理和模型模拟过程中,这些优势明显的方法在河口潮间带底栖生态系统监测和评价研究中还未能大范围进行应用,亟待发展和推广。
参考文献:
[1] GODET L,FOURNIER J,TOUPOINT N,et al.Mapping and monitoring intertidal benthic habitats:a review of techniques and a proposal for a new visual methodology for the European coasts[J].Progress in Physical Geography,2009,33(3):378-402.
[2] ZHARIKOV Y,SKILLETER G A,LONERAGAN N R,et al.Mapping and characterising subtropical estuarine landscapes using aerial photography and GIS for potential application in wildlife conservation and management[J].Biological Conservation 2005,125(1):87-100.
[3] MCLUSKY D S.The estuarine ecosystem[M].New York:Chapman&Hall,1989.
[4] 沈新强,陈亚瞿,罗民波,等.长江口底栖生物修复的初步研究[J].农业环境科学学报,2006,25(2):373-376.
SHEN X Q,CHEN Y Q,LUO M B,et al.Preliminary study on the restoration of benthos in the Changjiang Estruary[J].Journal of Agro-Environment Science,2006,25(2):373-376.
[5] HESS G R,CHESHIRE H M.Spatial information technologies-integrating the tools with the curricula[J].Journal of Forestry,2002,100(1):29-34.
[6] 张婷婷,张 涛,侯俊利,等.空间信息技术在渔业资源及生态环境监测与评价中的应用[J].海洋渔业,2014,36(3):272-281.
ZHANG T T,ZHANG T,HOU J L,et al.Application of spatial information technologies in monitoring and assessment of fishery resources and ecological environment[J].Marine Fisheries,2014,36(3):272-281.
[7] BROWN C J,SMITH S J,LAWTON P,et al.Benthic habitat mapping:A review of progress towards improved understanding of the spatial ecology of the seafloor using acoustic techniques[J].Estuarine Coastal and Shelf Science,2011,92(3):502-520.
[8] LENGYEL S,KOBLER A,KUTNAR L,et al.A review and a framework for the integration of biodiversity monitoring at the habitat level[J].Biodiversity and Conservation,2008,17(14):3341-3356.
[9] SHOKRI M R,GLADSTONE W.Limitations of habitats as biodiversity surrogates for conservation planning in estuaries[J].EnvironmentalMonitoring and Assessment,2013,185(4):3477-3492.
[10] 童春富,章飞军,陆健健.长江口海三棱藨草带生长季大型底栖动物群落变化特征[J].动物学研究,2007,6(6):640-646.
TONG C F,ZHANG F J,LU J J.Variation characteristics of the macrobenthic fauna community in the Scirpusmariqueter zone of the Yangtze Estuary during the growing seasons[J].Zoological Research,2007,6(6):640-646.
[11] 谢志发,何文珊,刘文亮,等.不同发育时间的互花米草盐沼对大型底栖动物群落的影响[J].生态学杂志,2008,27(1):63-67.
XIE Z F,HE W S,LIU W L,et al.Influence of Spartina alterniflora salt marsh at its different development stages on macrobenthos[J].Chinese Journal of Ecology,2008,27(1):63-67.
[12] RODRIGUESA M,QUINTINO V,SAMPAIO L,et al.Benthic biodiversity patterns in Ria de Aveiro,Western Portugal: Environmental-biological relationships[J].Estuarine Coastal and Shelf Science,2011,95(2-3):338-348.
[13] 宋慈玉,储忝江,盛 强,等.长江口盐沼分级潮沟系统中大型底栖动物群落结构特征[J].复旦学报(自然科学版),2011,50(3):253-259.
SONG C Y,CHU T J,SHENG Q,et al.Macrobenthos communities in salt marshes of the Yangtze River Esturay along a creek-order gradient[J].Journal of Fudan University(Natural Science),2011,50(3):253-259.
[14] BARNESR S K.Distribution patterns of macrobenthic biodiversity in the intertidal seagrass beds of an estuarine system,and their conservation significance[J].Biodiversity and Conservation,2013,22(2):357-372.
[15] 全为民,赵云龙,朱江兴,等.上海市潮滩湿地大型底栖动物的空间分布格局 [J].生态学报,2008,28(10):5179-5187.
QUANW M,ZHAO Y L,ZHU J X,et al.The spatial pattern ofmacrozoobenthic communities in the tidal wetlands of Shanghai City[J].Acta Ecologica Sinica,2008,28(10):5179-5187.
[16] GAEVER S V,OLU K,DERYCKE S,et al.Metazoan meiofaunal communities at cold seeps along the Norwegian margin:Influence of habitat heterogeneity and evidence for connection with shallow-water habitats[J].Deep-Sea Research Part I-Oceanographic Research Papers,2009,56(5):772-785.
[17] DEGRAER S,VERFAILLIE E,WILLEMSW,et al.Habitat suitability modelling as a mapping tool for macrobenthic communities:An example from the Belgian part of the North Sea[J].Continental Shelf Research,2008,28(3):369-379.
[18] ENGLE V D,SUMMERS J K,GASTON G R.A benthic index of environmental-condition of Gulf of Mexico estuaries[J].Estuaries,1994,17(2):372-384.
[19] KASHIAN D R,BURTON T M.A comparison of macroinvertebrates of two Great Lakes coastal wetlands:Testing potential metrics for an index of ecological integrity[J].Journal of Great Lakes Research,2000,26(4):460-481.
[20] LERCARID,DEFEO O.Variation of a sandy beach macrobenthic community along a human-induced environmental gradient[J].Estuarine Coastal and Shelf Science,2003,58(3):17-24.
[21] 章飞军,童春富,谢志发,等.长江口潮间带大型底栖动物群落演替[J].生态学报,2007,27(12):4944-4952.
ZHANG F J,TONG C F,XIE Z F,et al.The recolonisation progress of intertidal benthic fauna community in the Changjiang Estuary[J].Acta Ecologica Sinica,2007,27(12):4944-4952.
[22] NASER H A.Effects of reclamation on macrobenthic assemblages in the coastline of the Arabian Gulf:A microcosm experimental approach[J].Marine Pollution Bulletin,2011,62(3):520-524.
[23] DIAZR J,SOLANM,VALENTE RM.A review of approaches for classifying benthic habitats and evaluating habitat quality[J].Journal of Environmental Management,2004,73(3):165-181.
[24] BENDELL L I,WAN P C Y.Application of aerial photography in combination with GIS for coastal management at small spatial scales:a case study of shellfish aquaculture[J].Journal of Coastal Conservation,2011,15(4):417-431.
[25] FIRBANK L G,BARR C J,BUNCE R G H,et al.Assessing stock and change in land cover and biodiversity in GB:an introduction to Countryside Survey 2000[J].Journal of Environmental Management,2003,67(3):207-218.
[26] BRUNS D A.Macroinvertebrate response to land cover,habitat,and water chemistry in a miningimpacted river ecosystem:A GISwatershed analysis[J].Aquatic Sciences,2005,67(4):403-423.
[27] JENSEN O P,CHRISTMAN M C,MILLER T J. Landscape-based geostatistics:a case study of the distribution of blue crab in Chesapeake Bay[J].Environmetrics,2006,17(6):605-621.
[28] RAKOCINSKIC F.Evaluatingmacrobenthic process indicators in relation to organic enrichment and hypoxia[J].Ecological Indicators,2012,13(1):1-12.
[29] 韩 震,恽才兴,戴志军,等.淤泥质潮滩高程及冲淤变化遥感定量反演方法研究——以长江口崇明东滩为例[J].海洋湖沼通报,2009,11(1):12-18.
HAN Z,YUN C X,DAIZ J,et al.Remote sensing quantitative inversion research of the silt tidal flat elevation and deposition and erosion—Chongming Dongtan as the example[J].Transactions of Oceanology and Limnology,2009,11(1):12-18.
[30] OUYANG Z T,GAO Y,XIE X,et al.Spectral discrimination of the invasive plant Spartina alterniflora at multiple phenological stages in a saltmarsh wetland[J].Plos One,2013,8(6):e67315.
[31] ZHANG M Q,GUO H Q,XIE X,et al.Identification of land-cover characteristics using MODIS time series data:An application in the Yangtze River Estuary[J].Plos One,2013,8(7):e70079.
[32] 郭永飞.长江口水边线和潮沟遥感信息提取研究[D].上海:上海海洋大学,2011.
GUO Y F.The study on waterside line and tidal channel remote sensing information extraction in the Yangzte River Estuary[D].Shanghai:Shanghai Ocean University,2011.
[33] 彭 飞.长江口南汇潮滩土壤湿度遥感研究[D].上海:上海海洋大学,2012.
PENG F.Soil moiture quantitative study of the Nanhui tidal flat in the Yangzte River Esturay[D].Shanghai:ShanghaiOcean University,2012.
[34] FREITASR,RODRIGUESA M and QUINTINO V.Benthic biotopes remote sensing using acoustics[J].Journal of Experimental Marine Biology and Ecology,2003,285:339-353.
[35] ZIV Y.The effect of habitatheterogeneity on species diversity patterns:a community-level approach using an object-oriented landscape simulation model(SHALOM)[J].Ecological Modelling,1998,111(2-3):135-170.
[36] 丁 骏.长江口中华鲟栖息地景观格局变化及其适宜性评价[D].上海:上海海洋大学.2012.
DING J.The Chinese sturgeon habitat landscapepattern change in the Yangtze Estuary and its habitat suitability assessment[D].Shanghai:Shanghai Ocean University,2012.
[37] 平 原.南汇边滩湿地景观格局分析和生态系统健康评价[D].上海:华东师范大学,2010.
PING Y.The landscape pattern analysis and assessment of wetland ecosystem health for Nanhui Biantan,Shanghai[D].Shanghai:East China Normal University,2010.
[38] 孙永光.长江口不同年限围垦区景观结构与功能分异[D].上海:华东师范大学,2011.
SUN Y G.Differentiation of landscape structure and function during different reclamation time at the Changjiang River Estuary[D].Shanghai:East China Normal University,2011.
[39] PIERRI-DAUNT A B,TANAKA M O.Assessing habitat fragmentation on marine epifaunal macroinvertebrate communities:an experimental approach[J].Landscape Ecology,2014,29(1):17-28.
[40] JUAN SD,HEWITT J.Relative importance of local biotic and environmental factors versus regional factors in driving macrobenthic species richness in intertidal areas[J].Marine Ecology Progress Series,2011,423(1):117-129.
[41] BEATRIZ P,TANAKA M O.Assessing habitat fragmentation on marine epifaunal macroinvertebrate communities:an experimental approach[J].Landscape Ecology,2014,29(1):17-28.
[42] THRUSH SF,HALLIDAY J,HEWITT JE,et al.The effects of habitat loss,fragmentation,and community homogenization on resilience in estuaries[J].Ecological Applications,2008,18(1):12-21.
[43] ELITH J,LEATHWICK J R.Species Distribution Models:Ecological Explanation and Prediction Across Space and Time[J].Annual Review of Ecology Evolution and Systematics,2009,40(40):677-697.
[44] WANG K,FRANKLIN S E,GUO X L,et al.Problems in remote sensing of landscapes and habitats[J].Progress in Physical Geography,2009,33(6):747-768.
[45] LECOMTE JB,BENOIT H P,ETIENNE M P,etal.Modeling the habitat associations and spatial distribution of benthic macroinvertebrates:A hierarchical Bayesianmodel for zero-inflated biomass data[J].Ecological Modelling,2013,265(1767):74-84.
[46] MARTIN TG,WINTLE B A,RHODES JR,et al.Zero tolerance ecology:improving ecological inference by modelling the source of zero observations[J].Ecology Letters,2005,8(11):1235-1246.
[47] MAUNDER M N,PUNT A E.Standardizing catch and effort data:a review of recent approaches[J].Fisheries Research,2004,70(2-3):141-159.
[48] STEFANSSON G.Analysis of groundfish survey abundance data:Combining the GLM and delta approaches[J].Ices Journal of Marine Science,1996,53(3):577-588.
[49] BUYANTUYEV A,WU J G.Effects of thematic resolution on landscape pattern analysis[J].Landscape Ecology,2007,22(1):7-13.
[50] HUANG C,GEIGER E L,KUPFER JA.Sensitivity of landscape metrics to classification scheme[J].International Journal of Remote Sensing,2006,27(14):2927-2948.
[51] CASTILLA G,LARKIN K,LINKE J,et al.The impact of thematic resolution on the patch-mosaic model of natural landscapes[J].Landscape Ecology,2009,24(1):15-23.
[52] MACEDO D R,HUGHESR M,LIGEIRO R,et al.The relative influence of catchment and site variables on fish and macroinvertebrate richness in cerrado biome streams[J].Landscape Ecology,2014,29(6):1001-1016.
[53] KOENIG W D.Spatial autocorrelation of ecological phenomena[J].Trends in Ecology&Evolution,1999,14(1):22-26.
[54] LICHSTEIN JW,SIMONSTR,SHRINER SA,et al.Spatial autocorrelation and autoregressivemodels in ecology[J].Ecological Monographs,2002,72(3):445-463.
[55] KISSLING W D,CARL G.Spatial autocorrelation and the selection of simultaneous autoregressive models[J].Global Ecology and Biogeography,2008,17(1):59-71.
[56] ELITH J,GRAHAM C H,ANDERSON R P,et al.Novel methods improve prediction of species'distributions from occurrence data[J].Ecography,2006,29(2):129-151.
[57] GALPARSORO I,BORJA A,BALD J,et al.Predicting suitable habitat for the European lobster(Homarus gammarus),on the Basque continental shelf(Bay of Biscay),using Ecological-NicheFactor Analysis[J].Ecological Modelling,2009,220(4):556-567.
[58] IAMPIETRO P J,YOUNG M A,KVITEK R G.Multivariate Prediction of Rockfish Habitat Suitability in Cordell Bank National Marine Sanctuary and Del Monte Shalebeds,California,USA[J].Marine Geodesy,2008,31(4):359-371.
[59] CONNOR DW,ALLEN JH,GOLDING N,et al.The Marine Habitat Classification for Britain and Ireland Version 04.05.[Z].2004.www.jncc.gov.uk/MarineHabitatClassification(internet version).
[60] NODA T.Spatialhierarchicalapproach in community ecology:a way beyond high context-dependency and low predictability in local phenomena[J].Population Ecology,2004,46(2):105-117.
[61] THRUSH S F,HEWITT JE,LOHRER A M,et al.When small changes matter:the role of cross-scale interactions between habitat and ecological connectivity in recovery[J].Ecological Applications,2013,23(1):226-238.
[62] POTTER K M,CUBBAGE FW,SCHABERG R H.Multiple-scale landscape predictors of benthic macroinvertebrate community structure in North Carolina[J].Landscape and Urban Planning,2005,71(2-4):77-90.
[63] YEUNG C,MCCONNAUGHEY R A.Using acoustic backscatter from a sidescan sonar to explain fish and invertebrate distributions:a case study in Bristol Bay,Alaska[J].Ices Journal of Marine Science,2008,65(2):242-254.
[64] CALLAWAY A,SMYTH J,BROWN C J,et al.The impact of scour processes on a smothered reef system in the Irish Sea[J].Estuarine Coastal and Shelf Science,2009,84(3):409-418.
A review of application of spatial technology in mapping intertidal habitat in estuary
ZHANG Ting-ting,ZHAO Feng,HOU Jun-li,GAO Yu,ZHUANG Ping
(Key Laboratory of East China Sea&Oceanic Fishery Resources Exploitation and Utilization,East China Sea fisheries Research Institute,Chinese Academy of Fishery Sciences,Shanghai200090,China)
Mapping the changes of intertidal benthic habitat(IBH)in estuary provides important scientific proofs and management suggestions formany areas,e.g.,the biological conservation of wetland in estuary,the assessment of benthic habitat quality and ecological recovery of damaged habitat.With the development of spatial information technologies(SIT),scientists have the ability to acquire more explicit,holistic and systematic monitoring and assessment for intertidal benthic habitat in estuary.From the spatial context,this article summarized six aspects of progress in mapping intertidal habitat in estuary recently:(1)new spatialtemporal characteristics of IBH definition in esturary;(2)description of characteristics of IBH in estuary;(3)acquisition methods of IBH characteristics data in estuary;(4)data processing;(5)mapping IBH with differentmodels based on species/community;(6)multiple temporal-spatial scales analysis.Based on these progresses,the IBH in estuary can be fully,systematically and explicitly characterized.By consideration of issues in spatial ecology(boundary segmentation,scales effects,classification accuracy,dynamics change,etc.),the distributionmodelling of benthic species/community and habitatmapping becomemore convenient with more spatial explicitly,realistic and holistic results.Finally,this paper proposed that mapping with spatial information technologieswould bring new prosepects for studies of ecosystem evaluation and ecological function in intertial habitats.Nevertheless,there are some issues that should be carefully deltwith inmapping with spatial information technologies.For example,characteristics of intertidal benthic habitat in estuary should be seriously chosen and identified as the most effective indicators with proper scientific statisitics at different scales.Presently,the spatial information technologies have not been widely applied in mapping intertidal habitat in estuary,which needs urgent development and promotion.
spatial information technologies;fisheries resources;ecological monitoring;ecological assessment;application prospect
S 931
A
1004-2490(2016)04-0424-13
2015-05-26
国家自然科学基金(31400410)
张婷婷(1985-),女,助理研究员,博士,主要从事河口生态监测和评价科研工作。E-mail:zhangtt@ecsf.ac.cn
猜你喜欢
百科探秘·海底世界(2024年2期)2024-02-27 14:37:15
疯狂英语·初中版(2023年7期)2023-08-18 05:01:35
海洋信息技术与应用(2020年4期)2021-01-18 06:21:36
海洋通报(2020年3期)2020-10-20 09:26:36
汉语世界(The World of Chinese)(2018年3期)2018-10-22 01:50:04
石油化工建设(2018年1期)2018-07-10 09:49:44
厦门航空(2018年4期)2018-04-25 10:49:27
少儿科学周刊·少年版(2015年11期)2015-12-17 20:59:10
少儿科学周刊·少年版(2015年11期)2015-12-17 20:57:47
少儿科学周刊·少年版(2015年11期)2015-12-17 20:55:51
