
莲雾碳水化合物及矿质元素含量对其花芽分化的影响
2016-03-01 12:57张绿萍蔡永强金吉林陈守一
贵州农业科学 2016年3期
张绿萍, 蔡永强, 金吉林, 陈守一
(贵州省果树科学研究所, 贵州 贵阳 550006)
莲雾碳水化合物及矿质元素含量对其花芽分化的影响
张绿萍, 蔡永强, 金吉林, 陈守一
(贵州省果树科学研究所, 贵州 贵阳 550006)
为莲雾生产上的产期调节提供理论依据与参考,通过对紫红种莲雾和粉红种莲雾进行催花处理,以自然生长的粉红种莲雾(即不作催花处理)为对照(CK)进行大田试验,探讨莲雾叶片碳水化合物及矿质元素含量对其花芽分化的影响。结果表明:莲雾花芽分化前需要积累足够的可溶性总糖和淀粉等碳水化合物,才能确保喷施催花药剂后花芽的分化;莲雾花芽分化与N积累量的关系不密切,而与P、K积累量的关系密切,较高的P含量和适当低的K含量有利于莲雾的花芽分化;莲雾花芽分化与C/N有密切关系,长时间维持高的C/N有利于其花芽分化。
莲雾; 叶片; 花芽分化; 碳水化合物; 矿质元素
莲雾(Syzygiumsamarangense)系桃金娘科(Myrtaceae)蒲桃属植物,有特殊水果风味和很高的营养价值;在自然环境下,4月中旬抽新穗萌发花芽,5月中旬开花并结果;正季果由于高温、多雨,易引起裂果、落果,且果实着色差、甜度低,严重影响莲雾的果实品质[1]。因莲雾只要枝条成熟、养分充足、环境条件适宜就能进行花芽分化。目前,台湾、海南及福建等地结合修剪、环割、遮阳及化学药剂处理等栽培技术调节莲雾产期,并取得较好效果[2-4]。但实践操作中发现,新梢成熟度、台风和低温等因素均会影响莲雾产期调节的成败,调节成功的莲雾花果通常会聚集在植株的东南侧,而在北侧很少甚至没有[5]。碳水化合物和矿质元素的积累与花芽分化密切相关[6],莲雾作为花芽随时分化型的乔木类果树,生产上经常应用产期调节技术,未见其花芽分化与碳水化合物和矿质元素积累相关性的研究报道。为此,笔者以易催花的粉红种莲雾和不易催花的紫红种莲雾为试材,设自然生长粉红种莲雾为对照,对粉红种和紫红种莲雾进行催花处理,研究莲雾花芽分化与碳水化合物及矿质元素含量的关系,以期为莲雾生产上的产期调节提供理论依据与参考。
1 材料与方法
1.1 试验材料
供试品种为容易催花成功的粉红种和不容易催花成功的紫红种,树龄6年,长势一致,粉红种8株,紫红种4株。
1.2 试验设计
试验于2013年在贵州省果树科学研究所(罗甸)莲雾种植场内进行,园地的土质属紫色壤土,肥力中等。试验共设置自然生长的粉红种莲雾(即不作催花处理)为对照(CK)、紫红种莲雾催花处理(处理1)和粉红种莲雾催花处理(处理2)3个处理。
催花处理:处理1和处理2的催花处理均包含遮阳处理、环割处理和药剂处理。7月5日开始进行单株遮阳(遮光率90%~95%)处理;8月5日进行环割处理;8月27日揭网后喷施催花药剂(48%乐斯本乳剂200倍+80 mg/LNAA+10 g/L尿素+1.25 mL/L爱多收)。取样:遮阳及环割处理期分别于7月5日、7月20日、8月4日及8月19日取样,药剂处理期自8月27日起,每3 d取样1次,到莲雾花芽出现 (9月12日)止。取样方法:按东南西北方向,取新成熟枝梢10片顶端健康叶,共40片叶,将叶片清洗干净后,再用纯净水漂洗1次,吸干叶面水分待用。
1.3 开花情况调查
催花前在树冠中上部随机选定40条枝,平均每个方向10条,于9月25日,调查开花枝条数,根据萌发花芽枝条数/调查枝条数(40条)计算开花枝条率;调查记录枝条的花芽数和叶芽数,根据花芽数/(花芽数+叶芽数)×100%计算枝条萌芽率;统计各处理莲雾的花穗数。
1.4 指标测定
叶片115℃杀青10 min,80℃烘至恒重,植物组织粉碎机粉碎过目备用。可溶性总糖采用蒽酮比色法测定,淀粉含量采用酸水解法测定,全氮含量采用凯氏定氮法测定,磷含量采用钼锑抗比色法测定,钾含量采用火焰光度计法测定,碳含量采用重铬酸钾容量法测定。
1.5 数据处理
采用Microsoft Excel 2013进行。
2 结果与分析
2.1 不同处理莲雾的花芽分化
从表可知,粉红种莲雾经催花处理(处理2)后有4株出现花穗,总花穗数为386串,达96.5串/株,于9月15-20日,大量萌发花芽,新梢萌发较少;紫红种莲雾经催花处理(处理1)后仅1株出现3穗花序,4株均在9月10日开始萌发大量叶芽;不作催花处理的粉红种莲雾(CK)分别于7月中旬和8月底萌发大量新梢,没有成花。

表 不同处理莲雾叶芽和花芽的萌发情况
注:-表示在试验观察期未萌芽。
Note: -, Non germination
2.2 不同处理各时期莲雾叶片可溶性总糖和淀粉的含量变化
2.2.1 可溶性总糖 从图1看出,遮阳及环割处理期,处理1和处理2叶片可溶性总糖含量逐渐上升,处理2于8月27日达峰值,为187.85 mg/g,比处理1高53.01 mg/g。揭网喷施催花药剂后,处理2叶片可溶性总糖逐渐下降,但一直高于处理1和对照;处理1的可溶性总糖含量于9月30日达峰值,为151.62 mg/g;对照的可溶性糖为116.5~139.07 mg/g,明显低于处理2。遮阳处理后,虽然光合作用减弱,减缓了可溶性糖的积累速率,但由于抑制了新梢萌发,整体上还是有利于莲雾植株可溶性总糖的积累;揭网喷施催花药剂后,处理植株的花芽分化或新梢的大量萌发需消耗大量的可溶性总糖,从而致其可溶性糖含量的下降,结合3种处理花芽分化的情况,推测可溶性总糖含量的多少可能决定莲雾能否进行花芽的孕育和发端,而花芽形态分化可能需要一定的可溶性总糖积累量。
2.2.2 淀粉 从图1还看出,随着新梢的萌发和成熟,对照的淀粉含量为10.63%~14.86%。处理1和处理2淀粉含量呈先缓慢下降而后略有上升的变化趋势。揭网喷施催花药剂后,处理1和处理2的淀粉含量快速上升。遮阳及环割处理期,处理2叶片的淀粉下降量比处理1少,揭网喷施催花药剂后,从初期至9月9日,处理2叶片淀粉积累量明显高于处理1,其中8月30日,叶片的淀粉含量为15.56%,比处理1高4.61%; 9月12日,处理2的花芽开始萌发,叶片淀粉含量略有回落;处理1叶片的淀粉含量则继续上升,为16.5%左右,结合3种处理花芽分化的情况,推测叶片淀粉的积累量对莲雾花芽分化具有明显的影响。
2.3 不同处理各时期莲雾叶片碳的含量变化
从图2看出,遮阳和环割处理期,对照叶片的有机碳含量较高,尤其是在萌发新梢成熟(7月中旬)后而下一批新梢未萌芽时(8月19日)最高,达607.39 mg/g,比处理1和处理2分别提高72.9 mg/g和32.53 mg/g;揭网喷施催花药剂后,处理1和处理2叶片的有机碳含量快速上升,分别于9月3日和9月6日达峰值,为582.95 mg/g和603.78 mg/g;花芽或叶芽萌发前(9月9日),又快速减少,处理1由于大量叶芽萌发,其叶片的有机碳含量下降至511.9 mg/g;整个催花处理过程中,处理2由于大量花芽萌发,叶片的有机碳含量明显高于几乎没有花芽萌发的处理1;而对照叶片的有机碳含量的峰值还高于处理2。推测莲雾花芽分化除了需要有一定的有机碳积累量,可能还与是否在短期内快速积累有关。
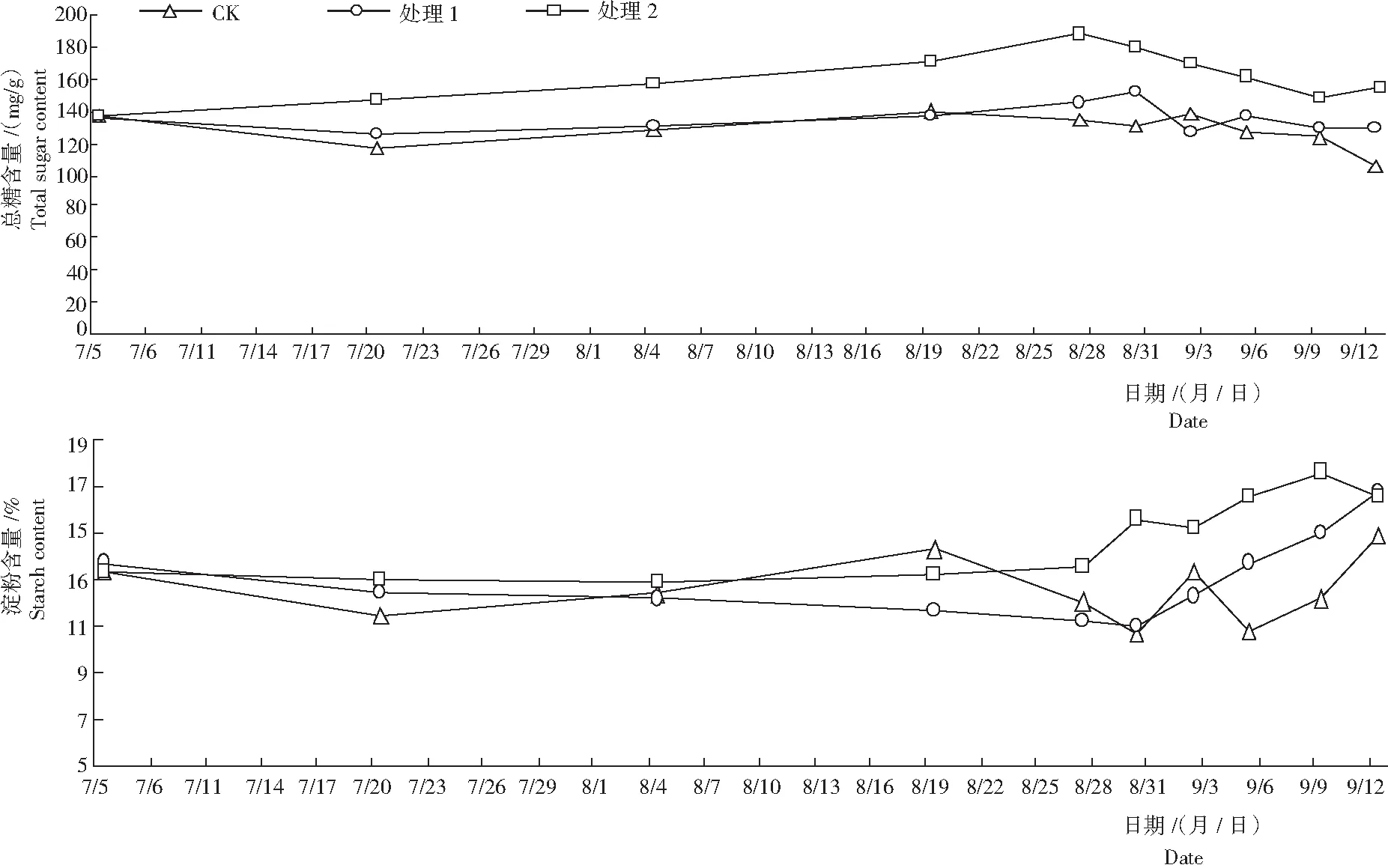
图1 不同处理各时期莲雾叶片可溶性总糖和淀粉的含量
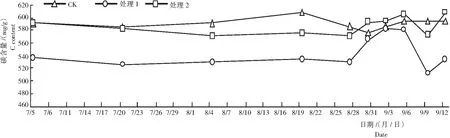
图2 不同处理各时期莲雾叶片碳的含量
2.4 不同处理各时期莲雾叶片氮(N)、磷(P)和钾(K)的含量变化
2.4.1 氮含量 由图3看出,随着新梢萌发和成熟,对照叶片的N含量为17.9~18.9 mg/g,始终高于处理2,8月4日后,同时高于处理1。在遮阳和环割处理期,处理1和处理2叶片的N含量呈平缓的下降趋势;揭网喷施催花药剂初期,处理1和处理2叶片的N含量快速上升,于8月30日达峰值,为18 mg/g左右,而后又急速下降;处理2和处理1分别于9月2日和9月5日达最低值,均为16.1 mg/g,之后2个处理叶片的N含量又开始缓慢上升。经催花处理的粉红种莲雾和紫红种莲雾的氮含量的变化趋势相似,但未见紫红种花芽分化,推测氮含量的多少与莲雾是否进行花芽分化的相关性不大。
2.4.2 磷含量 由图3还看出,随着新梢的萌发和成熟,CK叶片的P含量为2.28~2.57 mg/g。
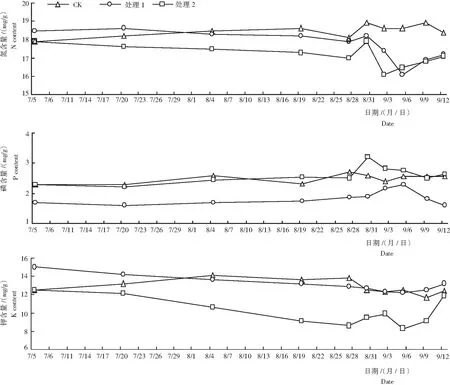
图3 不同处理各时期莲雾叶片氮、磷和钾的含量
Fig.3 Nitrogen, phosphorus and potassium content inS.amarangenseleaves in different growth date under different treatment
在遮阳和环割期,处理1和处理2叶片的P含量呈平缓的上升趋势;揭网喷施催花药剂后,处理1和处理2叶片的P含量快速上升后又逐渐下降,且处理1叶片的P含量一直低于处理2。处理1于9月5日达峰值,为2.29 mg/g;处理2于8月30日达到峰值,为3.2 mg/g;紫红种莲雾(处理1)不易催花成功可能与其较低的P积累量有一定关系。
2.4.3 钾含量 从图3可知,随着新梢萌发和成熟,CK叶片的K含量为11.75~14.07 mg/g。处理1叶片K含量呈缓慢下降趋势,至9月6日后开始回升。在遮阳和环割期,处理2叶片的K含量持续下降,从7月5日的12.48 mg/g下降至8月27的8.63 mg/g。揭网喷施催花药剂后到有花芽萌发期,叶片的K含量呈上升→下降→上升的变化趋势。处理2叶片的K含量明显低于处理1和CK。处理1和CK从8月初开始,两者叶片的K含量较接近。因此,叶片钾含量高可能不利于莲雾花芽分化。
2.5 不同处理各时期莲雾叶片的碳氮比变化
从图4可知,随着新梢萌发和成熟,CK叶片的C/N为28.87~32.98。处理1和处理2叶片的C/N的变化趋势一致,遮阳和环割处理期呈平缓的上升趋势;揭网喷施催花药剂后,C/N快速上升,达峰值后下降至谷值后又回升。8月27至9月2日,处理2叶片的C/N明显高于处理1,最大相差值为4.87,达到峰值后叶片的C/N维持3 d(9月2至9月5日)而后略有下降;处理1叶片的C/N达到峰值后又急速下降。9月12日处理2花芽萌发,C/N达39.48;处理1新梢大量萌发,C/N为31.01,比处理2低8.47。可能是由于新梢的生长比花芽快,短期需要更多的营养物质所致,从而使处理1后期的C/N下降速率比粉红种快,而回升值低于处理2。推测较高的C/N有利于莲雾花芽分化,C/N值低或维持时间短,不利于其花芽分化。
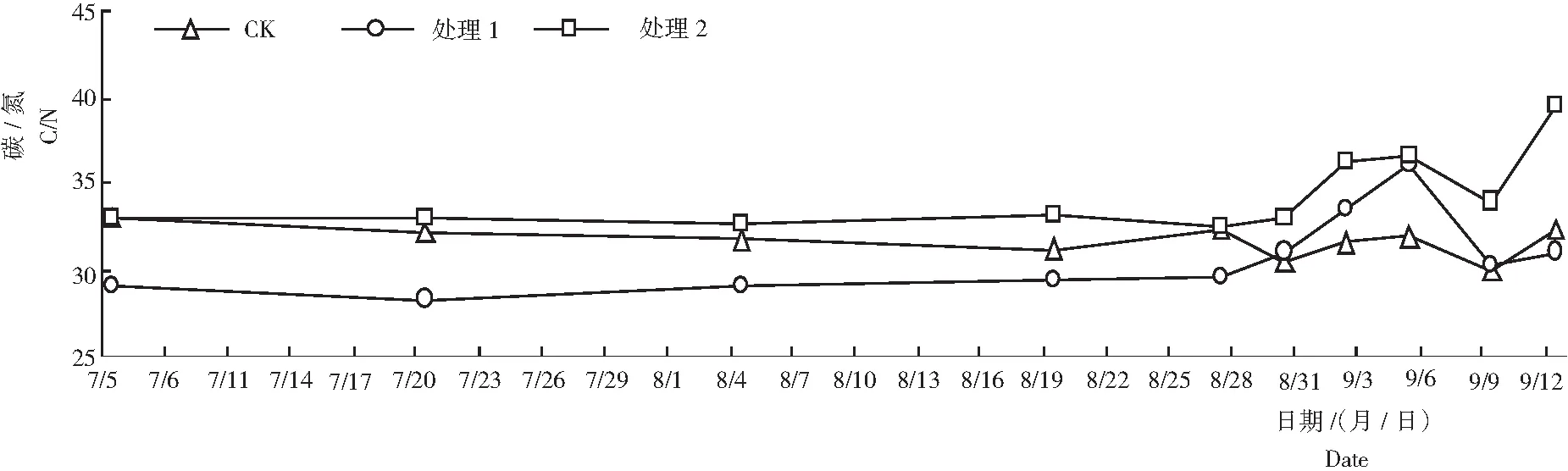
图4 不同处理各时期莲雾叶片的碳氮比
3 结论与讨论
1) 碳水化合物既是结构物质,又是能量的提供者,其积累与花芽分化密切相关[7]。试验表明,板栗叶片中高水平的可溶性糖明显促进其花芽分化,高含量的淀粉也有利于其花芽分化[8]。曾辉等[9]研究结果表明,枝条和叶片中可溶性糖的积累有利于澳洲坚果花芽的分化。叶片中可溶性糖的积累有利于草莓[10]、奈李[11]的花芽分化。吴月燕等[12]表明,葡萄花芽分化进度与可溶性糖呈显著正相关,叶片中的淀粉积累有利于花芽分化,淀粉含量与花芽分化呈显著正相关。郭金丽等[13]研究指出,苹果梨花芽生理分化期,成花短枝和叶片中淀粉积累快,形态分化期,成花短枝中淀粉大量积累;成花诱导并不一定要求很高的碳水化合物水平,而是强调其可利用性。本研究结果表明,莲雾通过遮阳和环割处理后,有利于叶片可溶性总糖和淀粉的积累,有大量花芽萌发的粉红种莲雾的积累量明显的高于几乎无花芽萌发的紫红种。可见,莲雾花芽分化前需要积累足够的可用性总糖和淀粉等碳水化合物,才能保证喷施催花药剂后,莲雾花芽分化的启动及进程。对于揭网喷施催花药剂后可溶性总糖的含量逐渐下降,则可能是由于莲雾花芽分化(粉红种)和叶芽大量萌发(紫红种)消耗所致,而淀粉含量上升可能是因为揭网后光合作用明显加强,合成量远大于消耗量所引起的。
2) 矿质元素是细胞的结构物质,是植物生命活动的调节者,在植物体内起电化学平衡的作用,是酶的活化剂,与碳水化合物代谢密切相关,与生长素和乙烯等激素有着密切的联系[14]。荔枝从花芽生理分化至开花前,叶片N、P、K含量逐渐上升,开花前达峰值,其中N变化较明显, P、K变化幅度较小,结论指出花芽分化初期大量的N、P、K积累可促进荔枝花芽分化。朱治强[15]研究指出,氮、磷、钾、钙、镁、铁、锌、铜、硼与香蕉植株的花芽分化关系密切。本研究结果表明,莲雾花芽是否分化与N的积累量的关系不密切,而与P、K的积累关系密切,较高的P含量和适当低的K含量可能有利于莲雾的花芽分化。
3) Klebs最早提出,只有当植物体内碳水化合物的积累比含氮化合物在数量上占优势时,植物才开始开花。Kraus和Kraybiil通过对番茄的研究提出了著名的碳氮比学说,即营养生长的强度和花芽的形成取决于碳水化合物与氮的数量之比[6,16-18]。该假说自提出后在多种果树上获得广泛支持[6,17-19]。本研究结果表明,莲雾花芽分化与C/N有密切关系,高C/N有利于其花芽分化,且较高C/N可能还需要维持一定的时间才能保证其花芽分化。
[1] 余 东,陈志峰,魏秀清,等.福建莲雾产期安排及调节关键技术[J].中国南方果树,2012,42(3):99-100.
[2] 韩 剑,罗仕争,李海明.海南莲雾的高产栽培技术[J].中国南方果树,2009,38(5):40-42.
[3] 黄邦贵,符 柏.防城港市莲雾生产现状及发展思路[J].调查研究,2009(3):20-21.
[4] 刘代兴,赵志平,李国华.印尼红莲雾产期调控试验[J].广东农业科学,2007(2):3-5.
[5] 周红玲,郑加协.莲雾产期调节技术及其在生产上的应用[J].实用技术,2010(3):53-55.
[6] 曹尚银,张秋明,吴顺.果树花芽分化机理研究进展[J].果树学报,2003,20(5):345-350.
[7] 曲 波,张 微,陈旭辉.植物花芽分化研究进展[J].中国农学通报,2010,26(24):109-114.
[8] 程 华,李琳玲,王少斌,等.板栗八月红花芽分化期相关营养物质含量的变化[J].湖北农业科学,2013,52(22):5502-5505.
[9] 曾 辉,杜丽清,邹明宏,等.澳洲坚果花芽分化期碳水化合物含量的变化动态[J].经济林研究,2013,31(2):65-70.
[10] 孙乃波,张志宏.草莓花芽分化过程中叶片碳水化合物和蛋白质含量的变化[J].安徽农业科学,2006,34(1):2328-2329.
[11] 钟晓红,罗先实,陈爱华.奈李花芽分化与体内主要代谢产物含量的关系[J].湖南农业大学学报,1999,25(1):31-35.
[12] 吴月燕,李培民,吴秋峰.葡萄叶片内碳水化合物及蛋白质代谢对花芽分化的影响[J].浙江万里学院学报,2002,15(4):54-57.
[13] 郭金丽,张玉兰.苹果梨花芽分化期蛋白质、淀粉代谢的研究[J].内蒙古农牧学院学报,1999,20(2):80-82.
[14] 许伟东.杨梅花芽生理分化与体内部分物质代谢关系研究[D].福州:福建农林大学,2008.
[15] 朱治强.香蕉植株的养分吸收积累与花芽分化的关系研究[D].儋州:华南热带农业大学,2003.
[16] 都荣庭.果树栽培学总论[M].3版.北京:中国农业出版社,2006:63-71.
[17] 吴邦良,夏春森,赵宗方,等.果树开花结实生理和调控技术[M].北京:科学技术出版社,1995:25-30.
[18] 李学柱,胡运权,孔 炎,等.甜橙大小年结果的研究——甜橙花芽分化及其生理代谢[J].园艺学报,1980,7(4):17-23.
[19] 李兴军,杨映根,郑文菊,等.果树花芽孕育的研究概况[J].植物学通报,2002,19(4):385-395.
(责任编辑: 杨 林)
Effects of Carbohydrate and Mineral Elements Content on Flower Bud Differentiation ofSyzygiumsamarangense
ZHAMG Lvping, CAI Yongqiang, JIN Jilin, CHEN Shouyi
(GuizhouFruitInstitute,Guiyang,Guizhou, 550006,China)
Purplish red and pinkS.amarangensewere treated by the combining method of spraying flower forcing agent, sun-shade and girdling to discuss the effect of carbohydrate and mineral elements content in leaves on flower bud differentiation ofS.amarangenseand provide the theoretical basis and reference for harvesting adjustment ofS.amarangense. Results: The enough accumulation of available total sugar and starch in leaves before flower bud differentiation ofS.amarangensecan guarantee the differentiation of flower buds sprayed with flower forcing agent. The flower bud differentiation is not closely related to nitrogen accumulation but is closely related to P and K accumulation. High P and proper low K content are beneficial to flower bud differentiation. The flower bud differentiation is closely related to C/N ratio, which indicates that maintaining high C/N ratio for a long time is beneficial to flower bud differentiation.
Syzygiumsamarangense; leaf; bud differentiation; carbohydrate; mineral element
2015-10-27; 2016-02-28修回
贵州省科学技术基金项目“莲雾花芽分化的生理机制”[黔科合J字(2011)2166]
张绿萍(1982-),女,副研究员,硕士,从事果树栽培生理与采后生理研究。E-mail:zlvping@163.com
1001-3601(2016)03-0128-0124-05
S667
A
园艺·中药材
Horticulture·CHinese Herbal Medicimes
猜你喜欢
小品文选刊(2018年13期)2018-11-14
小品文选刊(2018年7期)2018-07-05
意林(2018年10期)2018-05-09
农家科技中旬版(2017年5期)2017-07-14
科技与创新(2016年22期)2017-03-30
科技视界(2016年20期)2016-09-29
特产研究(2016年3期)2016-04-12
江苏农业科学(2014年11期)2015-01-15
天津农林科技(2012年3期)2012-05-14
