
沱江浮游植物群落特征及水质评价*
2016-01-15 08:21:25谢碧文齐泽民杨品红
海洋与湖沼 2016年4期
陶 敏 谢碧文 齐泽民 杨品红 李 斌①
(1. 内江师范学院生命科学学院 长江上游鱼类资源保护与利用四川省重点实验室 内江 641112; 2. 湖南文理学院 水产高效健康生产湖南省协同创新中心 环洞庭湖水产健康养殖及加工湖南省重点实验室 动物学湖南省高校重点实验室 常德415000)
水质是水生态系统健康的重要指标, 进行水质评价是水体开发利用、保护与生态恢复的前提与基础(焦珂伟等, 2015)。早期的水质评价多针对理化指标(Bozzetti et al, 2004)。然而由于环境与生物之间的复杂相互作用, 仅仅用理化指标无法完整体现水体的健康状况。作为水域的初级生产者, 浮游植物是整个水生态系统物质循环和能量流动的基础(王爱爱等,2014), 在维持水生态系统平衡中起着重要作用。浮游植物群落结构对水环境条件变化敏感, 其种类组成和分布格局对环境变化具有重要指示作用, 已被广泛应用于水质评价中(况琪军等, 2005; 王爱爱等,2014; 苟婷等, 2015; 杨敏等, 2015)。
沱江为长江上游重要支流, 流域内人口密度高,工农业发达, 污染严重, 富营养化程度在四川省主要河流中最高(刘佳等, 2012)。2003年的特大水污染事件(孙莉等, 2005)引起了学者们对沱江水质的密切关注, 从各种理化指标的角度对沱江水质进行评价(谢贤健等, 2009; 肖玖金等, 2011; 周实际等, 2011; 刘建平, 2013; 陈雨艳等, 2015; 周开锡等, 2015), 而生物学监测方面的研究仍停留在20世纪80、90年代(李进, 1986; 杨昌述, 1987)。本研究对沱江中下游10个断面开展采样调查, 从理化指标和浮游植物两个角度出发进行水质评价, 为沱江水体健康状况提供重要的基础数据, 也为水体的利用及河流生态修复提供参考资料。
1 材料与方法
1.1 样品采集与分析
1.1.1 采样点布设 根据研究江段周边环境特点,在城市区(莲花山、资阳一桥、西林渡口、沱桥)、工业区或建筑工地附近(麻柳坝工业园、资州大桥)、城郊上游或乡镇(顺河场镇、五里店水电站、银山镇、二水厂)布设采样断面 10个(图 1)。各采样点环境描述参考陶敏等(2016)。

图1 采样点分布图Fig.1 Map of sampling sites
1.1.2 样品采集 于2013年8月(丰水期)与11月(枯水期)在研究江段进行采样和调查。表层水样以辛德勒采水器(Schindler-Patalas trap)采集。水化学及叶绿素样品直接装入干净的样品瓶密封, 冷藏带回实验室。同时将1L浮游植物定量样品装入样品瓶并立即加入鲁哥氏试剂(Lugol’s reagent) 现场固定。浮游植物定性样品用 25号浮游生物网在水面下作“∞”字型捞取并过滤收集, 加入4%甲醛现场固定。
1.1.3 浮游植物样品的处理与分析 将浮游植物定量样品在浮游生物沉淀器中避光静置沉降 48h, 以虹吸法去除上层清液后, 浓缩定容至 50mL, 加入数滴甲醛溶液保存。计数方法参照章宗涉等(1991), 种类鉴定参考胡鸿钧等(2006)。
1.1.4 理化指标的分析测定 使用塞氏盘(Secchi disc)、便携式溶解氧测量仪(YSI-550A)、电导率仪(Mettler Toledo SevenGo SG3)等设备对研究区域透明度(Secchi Depth, SD)、水温、pH、溶解氧(DO)和电导率(Conductivity)等进行现场测定。总氮(TN)、氨氮(NH4-N)、硝酸盐氮(NO3-N)、总磷(TP)、正磷酸盐(PO4-P)、高锰酸盐指数(CODMn)、叶绿素a(Chl a)含量在实验室中按照《水和废水监测分析方法》(国家环境保护总局, 2002)进行分析。
1.2 评价方法
1.2.1 优势种与优势度 浮游植物优势种依据各种的优势度指数(Y)值进行鉴别(刘成士等, 2015):

式中, Pi为第i种浮游植物丰度与样品中浮游植物总丰度的比值(ni/N), fi为第i种浮游植物在各采样点出现的频度。Y≥0.02的物种为优势种。
1.2.2 综合营养状态指数 以总氮(TN)、总磷(TP)、高锰酸盐指数(CODMn)、叶绿素a(Chl a)和透明度(SD)为评价指标, 采用湖泊(水库)富营养化评价方法及分级技术规定(中国环境监测总站, 2001)中的综合营养状态指数 TLI(∑)评价方法与标准对沱江营养状态进行分析。

式中, Wj为第 j种参数的营养状态指数的相关权重;TLI(j)为第 j种参数的营养状态指数。TLI(∑)≤30时为贫营养状态; 30<TLI(∑)≤50为中营养状态;TLI(∑)>50 则为富营养化状态, 其中 50<TLI(∑)≤60为轻度富营养化, 60<TLI(∑)≤70为中度富营养化,TLI(∑)>70 为重度富营养化。
1.2.3 多样性指数 同时运用 Shannon-Wiener多样性指数(H′)、Margalef丰富度指数(d)、Pielou均匀度指数(J)进行评价。计算公式分别为
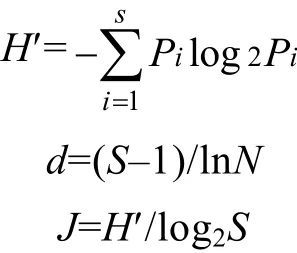
式中, S为样品中物种种类总数, n为样品中第i种浮游植物的丰度, N为样品中浮游植物总丰度。H’>3时为轻或无污染, 1—3为中污染, 其中2—3为β-中污,1—2为 α-中污, 在 0—1之间为重污染。d>5时为清洁, 在4—5之间为寡污型, 3—4之间为β-中污, 1—3为α-中污, 0—1为重污染。J值在0.8—1.0之间时为清洁, 0.5—0.8为寡污型, 0.4—0.5为β-中污, 0.3—0.4为α-中污, 0—0.3为重污染(况琪军等, 2005; 周广杰等, 2006; 郭春燕等, 2010; Spatharis et al, 2011; 刘成士等, 2015; 杨敏等, 2015; 苟婷等, 2015)。
1.3 数据处理
本文统计分析采用 SPSS 19.0与 Salford Predictive Modeler (SPM) 7.0软件实现; 基本图形绘制在Origin 9.0软件中完成。
1.3.1 分类回归树 为分析营养状态与生物多样性与浮游植物密度之间的关系, 以H′、J、d、TLI(∑)以及采样点类型(城市、农村、工业区)为预测指标, 采用分类回归树(classification and regression tree, CART)评价这5个预测指标的对浮游植物密度的重要性。对浮游植物密度进行常用对数(lg)转换, 采用R2评价CART预测效果。CART分析采用Salford Predictive Modeler (SPM) 7.0。
1.3.2 聚类分析 为探寻浮游植物的空间分布差异, 根据浮游植物种类和密度, 利用SPSS 19.0软件对不同采样断面的浮游植物进行聚类分析。浮游植物密度经过平方根转换。
2 结果与讨论
2.1 沱江环境参数与营养状态
2.1.1 主要环境参数 从季节来看, 沱江水体中的浮游植物密度、叶绿素a、TN与TP数值总体呈现出丰水期高于枯水期的趋势; 与之相反, 透明度则表现为枯水期较高(图2、图3)。这一现象可能与丰水期较大的地表径流量带来的丰富氮磷(夏品华等, 2011)及该时期适宜的水温、光照有关。其余参数无明显的季节差异。
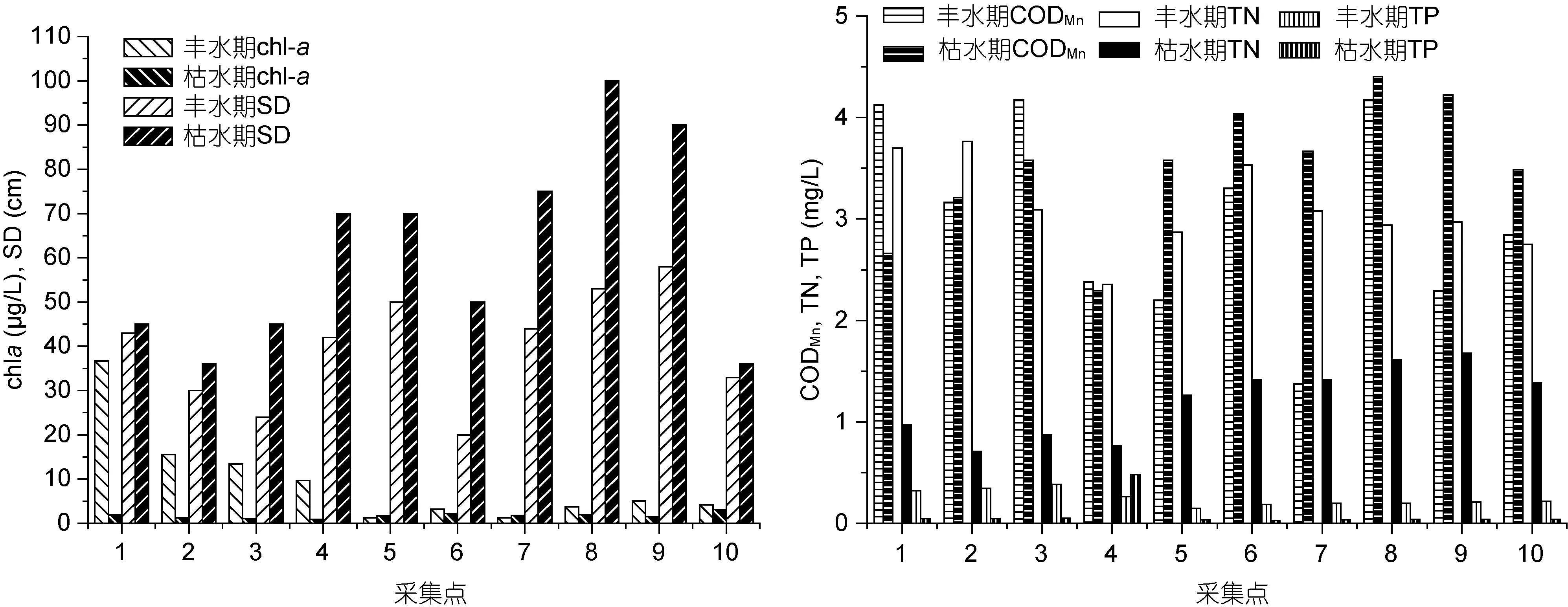
图2 叶绿素a、透明度、高锰酸盐指数、总氮和总磷的空间分布Fig.2 Spatial distribution of chlorophyll a, Secchi depth (SD), CODMn, TN and TP

图3 浮游植物种类数与密度分布Fig.3 Spatial distribution of phytoplankton species and density
从空间分布来看, 丰水期研究区域上游河段(资阳市区) 的叶绿素 a、浮游植物密度与 TP等参数值均高于中、下游河段(资中、内江市区), 见图2、图3。这可能与沱江上游磷矿开采有关(施泽明等, 2012),在丰水期高氮含量的情况下, 浮游植物的生长主要受到磷的限制(田时弥等, 2015), 故而上游水域因地表径流带来较多的磷, 浮游植物与叶绿素含量均较高。枯水季节叶绿素a、浮游植物密度与TN在中、下河段较高(图2、图3), 说明枯水期由于地表径流冲刷力度降低, 带入沱江中的氮素逐减少并成为藻类生长限制因子。在两个季节中, 透明度变化与浮游植物密度变化规律不相符(图2、图3), 可能与沱江水体中泥沙含量有关。
2.1.2 综合营养状态指数 根据综合营养状态指数 TLI(∑)的大小对研究江段水体进行富营养化状态评价(图4a), 沱江水质总体处于中-富营养状态。与四川省其它主要河流如岷江、嘉陵江等相比, 沱江的TLI(∑)较高, 富营养化较为严重(刘佳等, 2012)。枯水季节水体整体为中营养状态(<50), 水质良好; 丰水期多存在各种程度的污染, 尤其是位于资阳市区的 1—3号采样点, 富营养化程度较高(>60)。丰水期较大的降水冲刷使得更多的氮磷营养进入水体, 尤其是较接近上游磷矿的江段, 富营养化水平较高, 而中、下游水体的自净作用在一定程度上缓解了富营养化进程。
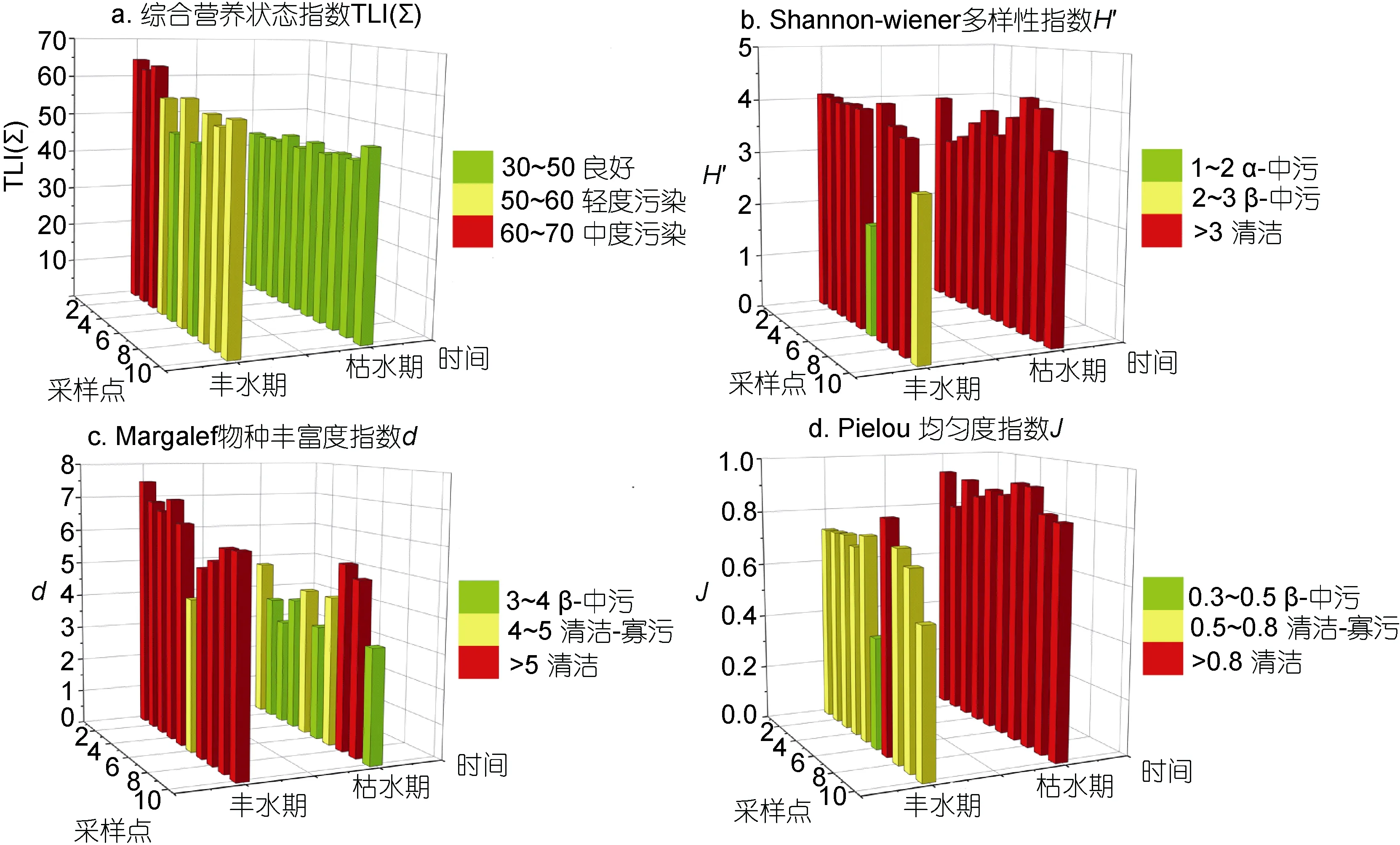
图4 各种参数的空间分布情况Fig.4 Spatial distribution of various indicators
2.2 浮游植物群落结构分析与水质评价
2.2.1 浮游植物群落组成与优势种 调查期间共检出浮游植物7门88种(属), 其中绿藻种类数最多为37种(属), 硅藻23种(属), 蓝藻15种(属), 其它门类13种(属)。浮游植物种类数在空间分布上变化不大,但在时间分布上有较大差异(图 3), 该差异主要由蓝藻和绿藻引起。丰水期各断面种类数变化范围为30—52种, 以6#最低, 4#最高; 枯水期为12—28种,3#最低, 9#最高。根据图3, 丰水期浮游植物密度主要由蓝藻(35%)、硅藻(35%)和绿藻(24%)组成; 枯水期硅藻成为优势类群(44%), 隐藻数量增多(18%), 蓝藻和绿藻水平相当(15%)。浮游植物群落结构是评价水体富营养化的重要参数。一般认为甲藻、隐藻和硅藻易在中营养型水体中占优势, 绿藻和蓝藻则易在富营养型水体中占优势(金相灿等, 1990)。沱江丰、枯水季节浮游植物群落结构分别为绿藻+硅藻+蓝藻和硅藻+绿藻+隐藻, 由此推断该江段目前应处于中到富营养状态, 尤其是丰水期, 富营养化程度更高, 这与前文营养状态分析结果一致。
研究江段浮游植物优势种较多且优势度均不高(表 1), 表明其群落结构较复杂也较为完整。硅藻在两个季节均有优势, 其中梅尼小环藻(Cyclotellameneghiniana)和花环小环藻(C. operculata)存在丰水期高峰, 脆杆藻(Fragilariasp.)、线形舟形藻(Navicula graciloides)和针杆藻(Synedrasp.)则在枯水期占优势。蓝藻和隐藻有明显的季节差异: 蓝藻为丰水期优势类群, 优势种为湖泊假鱼腥藻(Pseudanabaena limnetica)、拉氏拟柱孢藻(Cylindrospermopsis raciborskii)、微小平裂藻(Merismopedia tenuissima)和细小平裂藻(M. minima); 隐藻则主要在枯水季节占优势, 包括尖尾蓝隐藻(Chroomonas acuta)、卵形隐藻(Cryptomonas ovata)和啮蚀隐藻(C. erosa)。其它类群如绿藻和金藻优势种类少且优势度均较低。丰水期优势度最高的梅尼小环藻和微小平裂藻均为 α-中污指示种(沈蕴芬等, 1990), 且富营养化水体中常见的丝状蓝藻湖泊假鱼腥藻(黄成等, 2011)等出现频度亦较高, 说明该季节沱江已处于富营养化状态。枯水季节优势种则反映出寡污-中污状态(沈蕴芬等, 1990)。

表1 沱江浮游植物优势种及优势度Tab.1 Dominant species and dominance degree of phytoplankton in Tuojiang River
2.2.2 浮游植物群落聚类分析 聚类分析(图 5)结果显示, 研究江段中游3个位于五里店水电站坝下的断面(5#、6#、7#)聚集为集合 1; 下游天宫堂水电站坝上 3个断面(8#、9#、10#)与中上游的 4#断面共同组成集合2; 上游南津驿水电站坝上3个断面(1#、2#、3#)则聚为集合3。3个集合的蓝藻、绿藻、硅藻、隐藻比例分别为15.6%、19.4%、50%、8.7%和33.7%、20.3%、30.5%、8.7%以及21.5%、17.7%、41.4、16.9%,硅藻在坝下集合 1中所占比例最大, 蓝藻+绿藻在坝上集合2中最多, 隐藻则在坝上集合3中最多。由于水电坝的拦截作用, 大坝上下游之间生态条件会发生较大变化, 坝上湖泊区由于流速和透明度的变化其浮游植物群落结构也显著区别与坝下区域(韩博平,2010; 杨敏等, 2015), 因此与坝下集合 1相比, 坝上集合2与集合3中湖泊型蓝藻、绿藻类及隐藻增多,河流型硅藻类有所下降(邱光胜等, 2011), 这一变化趋势与嘉陵江等邻近水体接近(杨敏等, 2015)。
单因素方差分析(One-way ANOVA)显示, 浮游植物密度与NH+4数值均表现出从集合1到集合3明显升高的趋势(P<0.05, 表 2), 其余指标差异不显著(P>0.05, 表 2), 说明水电坝的建设与空间分布格局对水体多种营养盐浓度作用较小, 但对与浮游植物群落结构已有影响。由于水电坝的拦截作用, 大坝上游附近区域流速减缓, NH+4等营养积累(赵晨等, 2014), 适宜浮游植物增殖, 故坝上集合2与3中浮游植物密度与NH+4均高于坝下集合1; 同为坝上采样点, 较上游的集合3中NH+4浓度最高可能与沱江上游的企业污水排放有关(孙莉等, 2005),喜好NH+4的浮游植物(de Aldaet al, 1996)密度也随之更高。

图5 采样点聚类分析树状图Fig.5 Cluster analysis for the sampling sites

表2 沱江不同采样断面集群水体特征比较Tab.2 Comparison of biological and physiochemical parameters among sample sites in 3 groups
2.2.3 多样性指数 沱江浮游植物多样性指数变化趋势如图 4b—d。丰水期中各指数变化较为一致,物种多样性指数H′平均3.66, 大致处于清洁状态, 最高值出现在 7#断面(4.14), 最低为 6#断面(1.98); 物种丰富度指数d平均6.21, 整体为清洁, 1#断面最高(7.47), 6#断面最低(4.39); 均匀度指数J平均0.69, 多为清洁-寡污状态, 与多样性指数H′一样, 最高与最低值分别出现在6#(0.84)和7#(0.40)。6#与10#等断面物种多样性与均匀度指数较低呈中度污染状态, 是由于细胞很小的群体蓝藻, 如平裂藻属(Merismopediaspp.)的优势造成。尽管该类藻细胞数量较大, 但对浮游植物生物量和水质的影响却较小, 形成水华的可能性也较低, 根据此结果丰水期沱江处于清洁-寡污状态。枯水期各指数一致性较差: 多样性指数H′(3.13—4.21)与均匀度指数J(0.80—0.93)各点差异不明显; 物种丰富度指数d最低值出现在 3#(3.12), 最高值为 8#断面(5.40), 多数点处于轻度到中度污染状态。三个指数中, 物种丰富度指数d(图4)与浮游植物种类数(图 3)变化趋势一致性较好, 说明该指数对沱江中浮游植物群落物种数变化较为敏感, 因此按物种丰富度指数d评价标准, 枯水季节沱江各断面多处于轻度到中度污染状态。但是应当看到, 尽管从多样性来说沱江污染程度较低, 其实营养水平和浮游植物群落结构却呈现出明显的富营养化状态。由于环境的复杂性, 从不同角度评价水体状态均有其局限性(易斌等, 2007), 综合各种指标进行判定, 沱江污染程度仍不容乐观。
2.3 浮游植物密度与营养和物种多样性的关系
浮游植物群落结构受外界环境(营养水平)与内部因素(种内与种间关系, 即群落结构)的共同作用,从数据上来说, 浮游植物密度与营养状态指数以及多样性指数有某种关联。以浮游植物密度为因变量,综合营养状态指数和多样性指数为自变量所作的分类回归树模型R2=0.93, 说明这两类指数能较好地解释浮游植物密度的变化。该模型将浮游植物密度数据作了3次细分(图6)。第一个分支将4个物种丰富度较高(d>6.56)并且浮游植物密度较高(未进行 lg转换的平均值为 4.6×106cell/L)的样品与其它样品分离开来; 第二个分支又将 5个具有较低营养水平(TLI≤43.9)且浮游植物密度较低(平均值为0.4 ×106cell/L)的样品从数据集中分离出来; 第三个分支将剩余的 11个样品以J=0.85为标准分为两部分, 均匀度低(≤0.85)的 8个样品浮游植物密度相对较高(平均值为2.6×106cell/L), 均匀度较高(>0.85)的 3 个样品浮游植物密度较低(平均 1.0 ×106cell/L)。Shannon-Wiener多样性指数(H′)与采样点位置类型未被纳入模型中。该模型显示, 营养水平与物种丰富度较高, 但均匀度偏低的水体中, 浮游植物密度易出现高值。在营养物质富足的情况下, 浮游植物种类较多, 但有一部分为偶见种(俞秋佳, 2014), 少数优势种的大量繁殖降低了群落均匀度, 这些优势种若继续发展, 水体就有发生水华的潜在风险(冯波等, 2008)。对汉江硅藻水华的研究表明, 较丰富的物种多样性有可能是水华初期的特点(潘晓洁等, 2014)。因此在营养水平较高的水体中,为预警水华的发生, 丰富的浮游植物种类不仅不能成为放松警惕的原因, 更应成为密切监测的理由。

图6 浮游植物密度与综合营养状态指数、多样性指数间的关系Fig.6 Classification and regression tree of phytoplankton abundance versus trophic level index and diversity index x为未进行lg转换的浮游植物密度平均值; n为样本数量
3 结论
根据综合营养状态指数判断沱江水体处于中-富营养状态, 富营养化程度较高。由于雨水淋溶冲刷作用和上游磷矿的影响, 丰水期污染程度高于枯水期,上游断面高于较下游断面。
从浮游植物群落组成来看, 沱江为中-富营养型水体, 其中丰水期为绿藻+硅藻+蓝藻的形式, 枯水期为硅藻+绿藻+隐藻的形式, 仍表现出丰水期富营养化程度高于枯水期的状态。优势种类的组成显示, 沱江中浮游植物群落结构较为复杂和完整, 丰水期为α-中污染, 枯水期则为寡污-中污状态。由于沿江梯级大坝的影响, 沱江水文条件与 NH4+分布具有明显空间异质性, 导致了大坝上下游之间、上游断面与下游断面之间浮游植物群落结构与密度的差异, 整体表现出大坝上游湖泊型藻类数量增加、上游断面浮游植物密度高于下游断面的趋势。尽管多样性指数显示沱江污染状态较轻, 综合考虑各类指标的局限性, 沱江污染程度仍不容乐观。
沱江中浮游植物密度受到营养和群落结构的显著影响: 在营养水平高, 物种丰富度较大但均匀度较低的情况下, 容易暴发水华。因此对于营养物质含量丰富并且浮游植物种类数较多的水体, 不能放松警惕, 应加强监控。
王爱爱, 冯 佳, 谢树莲, 2014. 汾河中下游浮游藻类群落特征及水质分析. 环境科学, 5(3): 915—923
中国环境监测总站, 2001. 湖泊(水库)富营养化评价方法及分级技术规定. 北京: 中国环境科学出版社
田时弥, 杨 扬, 乔永民等, 2015. 珠江流域东江干流浮游植物叶绿素 a时空分布及与环境因子的关系. 湖泊科学,27(1): 31—37
冯 波, 陈俊辉, 卢伙胜, 2008. 茂名市大放鸡岛海域浮游植物群落的季节变化. 海洋渔业, 30(1): 67—73
刘 佳, 徐 亮, 张秋劲等, 2012. 四川省水体富营养状况时空分布调查. 中国环境监测, 28(5): 6—8
刘成士, 黄 燕, 刘 瑜等, 2015. 青山水库轮虫群落结构特征的时空差异对水质变化的影响. 海洋与湖沼, 46(3):694—702
刘建平, 2013. 沱江富顺段水质分析及污染状况研究. 四川环境, 32(2): 23—26
孙 莉, 朱鸿斌, 张成云等, 2005. 一起工业废水污染沱江水源水事故的调查. 环境与健康杂志, 22(3): 192—193
李 进, 1986. 沱江水质污染的微生物监测评价. 四川环境,5(3): 26—32
杨 敏, 张 晟, 刘朔孺, 2015. 草街水库蓄水后嘉陵江浮游植物群落特征及水质评价. 环境科学, 36(7): 2480—2486
杨昌述, 1987. 沱江干流底栖动物调查及水质生物学评价. 环境科学, 8(5): 50—56
肖玖金, 谭周亮, 李旭东等, 2011. 基于物元分析法的沱江地表水水质评价. 安全与环境学报, 11(1): 104—108
邱光胜, 胡 圣, 叶 丹等, 2011. 三峡库区支流富营养化及水华现状研究. 长江流域资源与环境, 20(3): 311—316
况琪军, 马沛明, 胡征宇等, 2005. 湖泊富营养化的藻类生物学评价与治理研究进展. 安全与环境学报, 5(2): 87—91
沈韫芬, 章宗涉, 龚循矩等, 1990. 微型生物监测新技术. 北京: 中国建筑工业出版社, 1—524
陈雨艳, 余 恒, 向秋实等, 2015. 沱江流域水环境质量分析.四川环境, 34(2): 85—89
苟 婷, 马千里, 许振成等, 2015. 贝江浮游藻类群落特征及富营养化风险分析. 环境科学, 36(3): 946—954
国家环境保护总局, 2002. 水和废水监测分析方法(第四版).北京: 中国环境科学出版社
易 斌, 吴玲玲, 林 端等, 2007. 营养指数法和浮游植物多样性指数法在评价富营养化上的差异. 见: 中国海洋学会赤潮研究与防治专业委员会第二届学术研讨会. 上海: 中国海洋学会
金相灿, 屠清瑛, 1990. 湖泊富营养化调查规范(第二版). 北京: 中国环境科学出版社
周广杰, 况琪军, 胡征宇等, 2006. 三峡库区四条支流藻类多样性评价及“水华”防治. 中国环境科学, 26(3): 337—341
周开锡, 李 敏, 吴 建, 2015. 沱江河内江段主要监控断面水质变化趋势分析. 见: 2015年中国环境科学学会学术年会论文集(第一卷). 深圳: 中国环境科学学会
周实际, 朱 俊, 张 科等, 2011. 沱江内江城区段水质分析.环境科学与管理, 36(10): 114—116
赵 晨, 董世魁, 刘世梁等, 2014. 漫湾大坝上下游沉积物重金属与营养元素分布特征及环境风险评价. 环境科学学报, 34(9): 2417—2425
胡鸿钧, 魏印心, 2006. 中国淡水藻类—系统、分类及生态. 北京: 科学出版社, 23—915
俞秋佳, 2014. 苏州河浮游植物群落变化特征及其影响因子研究. 上海: 华东师范大学硕士学位论文
施泽明, 倪师军, 张成江等, 2012. 沱江流域磷矿开采和加工过程中放射性环境问题探讨. 地球科学进展, 27(10):1134—1139
夏品华, 李秋华, 林 陶等, 2011. 贵州高原百花湖水库湖沼学变量特征及环境效应. 环境科学学报, 31(8):1660—1669
郭春燕, 冯 佳, 谢树莲, 2010. 山西晋阳湖浮游藻类分布的时空格局及水质分析. 湖泊科学, 22(2): 251—255
陶 敏, 王永明, 谢碧文等, 2016. 沱江浮游生物群落时空分布及相关环境因子分析. 水生生物学报, 40(2): 301—312
黄 成, 侯 伟, 顾继光等, 2011. 珠江三角洲城市周边典型中小型水库富营养化与蓝藻种群动态. 应用与环境生物学报, 17(3): 295—302
章宗涉, 黄祥飞, 1991. 淡水浮游生物研究方法. 北京: 科学出版社
韩博平, 2010. 中国水库生态学研究的回顾与展望. 湖泊科学,22(2): 151—160
焦珂伟, 周启星, 2015. 基于水质与生物指标的松花江流域水生态健康评价. 生态学杂志, 34(6): 1731—1737
谢贤健, 兰代萍, 2009. 基于因子分析法的沱江流域地表水水质的综合评价. 安徽农业科学, 37(3): 1304—1306
潘晓洁, 朱爱民, 郑志伟等, 2014. 汉江中下游春季浮游植物群落结构特征及其影响因素. 生态学杂志, 33(1): 33—40
Bozzetti M, Schulz U H, 2004. An index of biotic integrity based on fish assemblages for subtropical streams in southern Brazil. Hydrobiologia, 529(1): 133—144
de Alda J A G O, Tapia M I, Franck F et al, 1996. Changes in nitrogen source modify distribution of excitation energy in the cyanobacterium Phormidium laminosum. Physiologia Plantarum, 97(1): 69—78
Spatharis S, Roelke D L, Dimitrakopoulos P G et al, 2011.Analyzing the (mis) behavior of Shannon index in eutrophication studies using field and simulated phytoplankton assemblages. Ecological Indicators, 11(2): 697—703
猜你喜欢
中国新闻周刊(2024年19期)2024-06-21 02:38:27
食品安全导刊·中旬刊(2023年4期)2023-05-31 21:35:50
特区文学·诗(2022年1期)2022-02-26 03:43:38
潍坊学院学报(2020年2期)2021-01-18 07:02:00
中国卫生产业(2020年25期)2020-11-30 08:50:15
流行色(2019年10期)2019-12-06 08:13:26
——多棘沱江龙
娃娃乐园·综合智能(2018年23期)2018-12-26 08:54:12
华人时刊(2018年17期)2018-12-07 01:01:56
黄河之声(2018年17期)2018-11-20 10:43:40
环境科技(2016年2期)2016-11-08 12:18:22
