
黄土区长期封育草地优势物种生态位宽度与生态位重叠对不同干扰的响应特征
2016-01-28 03:35:58井光花程积民苏纪帅魏琳史晓晓金晶炜
草业学报 2015年9期
井光花,程积民,苏纪帅,魏琳,史晓晓,金晶炜
(1.中国科学院水利部水土保持研究所,陕西 杨凌 712100;2.中国科学院研究生院,北京100049;3.西北农林科技大学,陕西 杨凌 712100)
黄土区长期封育草地优势物种生态位宽度与生态位重叠对不同干扰的响应特征
井光花1,2,程积民1*,苏纪帅3,魏琳1,2,史晓晓3,金晶炜1
(1.中国科学院水利部水土保持研究所,陕西 杨凌 712100;2.中国科学院研究生院,北京100049;3.西北农林科技大学,陕西 杨凌 712100)
摘要:运用Levins生态位宽度指数和Pianka生态位重叠指数及物种丰富度,初步分析了长期封育草地优势种群生态位对放牧、火烧和施肥的响应特征。结果表明:在封育(对照组)、放牧、火烧和施肥干扰下,生态位宽度最大的物种依次是白颖苔草、中华小苦荬、白莲蒿和本氏针茅;建群种本氏针茅在不同干扰下生态位宽度表现为施肥(1.362)>火烧(1.244)>封育(1.088)>放牧(1.026),物种丰富度指数则依次为放牧>施肥>火烧>封育。所有物种和优势种的生态位重叠指数平均值均表现为施肥>封育>火烧>放牧,且在各干扰下,生态位重叠指数最大的种对依次为本氏针茅和白颖苔草、大针茅和白颖苔草、大针茅和白颖苔草以及中华小苦荬和硬质早熟禾。综上所述,对长期封育的草地进行适度的干扰可提高物种丰富度,改善种群的生态位配置和最大限度地利用环境资源。
关键词:生态位宽度; 生态位重叠;干扰类型;典型草原
Response of dominant population niche breadths and niche overlaps to various disturbance factors in typical steppe fenced grassland of China’s Loess Plateau region
JING Guang-Hua1,2, CHENG Ji-Min1*, SU Ji-Shuai3, WEI Lin1,2, SHI Xiao-Xiao3, JIN Jing-Wei1
1.InstituteofSoilandWaterConservation,ChineseAcademyofSciences&MinistryofWaterResources,Yangling712100,China;2.GraduateUniversityofChineseAcademyofSciences,Beijing100049,China;3.NorthwestAgricultural&ForestryUniversity,Yangling712100,China
Abstract:The Levins niche breadth and Pianka niche overlap indices were used to assess the response of plant population niches to grazing, burning and fertilization in permanently fenced grassland of the Loess Plateau region. Species with highest niche width in fenced (control group), grazed, burned and fertilized grasslands were Carex rigesaens, Ixeridium chinense, Artemisia sacrorum, and Stipa bungeana, respectively. The niche breadth of S. bungeana in different disturbance situations (ranked highest to lowest) was: fertilization (1.362)>burning (1.244)>fencing (1.088)>grazing (1.026). The richness index for the same four treatments ranked: grazing>fertilization>burning>fencing. The average values of niche overlap of all the species and dominant species ranked: fertilization>fencing>burning>grazing. The pairs of species with the highest niche overlap in fertilized, fenced, burned, and grazed grasslands were, respectively, S. bungeana and C. rigesaens, Stipa grandis and C. rigesaens, S. grandis and C. rigesaens, and I. chinense and Poa sphondylodes, respectively. These findings indicate that human disturbance can increase the species richness and improve plant population niche width to facilitate maximum resource use in permanently fenced grassland.
Key words:niche breadth; niche overlap; disturbances type; typical grassland
草地是陆地生态系统的主体类型之一,由于脆弱的生态环境和不合理的人类活动,成为对环境变化响应更为敏感的区域[1]。封育是当前退化草地恢复与重建的重要措施之一[2-3],在一定恢复年限内可提高物种丰富度[4],促进物种对资源利用的能力和生态位功能的发挥[5];但长时期封育会降低草地生产力和物种丰富度,不利于生态系统的稳定,故应对长期封育草地进行干扰。生态位是指种群在群落中的时空位置、功能和地位[6],能有效地反映物种对环境的适应和环境对物种的影响及其相互作用[7],深入研究长期封育草地优势种群生态位对不同干扰的响应有助于草地物种的更新和资源的利用。
目前,国内外学者基于生态位理论分析了森林和湿地优势种间的相互关系及其在群落中的位势[8-11],并在草地开展了大量研究。不同干扰和管理方式显著影响地上植物种类、盖度和高度等[12],而放牧、封育和施肥是草地生态系统管理和干扰类型[2,13-14],对草地生态系统和资源利用有重要的影响。韩范鸿等[15]研究了放牧梯度对内蒙古典型草原主要植物种群生态位的影响,董全民等[16]对放牧条件下高寒小嵩草(Kobresiaparva)草甸冷季草场种群生态位特征的研究表明物种的分布是既间断又连续;张晶晶和许冬梅[5]探讨了宁夏荒漠草原不同封育年限优势种群的生态位特征;陈子萱等[14]分析了施肥对玛曲高寒沙化草地主要植物种生态位的影响,认为施肥使禾草类生长旺盛,杂草类的优势地位逐渐失去;但多集中在退化草地,缺乏对长期封育草地的研究。同时,火烧作为影响草地物种丰富度和植被构成的重要因子[17],但火烧对种群生态位的响应也鲜有报道。
云雾山国家自然保护区采取封育措施恢复重建,成为我国西部最早、保护最完整的、黄土高原最具代表性以本氏针茅(Stipabungeana)为建群种的典型草原[18]。Jing等[4,19]认为封育可提高云雾山天然草地植被生物量、盖度、密度、丰富度及土壤养分含量(有机质、氮和磷),最佳封育年限为20年,封育期过长会抑制物种的更新和物种多样性;陈芙蓉等[20]和李媛等[21]研究表明施肥和火烧可进一步提高土壤质量和物种多样性,但缺乏长期封育草地优势种群生态位对放牧、火烧和施肥的响应研究。本研究以云雾山国家自然保护区为依托,以封育年限为20年的草地为参照,结合物种多样性和丰富度,分析优势种群生态位宽度和生态位重叠位对放牧、火烧和施肥的响应,以期为恢复草地的管理利用提供理论参考。
1材料与方法
1.1 研究区概况
云雾山国家自然保护区位于宁夏回族自治区固原市东北部45 km处(106°21′-106°27′ E,36°13′-36°19′ N),居于黄河流域的中上游地区,是黄土高原的中间地带,面积约为6660 hm2,海拔1800~2180 m。该区属于典型的温带大陆性半干旱季风气候,年均气温5℃,最热月为7月(约22~25℃),最冷月为1月(平均最低气温约-14℃),≥0℃的年积温为2370~2882℃。年降水量为400~450 mm,主要集中在7-9月,蒸发量为1330~1640 m,无霜期为4月中旬至9月底。地势南低北高,阳坡平缓,阴坡较陡,属于长梁缓坡丘陵区,土层深厚,土壤类型主要为黑垆土和黄绵土。保护区属于温带典型草原,其建群种和优势种为本氏针茅、大针茅(Stipagrandis)、百里香(Thymusmongolicus)和白莲蒿(Artemisiasacrorum)群落,伴生种以星毛委陵菜(Potentillaacaulis)、翼茎风毛菊(Saussureaalata)和白颖苔草(Carexaridula)为主,其中丛生禾本科植物本氏针茅分布范围最广。
1.2 试验设计
选取群落特征相近、地形一致和具有相同水热条件的封育20年样地,设置4种干扰处理,每个处理重复5次,面积为10 m×10 m,每个样方之间为2 m的分隔带,进行随机布设,实验时间为2011-2013年。1)封育:封育年限20年,无任何干扰活动,对照组;2)放牧,据产草量4500 kg/(hm2·年),每只羊需草3 kg/d,设置载畜率水平2.07羊单位/(hm2·半年),放牧绵羊数为4只,属于中度放牧。每年放牧30 d,分别在6,7和8月的中旬,放牧时间为16:00-18:00; 3)火烧:每年4月初,属于轻度火烧;4)施肥:每年5月中旬(返青时)施氮肥(尿素:N含量46%)和磷肥(P2O5含量12%),施肥量分别为21.74 g/(m2·年)和94.70 g/(m2·年)[22]。
1.3 研究方法
调查时间为2011、2012和2013年的9月(每年的生物量达到最大),在每个样方随机选取3个子样方(50 cm×50 cm),调查每个样方中出现的物种并测量高度、盖度、多度和频度等。高度由每种植物随机5株的平均值计算,盖度采用针刺法测定,多度是记录同一植物出现的个数(或丛数),地上生物量是将植物分种齐地面刈割,除去粘附的土壤、砾石、杂物后装袋,带回实验室烘干称重获取,频度为物种出现的样方数[20]。
1.4 评价指标和计算方法
1)重要值(important value)可确定各群落的优势种,能较客观地表达不同植物在群落中的作用与地位,其计算方法为[23]:
IV=(RH+RA+RC+RF+RB)/5
式中,RH为相对高度,RA为相对多度,RC为相对盖度,RF为相对频度,RB为相对生物量。

式中,Hi为某物种的高度(m),∑Hi为样方中所有植物高度之和(m);Ai为某物种的丰富度,∑Ai为所有物种丰富度之和;Ci为某物种的盖度(%),∑Ci为所有物种盖度之和(%);Fi为某物种的频度,∑Fi为所有物种的总频度;Bi为某物种的地上生物量(g/m2),∑Bi为所有物种的地上生物量(g/m2)。
2)生态位宽度(nichebreadth) 采用Levins生态位宽度计算公式[5]:

式中,Bi是种i的生态位宽度,Pij是种i对第j个资源的利用占它对全部资源利用的频度,Pij=nij/Nij,而Nij=∑nij, nij为物种i在资源梯度级j的重要值,本研究用样方代表资源位,r为样方数(r=15)。
3)生态位重叠(nicheoverlap) 采用Pianka生态位重叠指数,计算公式为[24]:

式中,Qik为物种i与物种k的重叠度指数,其他符号含义同上。
样地全部物种间生态位重叠值的总平均值=样地内全部物种间生态位重叠值总数/总种对数。
4)物种丰富度Patrick指数(R) R=S,S 为样地中的物种数
5)物种多样性(speciesdiversity) 采用Shannon-Wiener指数(H′),公式为[25]:
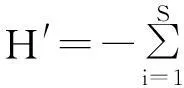
式中,S为样地中的物种数,Pi为样地中第i种的相对重要值。
2结果与分析
2.1 生态位宽度
不同干扰草地生态位宽度见表1,可知同一干扰下云雾山草地不同物种生态位宽度不同,不同干扰下相同物种的生态位宽度也不尽相同。从表1可以看出,封育草地,白颖苔草和大针茅、硬质早熟禾、本氏针茅的生态位宽度较大,菊科、豆科和唇形科植物(除个别植物外)的生态位宽度相对较低,说明莎草科和禾本科植物对资源的综合利用能力较强,其他科植物相对较弱。对封育草地放牧后,牲畜喜食物种大针茅和白颖苔草生态位宽度分别骤减了59.67%和25.43%;牲畜不喜食物种中华小苦荬出现并快速生长,成为放牧草地的优势种之一,星毛委陵菜和裂叶堇菜生态位宽度增幅也较大;但本氏针茅、扁穗冰草和硬质早熟禾优势地位并未受到显著影响。
表1显示,封育草地在遭遇火烧后,本氏针茅和白莲蒿生态位宽度分别增加了14.34%和45.43%,表明火烧促进了本氏针茅和白莲蒿生态位的拓宽;大针茅、硬质早熟禾、阿尔泰狗娃花、二裂委陵菜和白颖苔草的生态位宽度虽有所降低,但其值仍较高,说明其对火烧环境的适应能力较强;同时出现了猪毛蒿和急折百蕊草新物种。对封育草地施肥后,多数植物生态位宽度增加,本氏针茅生态位宽度跃居首位,增幅为21.88%,茅香变化最大,由0增加到0.853;大针茅和白颖苔草生态位宽度虽降低,但降幅较小,仍处于优势地位;多数菊科植物生态位宽度不同程度地增加,但阿尔泰狗娃花适应能力较差,其生态位宽度降低,由1.084到0。
表1还表明,建群种本氏针茅在火烧和施肥干扰后优势地位较为突显,说明在长期封育的草地进行干扰可促进建群种的生长,提高其适应环境和利用资源的能力。大针茅和白颖苔草在封育20年的生态位宽度值最大,而干扰不同程度上抑制了植物的发育和生长。综上所述,放牧整体上抑制了多数禾本科植物生态位的拓宽,施肥促进了禾本科植物的生长,火烧提高了菊科植物白莲蒿和火绒草的生态位宽度。
注:“-”表示未出现此物种。
Note: “-” represents that there is no this species.
2.2 生态重叠
根据表1,选取生态位宽度较大且在各处理均出现的物种1、2、3、4、6、7、8、16、18、22、25和27分析Pianka生态位重叠指数(表2和表3)。由表2可知,封育草地大针茅与白颖苔草的生态位重叠值最大,为0.956,表明大针茅和白颖苔草对资源的需求具有很强的相似性。白颖苔草与硬质早熟禾、大针茅与硬质早熟禾、硬质早熟禾和白莲蒿次之,其生态位重叠值分别为0.878、0.866和0.858。百里香与其他物种的生态重叠指数普遍偏低,说明百里香和其他物种对环境资源生态学要求存在差异。表2还表明,对封育草地放牧后,裂叶堇菜与青海苜蓿和白颖苔草的生态重叠指数稍有增加,除本氏针茅和扁穗冰草的生态重叠指数为0.803,整体上各优势种对生态位重叠值普遍偏低,说明放牧干扰存在种间对资源分化的作用。
注:第1列与第1行编号所代表物种同表1中各物种编号及相应物种。下同。
Note:The species which represented by numbers in the first column and the first row of the table are same with the species which was ordered with an identical number in Table 1. The same below.
表3显示,相比封育草地,火烧样地大部分种对生态位重叠指数有所降低,但本氏针茅和扁穗冰草、本氏针茅和硬质早熟禾、白莲蒿和白颖苔草的生态重叠指数呈增加趋势。样地生态位重叠值最大的是白颖苔草和大针茅,说明它们生物学特性较为相似,或在同一空间内生态需求互补性较强。百里香与扁穗冰草、裂叶堇菜和青海苜蓿的生态位重叠指数降为0,表明它们对资源利用相似程度较小。从表3也可看出,施肥样地各种对生态位重叠值整体偏高,大于0.8的高达15对。生态位宽度较大的物种间生态位重叠较高,如本氏针茅与白颖苔草和硬质早熟禾,其生态位重叠值分别为0.903和0.902,说明物种对环境资源具有相似的生态学要求。最小的生态位重叠值是阿尔泰狗娃花与二裂委陵菜,仅为0。
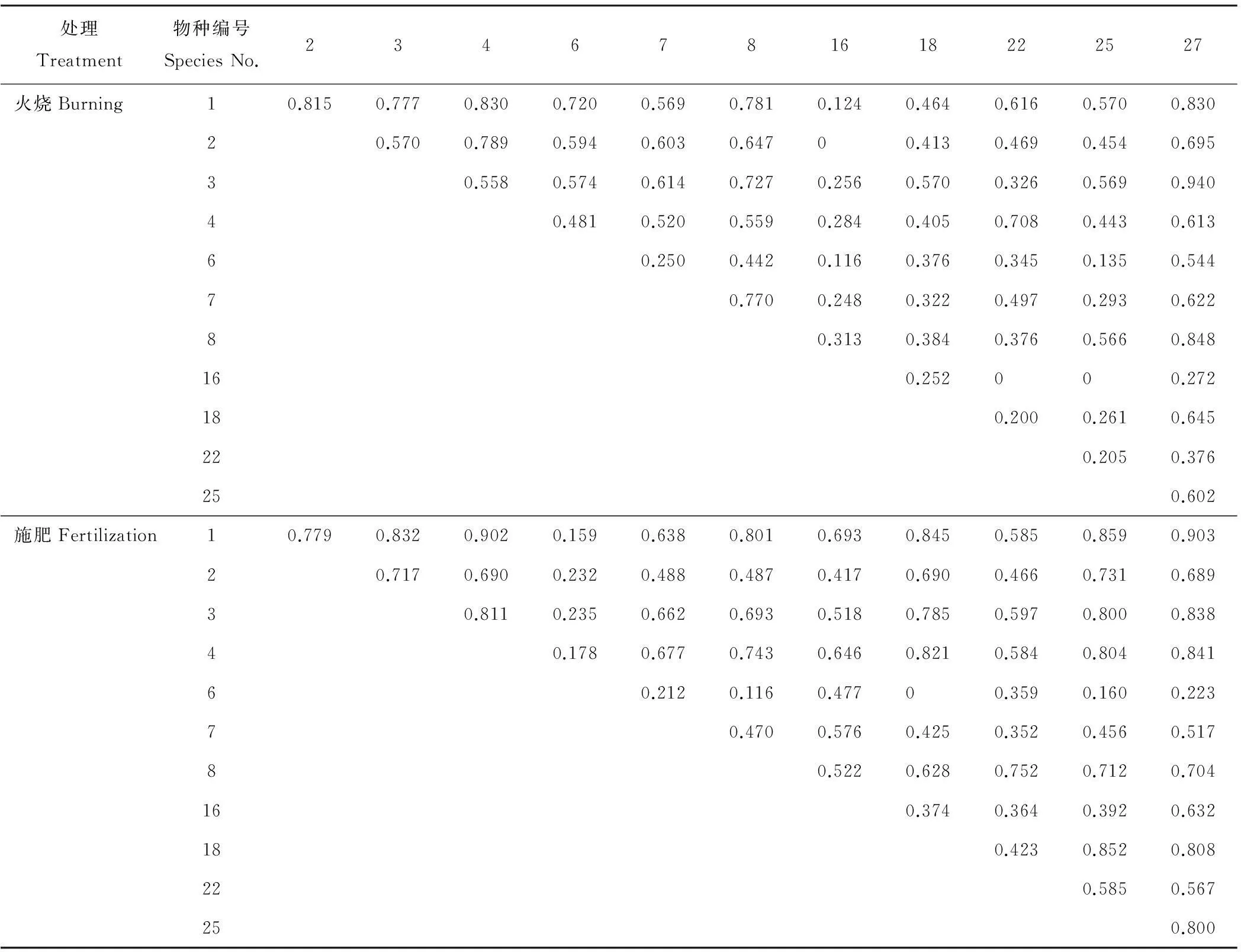
表3 火烧和施肥草地优势种的生态位重叠
综合表1~表3可知,各干扰草地多数生态位宽度较大的物种和其他物种有较高的重叠值,也有少数生态位宽度较大的物种未出现较高的生态位重叠指数,如火烧本氏针茅生态位宽度最大,但白颖苔草与大针茅的生态位重叠指数最高。表2和表3还表明,不同干扰方式对物种间生态位重叠指数的影响存在一定的差异,以本氏针茅、扁穗冰草和硬质早熟禾为例,本氏针茅和硬质早熟禾的生态位重叠值表现为施肥(0.902)>火烧(0.830)>封育(0.675)>放牧(0.523),而扁穗冰草和本氏针茅、硬质早熟禾的最高生态位重叠指数均出现在火烧样地,最低的重叠指数则发生在封育样地。
表4表明,所有物种和优势种的生态位重叠指数平均值均表现为施肥>封育>火烧>放牧,说明在封育草地上施肥提高了资源利用的共同性,而火烧的发生消除了封育草地的枯落物,为物种的生存提供了空间,物种间的竞争关系得到了缓和;在牲畜的啃食和践踏下,放牧促使生态位分化,降低了资源的利用和物种间的竞争。
2.3 物种多样性
对长期封育草地进行干扰后,物种多样性和丰富度均发生了显著变化,如图1。与封育草地相比,放牧的Shannon-Weiner指数增加了15.90%,火烧和施肥的Shannon-Weiner指数分别降低了14.03%和21.19%,说明放牧可提高物种多样性,而火烧和施肥抑制物种多样性。对封育草地进行放牧、火烧和施肥后,Patrick指数均呈现不同程度地增加,且放牧草地丰富度最高。

表4 不同干扰草地种群间生态位重叠指数的平均值

图1 不同干扰草地物种多样性指数和丰富度指数Fig.1 Species diversity and richness in different disturbed grassland 不同字母表示差异显著(P<0.05)。Different letters mean significant difference among disturbance type (P<0.05).
3讨论
生态位是指一个种群在生态系统中,在时间空间上所占据的位置及其与相关种群之间的功能关系与作用[6]。在无别的竞争物种存在时所占有的生态位叫基础生态位,只是一种理论上的生态位;但在自然界中,物种间存在捕食和竞争,任何物种占有的实际生态位比理论上的生态位小一些,称作实现生态位[26]。而生态位宽度和生态位重叠被认为是影响物种多样性及群落结构的决定因素和描述各物种的实现生态位及其相互关系的重要指标[27-28],都在一定程度上反映了植物自身的特性和对环境的适应性,二者既有联系,又有区别。生态位宽度是反映物种对环境资源利用状况的尺度,而生态位重叠则反映了种群之间对资源利用的相似程度和竞争关系[29]。研究认为,生态位宽度越大,对环境的适应能力越强,同时伴随着较高的生态位重叠,生态位宽度较小的物种则因对资源的利用能力较弱,分布较窄而与其他物种间有较低的生态位重叠[30];但由于资源存在高度的空间异质性或者不同演替阶段的竞争特点存在差异[8,31],较高的生态位重叠值也会出现在生态位宽度较小的物种间。
种群生态位除植物自身特性制约外,还受海拔高度、土壤湿度、人为干扰等环境因子的影响[9]。目前,放牧、施肥和火烧是人类在草地生态系统管理利用实践中的主要干扰类型,对生态系统过程产生重要影响。放牧是草地常见且高度复杂的干扰方式,牲畜的选择性采食和践踏改变了草地种群分布,从而影响草地种群生态位特征[32]。本试验放牧干扰的物种多样性和丰富度高于其他干扰方式,符合中度干扰理论,即中等干扰条件下,物种多样性最大[33]。由于牲畜选择性的采食抑制了优势种(本氏针茅)和优质牧草(大针茅)的生长,而耐牧草种、耐践踏草种和牲畜不喜食的杂类草明显增加,这为竞争力较弱物种的生存拓宽了空间[20]。本研究中,牲畜不喜食草种中华小苦荬的生态位宽度最大(1.067),耐牧草种白莲蒿和扁穗冰草以及耐践踏草种星毛委陵菜的生态位宽度较大,而牲畜喜食草种大针茅的生态位宽度仅为0.467。放牧干扰存在种间对资源分化的作用,从而引起生态位重叠程度降低[13],故放牧干扰的所有物种生态位重叠指数平均值低于其他干扰方式。
火烧是在时空上对种群的功能性状、群落组成和结构及生态系统功能有着显著影响的干扰因子[34]。本研究火烧发生在封育20年草地的生长季初期,消除了大量地上立枯物和凋落物,为急折百蕊草、猪毛蒿和翻白草等竞争较弱的物种提供了生存空间;同时,地表接受到的光照强度和土壤温度的增加刺激了种子的萌发,提高了物种的丰富度。由于本氏针茅耐火烧能力强,草地返青后能较快生长发育,本氏针茅的生态位宽度并未受到影响,反而增加。白莲蒿属于半灌木,根蘖性强,且分泌出的化感物质抑制其他物种的生长[35],故白莲蒿生态位宽度较大。但所有物种生态位重叠指数平均值在火烧后有所降低,可能是因为火烧消除的立枯物和凋落物,为物种提供了较多的光照资源[20],消减了物种间竞争。虽生态位宽度最大的是本氏针茅,但生态位重叠指数较大的是白颖苔草和大针茅、大针茅和扁穗冰草及白莲蒿和白颖苔草,可能是由于火烧引起了环境空间异质性。
施肥是提高草地生产力的有效手段,可引起植物群落多样性格局的变化,对草地的恢复和生态系统的稳定存在一定影响[36-37]。研究认为,施肥会降低物种丰富度[38],而本研究由于施肥时间较短[20],物种丰富度并未降低。对草地施肥后,植物得到较好的生长,相比之下喜肥的禾草植物具有更快的生长率,故施肥草地的多数物种生态位宽度拓宽,如本氏针茅生态位宽度高达1.362;但群落中优势种的旺盛生长,群落的密闭度增加,使群落下层不耐阴物种或者个体较小物种在群落中减少或死亡,如阿尔泰狗娃花生态位宽度由封育草地的1.084骤减为0。施肥样地物种间生态位重叠指数普遍偏高,如针茅类(本氏针茅和大针茅)和其他物种间的生态位重叠指数均大于0.6,是因为资源的增加改变了群落内物种的相对竞争力,使草地优势种地上、地下竞争能力增强[39]。同时,生态位宽度较大的物种间生态位重叠指数较大,说明施肥后共同利用的生态资源越多,相同生活型的物种竞争就越激烈[14]。
4结论
1)对云雾山长期封育草地放牧、火烧和放牧后,草地优势种依然是本氏针茅,且在施肥草地中生态位宽度最大(1.362)。放牧增加了耐牧草种的生态位宽度(如中华小苦荬),火烧显著提高了白莲蒿的生态位宽度(增幅为45.43%),施肥则总体上促进了禾本科的生长。
2)在各干扰中,生态位重叠指数最大的种对均未出现在生态位宽度较大的物种间。生态位重叠指数平均值均表现为施肥>封育>火烧>放牧,各干扰下生态位重叠指数最大的种对依次为本氏针茅和白颖苔草(0.913)、大针茅和白颖苔草(0.956)、大针茅和白颖苔草(0.940)以及中华小苦荬和硬质早熟禾(0.822)。
3)各干扰可提高物种丰富度,但放牧促进了物种多样性,而施肥和火烧抑制了物种多样性。
References:
[1]Zhou P, Liu G B, Xue S. Review of soil respiration and the impact factors on grassland ecosystem. Acta Prataculturae Sinica, 2009, 18(2): 184-193.
[2]Shan Q L, Xu Z, Ning F,etal. Influence of exclosure year on community structure and species diversity on a typical steppe. Acta Prataculturae Sinica, 2008, 17(6): 1-8.
[3]Wang C Y, Zhang J J, Lv Y L,etal. Effects of long-term grazing exclusion on soil organic carbon fractions in the grassland of Inner Mongolia. Acta Prataculturae Sinica, 2014, 23(5):31-39.
[4]Jing Z B, Cheng J M, Chen A. Assessment of vegetative ecological characteristics and the succession process during three decades of grazing exclusion in a continental steppe grassland. Ecological Engineering, 2013, 57:162-169.
[5]Zhang J J, Xu D M. Niche characteristics of dominant plant populations in desert steppe of Ningxia with different enclosure times. Acta Agrestia Sinica, 2013, 21(1):73-78.
[6]Dao R N, Wang S S, Li J Q,etal. Plant niche changes of the rehabilitating water and soil conservation vegetations in Taizi watershed. Journal of Northeast Forestry University, 2014, 42(1): 37-41, 70.
[7]Zhong Y, Zhang J, Liu Q B,etal. Niche characteristics of main herbage populations inEucalyptusgrandisplantation. Acta Prataculturae Sinica, 2010, 19(4): 16-21.
[8]Hu Z H, Qian H Y, Yu M J. The niche of dominant species populations inCastanopsiseyreiforest in Gutian Mountain National Nature Reserve. Acta Ecologica Sinica, 2009, 29(7): 3670-3677.
[9]Li B, Li S Q, Zhang J T. A study on niches of dominant species of subalpine meadow in the Yunding Mountain. Acta Prataculturae Sinica, 2010, 19(1): 6-13.
[10]Li F, Xie Y H, Chen X S,etal. Composition of aquatic plants and their niche characteristics in wetlands of the Yellow River Delta. Acta Ecologica Sinica, 2009, 29(11): 6257-6265.
[11]Germain R M, Gibert B. Hidden responses to environmental variation: maternal effects reveal species niche dimensions. Ecology Letters, 2014, 17(6): 662-669.
[12]Zhang Y X, Yao T, Wang G J,etal. Characteristics of vegetation and soil inorganic nitrogen concentrations under different disturbed habitats in a weak alpine ecosystem. Acta Prataculturae Sinica, 2014, 23(4): 245-252.
[13]Zhang T, Weng Y, Yao F J,etal. Effect of grazing intensity on ecological stoichiometry ofDeyeuxiaangustifoliaand meadow soil. Acta Prataculturae Sinica, 2014, 23(2):20-28.
[14]Chen Z X, Tian F P, Zheng Y. Effects of fertilization on niche of main plant species in Maqu alpine desertification meadow. Acta Agrestia Sinica, 2011, 19(4): 884-888.
[15]Han F H, Wang S P, Chen Z Z. Response of the major plant populations of Inner Mongolia typical steppe to grazing rate based on niche considerations. Acta Agrestia Sinica, 1999, 7(3): 204-210.
[16]Dong Q M, Zhao X Q, Ma Y S,etal. Effects of grazing on the major plant populations niche in alpineKobreciaparvameadow cold-seasonal pastureland. Chinese Journal of Grassland, 2006, 28(6): 10-17.
[17]Wu G L, Zhao L P, Wang D,etal. Effects of time-since-fire on vegetation composition and structures in semi-arid perennial grassland on the Loess Plateau, China. Clean-Soil, Air, Water, 2014, 42(1): 98-103.
[18]Chen F R, Cheng J M, Liu W,etal. Effects of different disturbances on soil physical and chemical properties in the typical grassland of Loess Region. Journal of Soil and Water Conservation, 2012, 26(2): 105-111.
[19]Jing Z B, Cheng J M, Su J S,etal. Changes in plant community composition and soil properties under 3-decade grazing exclusion in semiarid grassland. Ecological Engineering, 2014, 64: 171-178.
[20]Chen F R, Cheng J M, Liu W,etal. Effects of different disturbances on diversity and biomass of communities in the typical steppe of loess region. Acta Ecologica Sinica, 2013, 33(9): 2856-2866.
[21]Li Y, Cheng J M, Wei L,etal. Changes of soil chemical properties after different burning years in typical steppe of Yunwun Mountains. Acta Ecologica Sinica, 2013, 33(7): 2131-2138.
[22]Jia X X, Shao M A, Wei X R. Soil CO2efflux in response to the addition of water and fertilizer in temperate semiarid grassland in northern China. Plant Soil, 2013, 373: 125-141.
[23]Li R, Zhang K B, Liu Y F,etal. Plant community spatial distribution pattern of wetland ecosystem in a semi-arid region of northwestern China. Journal of Beijing Forestry University, 2008, 30(1): 6-13.
[24]Tang J M, Ai X R, Yi Y M,etal. Niche dynamics during restoration process for the dominant tree species in montane mixed evergreen and deciduous broadleaved forests at Mulinzi of southwest Hubei. Acta Ecologica Sinica, 2012, 32(20): 6334-6342.
[25]Niu K, Choler P, Bello F,etal. Fertilization decreases species diversity but increases functional diversity: A three-year experiment in a Tibetan alpine meadow. Agriculture Ecosystem & Environment, 2014, 182: 106-112.
[26]Yu S X, Orloci L. On the implications of fundamental, realized niche and niche center. Acta Scientiarum Naturalium Universitatis Sunyatseni, 1993, 32(4): 69-80.
[27]Li M, Jiang D M, Toshio O,etal. Niche characteristic of herbages in artificial sand-fixing communities in Horqin sandy land. Pratacultural Science, 2009, 26(8): 10-16.
[28]Zhang Z H, Liang S C, Hu G. Niche characteristics of dominant populations inCinnamomumburmanniicommunity on Karst Hills of Guilin. Forest Research, 2009, 22(1): 63-68.
[29]Silvertown J W. Plants in limestone pavements: tests of species interaction and niche separation. Journal of Ecology, 1983, 71: 819-828.
[30]Shi Z M, Cheng R M, Liu S R. Niche characteristics of plant populations in deciduous broad-leaved forest in Baotianman. Chinese Journal of Applied Ecology, 1999, 10(3): 265-269.
[31]Chen Z X, Tian F P, Niu J Y,etal. Effects of fertilization on niche of main plant species in Maqu alpine desertification meadow. Acta Agrestia Sinica, 2007, 15(6): 525-530.
[32]Wang R Z. The niche breadths and niche overlaps of main plant populations inLeymuschinesegrassland for grazing. Acta Phytoecologica Sinica, 1997, 21(4): 304-311.
[33]Wen L, Dong S K, Zhu L,etal. The effect of natural factors and disturbance intensity on spatial heterogeneity of plant diversity in alpine meadow. Acta Ecologica Sinica, 2011, 31(7): 1844-1854.
[34]Tang C Q, He L Y, Su W H,etal. Regeneration, recovery and succession of aPinusyunnanensiscommunity five years after a mega-fire in central Yunnan, China. Forest Ecology and Management, 2013, 294: 188-196.
[35]Wang H, Xie Y S, Cheng J M,etal. Allelopathic effects ofArtemisiasacrorumpopulation in typical steppe based on niche theory. Chinese Journal of Applied Ecology, 2012, 23(3): 673-678.
[36]Chen Y M, Li Z Z, Du G Z. Effects of fertilization on plant diversity and economic herbage groups in alpine meadow. Acta Botanica Boreali-Occidentalia Sinica, 2004, 24(3): 424-429.
[37]Wang H L, Niu J Y, Zheng H P,etal. Research on characteristics of plant niche and effects of fertilization on plant productivity in a Maqu Alpine desertified meadow. Acta Prataculturae Sinica, 2008, 17(6): 18-24.
[38]Niu K C, Luo Y J, Choler P,etal. The role of biomass allocation strategy on diversity loss due to fertilization. Basic and Applied Ecology, 2008, 9(5): 485-493.
[39]Gilliam F S. Response of the herbaceous layer of forest ecosystems to excess nitrogen deposition. Journal of Ecology, 2006, 94: 1176-1191.
参考文献:
[1]周萍, 刘国彬, 薛萐. 草地生态系统土壤呼吸及其影响因素研究进展. 草业学报, 2009, 18(2): 184-193.
[2]单贵莲, 徐柱, 宁发, 等. 围封年限对典型草原群落结构及物种多样性的影响. 草业学报, 2008, 17(6): 1-8.
[3]王春燕, 张晋京, 吕瑜良, 等. 长期封育对内蒙古羊草草地土壤有机碳组分的影响. 草业学报, 2014, 23(5):31-39.
[5]张晶晶, 许冬梅. 宁夏荒漠草原不同封育年限优势种群的生态位特征. 草地学报, 2013, 21(1):73-78.
[6]道如娜, 王树森, 李建强, 等. 太子河流域水源涵养林恢复植被的植物生态位变化. 东北林业大学学报, 2014, 42(1): 37-41, 70.
[7]钟宇, 张健, 刘泉波, 等. 巨桉人工林草本层主要种群的生态位分析. 草业学报, 2010, 19(4): 16-21.
[8]胡正华, 钱海源, 于明坚. 古田山国家级自然保护区甜槠林优势种群生态位. 生态学报, 2009, 29(7): 3670-3677.
[9]李斌, 李素清, 张金屯. 云顶山亚高山草甸优势种群生态位研究. 草业学报, 2010, 19(1): 6-13.
[10]李峰, 谢永宏, 陈心胜, 等. 黄河三角洲湿地水生植物组成及生态位.生态学报, 2009, 29(11): 6257-6265.
[12]张玉霞, 姚拓, 王国基, 等. 高寒生态脆弱区不同扰动生境草地植被及土壤无机氮变化特征. 草业学报, 2014, 23(4): 245-252.
[13]张婷, 翁月, 姚凤娇, 等. 放牧强度对草甸植物小叶章及土壤化学计量比的影响. 草业学报, 2014, 23(2):20-28.
[14]陈子萱, 田福平, 郑阳. 施肥对玛曲高寒沙化草地主要植物种生态位的影响. 草地学报, 2011, 19(4): 884-888.
[15]韩范鸿, 汪诗平, 陈佐忠. 以放牧率梯度研究内蒙古典型草原主要植物种群的生态位. 草地学报, 1999, 7(3): 204-210.
[16]董全民, 赵新全, 马玉寿, 等. 放牧对高寒小嵩草草甸冷季草场主要植物种群生态位的影响. 中国草地学报, 2006, 28(6): 10-17.
[18]陈芙蓉, 程积民, 刘伟, 等. 不同干扰对黄土区典型草原土壤理化性质的影响. 水土保持学报, 2012, 26(2): 105-111.
[20]陈芙蓉, 程积民, 刘伟, 等. 不同干扰对黄土区典型草原物种多样性和生物量的影响. 生态学报, 2013, 33(9): 2856-2866.
[21]李媛, 程积民, 魏琳, 等. 云雾山典型草原火烧不同恢复年限土壤化学性质变化. 生态学报, 2013, 33(7): 2131-2138.
[23]李瑞, 张克斌, 刘云芳, 等. 西北半干旱区湿地生态系统植物群落空间分布特征研究. 北京林业大学学报, 2008, 30(1): 6-13.
[24]汤景明, 艾训儒, 易咏梅, 等. 鄂西南木林子常绿落叶阔叶混交林恢复过程中优势树种生态位动态. 生态学报, 2012, 32(20): 6334-6342.
[26]余世孝, 奥罗西L. 基础与实现生态位及其中心点的涵义与测度. 中山大学学报(自然科学版), 1993, 32(4): 69-80.
[27]李明, 蒋德明, 押田敏雄, 等. 科尔沁沙地人工固沙群落草本植物种群生态位特征. 草业科学, 2009, 26(8): 10-16.
[28]张忠华, 梁士楚, 胡刚. 桂林岩溶石山阴香群落主要种群生态位研究. 林业科学研究, 2009, 22(1): 63-68.
[30]史作民, 程瑞梅, 刘世荣. 宝天曼落叶阔叶林种群生态位特征. 应用生态学报, 1999, 10(3): 265-269.
[31]陈子萱, 田福平, 牛俊义, 等. 施肥对玛曲高寒沙化草地主要植物种生态位的影响. 草地学报, 2007, 15(6): 525-530.
[32]王仁忠. 放牧影响下羊草草地主要植物种群生态位宽度与生态位重叠的研究. 植物生态学报, 1997, 21(4): 304-311.
[33]温璐, 董世魁, 朱磊, 等. 环境因子和干扰强度对高寒草甸植物多样性空间分异的影响. 生态学报, 2011, 31(7): 1844-1854.
[35]王辉, 谢永生, 程积民, 等. 基于生态位理论的典型草原铁杆蒿种群化感作用. 应用生态学报, 2012, 23(3): 673-678.
[36]陈亚明, 李自珍, 杜国祯. 施肥对高寒草甸植物多样性和经济类群的影响, 西北植物学报, 2004, 24(3): 424-429.
[37]王鹤龄, 牛俊义, 郑华平, 等. 玛曲高寒沙化草地生态位特征及其施肥改良研究. 草业学报, 2008, 17(6): 18-24.
井光花,程积民,苏纪帅,魏琳,史晓晓,金晶炜. 黄土区长期封育草地优势物种生态位宽度与生态位重叠对不同干扰的响应特征. 草业学报, 2015, 24(9): 43-52.
JING Guang-Hua, CHENG Ji-Min, SU Ji-Shuai, WEI Lin, SHI Xiao-Xiao, JIN Jing-Wei. Response of dominant population niche breadths and niche overlaps to various disturbance factors in typical steppe fenced grassland of China’s Loess Plateau region. Acta Prataculturae Sinica, 2015, 24(9): 43-52.
通讯作者*Corresponding author. E-mail: gyzcjm@ms.iswc.ac.cn
作者简介:井光花(1987-),女,山东济宁人,在读博士。E-mail:jingguanghuajgh@126.com
基金项目:国家自然科学基金重点项目“气候变化背景下黄土高原土地利用影响径流的空间尺度效应”(41230852),中国科学院战略性先导科技专项“应对气候变化的碳收支认证及相关问题”(XDA05050202)和农业部“现代农业产业技术体系建设专项”(CARS-35-40)资助。
收稿日期:2014-10-08;改回日期:2014-12-25
DOI:10.11686/cyxb2014410http://cyxb.lzu.edu.cn
