
2012年长江口及其邻近海域营养盐分布的季节变化及影响因素*
2016-01-15 08:21王文涛曹西华袁涌铨宋秀贤俞志明
海洋与湖沼 2016年4期
王文涛 曹西华 袁涌铨 宋秀贤 俞志明①
(1. 中国科学院海洋研究所 海洋生态与环境科学重点实验室 青岛 266071; 2. 青岛海洋科学与技术国家实验室 海洋生态与环境科学功能实验室 青岛 266071; 3. 中国科学院大学 北京 100049)
海水中的营养盐控制着海洋初级生产力, 也是浮游植物组成的重要元素。有研究表明, 含有氮、磷、硅的营养盐的浓度与比例直接影响浮游植物的种群动态和群落结构(Hori et al, 1998; Hodgkiss et al, 2004;Mallin et al, 2004)。近年来, 受到人类活动的影响, 许多近岸海域出现海水富营养化现象, 导致有害藻华的暴发(HABs, Harmful Algal Blooms), 造成赤潮等生态灾害, 严重危害了海洋生态系统的健康(Glibert et al, 2001; Diaz et al, 2008)。作为世界第三, 同时也是太平洋西岸最大的河流, 长江每年向邻近的中国东海输入大量营养盐, 使得长江口海域富营养化加剧及赤潮等生态灾害频繁暴发, 对人类在这一区域的渔业生产和生态系统造成了严重危害(俞志明等,2001; 周名江等, 2003)。
针对这一现象, 许多学者对长江口海域的营养盐进行了大量研究。参考近些年的营养盐变化数据可以发现, 自 1985年以来由于受到长江径流量、输沙量以及人类使用化肥结构的改变, 长江口水域的 N/P比值呈M型下降趋势(陈慧敏等, 2011)。Chai等(2009)则结合了三峡大坝的建设情况, 通过分析 2002年至2006年期间长江口及邻近海域的营养盐浓度变化,发现自大坝蓄水以来长江口水域的 Si/N比值降低,而由于泥沙量输入减少导致这一海域的总磷含量也随之下降。也有学者发现长江口海域冬季铵盐和亚硝酸盐的增加可能与寒冷水团中的硝化反应降低有关,而硝酸盐与磷酸盐的浓度在最近十年中逐渐降低(Gao et al, 2012)。
虽然许多学者对长江口海域的营养盐进行了大量研究, 但是由于该区域人类活动密集, 并存在三峡等大型水利设施可能影响长江的输运, 导致河口海域的营养盐时空变化和年际差异等特征较为复杂, 因此需要开展持续性地调查和研究工作, 记录并分析这一关键区域营养盐的常年变化特征及影响因素。基于这一背景, 本研究调查了2012年四个季节长江口及邻近海域氮、磷、硅营养盐和总氮总磷的浓度, 结合历史资料, 分析营养盐的浓度变化、结构特点和潜在营养盐限制, 并讨论其不同成分的来源及年度变化特征。通过如上分析, 揭示营养盐含量变化对生态系统的结构功能和稳定性的影响。
1 材料与方法
1.1 采样站位
本文涉及调查区域为长江口及其邻近海域, 范围为 121.0°—123.5°E、30.5°—32.0°N, 调查站位 44个(图1), 分别于2012年3月4日—20日(冬季), 5月22日—28日(春季), 8月30日—9月6日(夏季), 12月10日—14日(秋季)共四个航次进行采样。根据盐度与浊度特征将调查海域分为三个区域, 区域一: 口门内,盐度≤10; 区域二: 最大浑浊带(文中统称浑浊带),10<盐度≤25; 区域三: 外海区, 盐度>25。
1.2 调查方法
根据各站位的水深, 采用颠倒式采水器分别采集表层、5m、10m、20m、30m和底层水样, 调查内容包括温度、盐度、叶绿素a(chl a)、硝酸盐(NO3)、亚硝酸盐(NO2)、氨氮(NH4)、磷酸盐(PO4)、硅酸盐(SiO3)、总氮(TN)及总磷(TP)等。采集的部分水样经高温灼烧处理过的Whatman GF/F滤膜过滤后装入洁净的聚乙烯瓶, 滴入氯仿固定并保存在-20°C下, 待测溶解态营养盐; 另有部分水样直接装入洁净的聚乙烯瓶中, 滴入浓硫酸固定并置于-20°C保存, 用于测定海水中的TN、TP。
1.3 分析方法
使用颠倒水温表测定海水温度, 仪器误差为±0.01°C, 使用 8803型盐度计测定海水盐度, 仪器误差为±0.01。Chl a 通过丙酮萃取后用荧光法测定, 检测限为0.01mg/m3(Parsons et al, 1984)。营养盐的测定参照《海洋调查规范》进行, 使用镉铜还原法测定硝酸盐, 重氮-偶氮法测定亚硝酸盐, 靛酚蓝法测定氨氮, 磷钼蓝法测定磷酸盐, 硅钼蓝法测定硅酸盐。用过硫酸钾氧化总氮总磷后, 分别使用镉铜还原法和磷钼蓝法测定。使用荷兰制造的 SKALAR连续流动分析仪测定各项营养盐浓度, 标准曲线满足相关系数≥0.999, 并借助标准溶液进行中间校准以控制数据质量。总溶解无机氮(DIN)为 NO3、NO2、NH4之和。
文中采样站位图使用 Surfer 12制作, 营养盐比值图用 Sigma Plot 12绘制, 相关性及潜在限制图用Excel 2010绘制。由于缺少秋季TN、TP与盐度的对应站位, 因此在该季节仅分析DIN、PO4和SiO3与盐度的相关性; 营养盐浓度均值差异及相关性的显著分析利用 SPSS Statistics 19中的单因素方差法(ANOVA)。
2 结果与讨论
2.1 营养盐时空分布
将调查海域划分为口门内、浑浊带和外海区, 分别代表长江河口淡水端、咸淡水混合区域和高盐外海水影响海域。在四个季节各区域内全水层营养盐的浓度范围及平均值如表1所示。
DIN、SiO3、TN和 PO4的浓度均表现出自口门内向外海区逐渐降低的分布特征, 前三者的降低幅度较为明显, 降低倍数约在 4—10倍之间; 随着远离河口区域, PO4平均浓度的降低幅度相对减弱, 最大浓度差不超过3倍, 且季节性差异较小。与其它营养盐不同, TP在浑浊带区域表现出最高平均浓度, 且在冬季呈现最高值。DIN和TN在长江口全海域的变化规律相似, 其平均浓度自冬季开始降低, 在夏季达到最低值后又逐渐升高(表1)。

表1 长江口及邻近海域全水层DIN、PO4、SiO3、TN、TP浓度(μmol/L)范围及平均值的季节变化特征Tab.1 Seasonal changes of DIN, PO4, SiO3, TN and TP concentrations in the whole layers of Changjiang River estuary and adjacent area
2.2 营养盐变化特点及影响因素
2.2.1 磷营养盐 与DIN、SiO3和TN相比, 自口门内向外海区 PO4浓度的降低幅度较小(表 1), 且与盐度的相关性相对较低(图 2), 可能是由于除长江冲淡水外, PO4还存在其它重要来源。作为重要的非保守性营养要素, PO4浓度在河口海域受到多种因素影响, 一方面受到浮游植物的吸收消耗所转移, 另一方面受到泥沙吸附和释放的影响(徐明德等, 2006), 咸淡水混合区域磷酸盐的含量相对稳定(黄自强等,1994)。另外, 黑潮水入侵东海(包括长江口及其邻近海域)被认为是 PO4的重要来源(Chen, 1999; Fang,2004), 本次调查也发现在 27和 34站底层出现盐度>34的高值区, 可能是外海高盐水入侵所致。Wang等人(2016)的研究也证实, 富含磷酸盐的黑潮东海分支可以入侵至外海区31°N的123°—123.5°E附近, 从而向该海域补充高浓度PO4。
与其它营养物质不同, 四个季节中 TP的最大平均值均出现在长江口浑浊带海域(表 1)。陈野等人(2014)的研究表明, 靠近河口的浑浊带海域蕴含的泥沙可以吸附大量颗粒态磷(PP), 而PP是TP的主要组成部分, 其浓度可以达到溶解态磷酸盐的 2—5倍(Froelich, 1988; 徐开钦等, 2004)。本文调查结果也显示, 各个季节长江口浑浊带的 TP平均浓度均为 PO4浓度的5倍以上。此外, Liu 等(2003)发现长江干流悬浮体浓度与 TP浓度呈现正相关特征, 因此推测 TP在浑浊带出现高值是由于泥沙吸附所致。
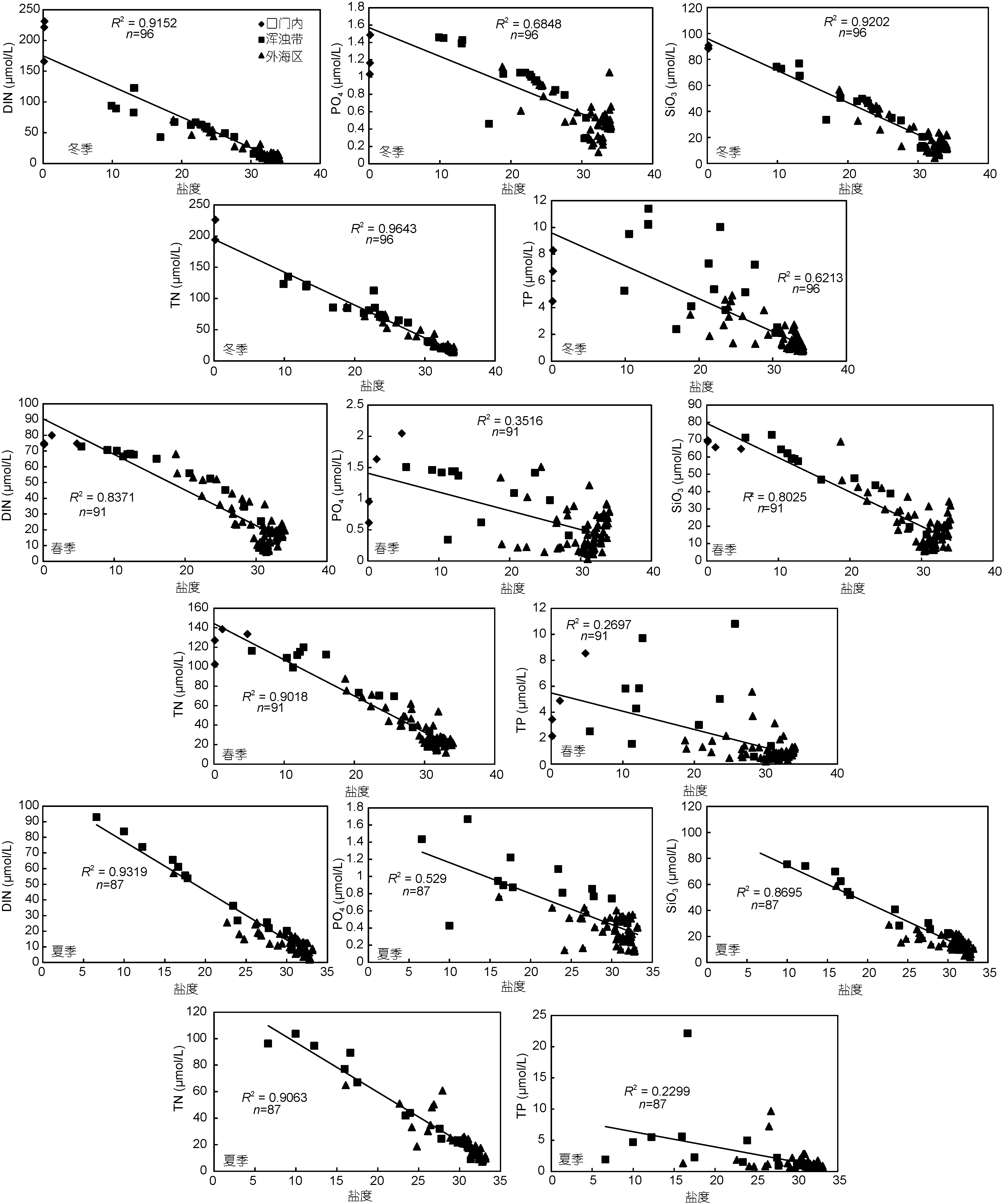
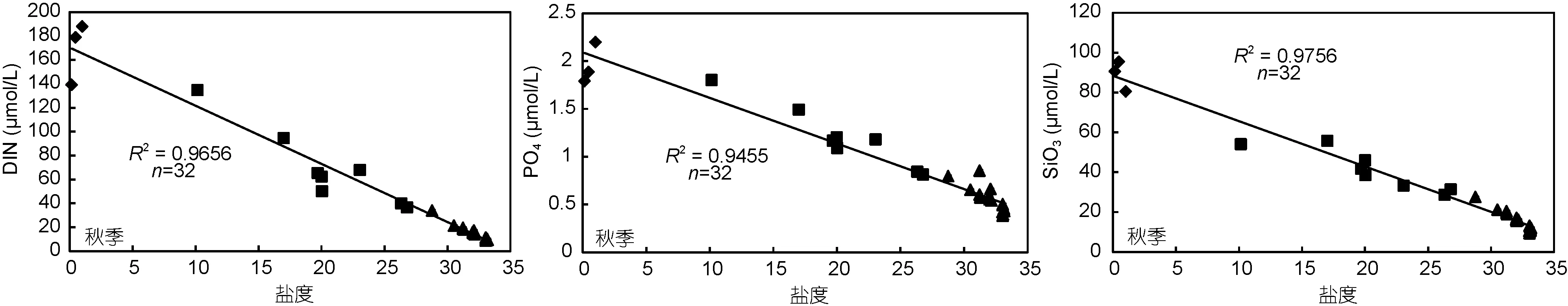
图2 长江口及邻近海域营养盐与盐度的相关性Fig.2 Correlations between nutrients and salinity in the Changjiang River estuary and adjacent area
2.2.2 氮营养盐 DIN及TN在夏季表现出较低的平均值, 其中DIN的分布特点与王江涛等(2012)总结的近50年来长江口DIN的季节变化特征相似, 即低浓度出现在春夏季节, 这一特征可能是由于浮游植物生长吸收所致。Chl a的季节变化显示, 在本研究四个季节的调查中chl a的平均浓度在夏季达到最高值, 而在秋季(12月)处于最低水平(图 3), 与 DIN和TN的变化特点相反。杨红等(2008)利用BOX模型研究发现, 在长江口及其邻近海域夏季河流输入 DIN通量小于交换流带出系统的 DIN通量。近些年甲藻赤潮逐渐增多, 对含氮营养盐的影响也更为显著(王江涛等, 2012), 因此推测夏季DIN和TN的低浓度是受浮游植物生长吸收所致。
此外, DIN和TN的平均浓度表现出口门内>浑浊带>外海区的规律, 且与盐度呈显著负相关(P<0.01,图2), 推测DIN和TN在长江口及邻近海域的主要来源是长江冲淡水。此特征与众多研究结果一致, 沈志良(1997)报道了在长江入海口区域 DIN的通量处于较高水平, 而 Wang等(2003)的研究也发现长江河口以东及东北区域的高营养盐来自长江冲淡水的贡献。

图3 长江口及邻近海域叶绿素a的平均浓度分布Fig.3 Distributions of average chl a concentrations in the Changjiang River estuary and adjacent area
2.2.3 年际变化分析 为了便于与以往的研究结果进行对比, 将本次调查区域的浑浊带与外海区统称为口门外区域, 口门内仍为长江口淡水端。本研究中主要营养盐浓度与以往调查结果的比较如表2所示, 本次调查的营养盐浓度范围取相应海域各个季节的平均值。
对比结果显示, 自上世纪八十年代以来 DIN和PO4在口门内的浓度呈现显著升高的特征, 其中第一个阶段的升高更为明显, 长江流域及沿海城镇化发展带来大量的工农业污水排放是该海域氮磷营养盐增加的主要原因(王奎等, 2011); 近十年氮、磷营养盐浓度也有一定程度的提升, 但变化幅度不大。在口门外海域, DIN与PO4也呈现出逐渐升高的特点, 第一阶段的升高幅度明显小于口门内的淡水端。然而, 口海域 SiO3的浓度则呈现相反特征, 第一阶段 SiO3浓度呈现下降特征, 近十年来变化幅度较小, Li等(2007)认为长江流域水利工程的修建拦截了大量沉积物质,导致入海的SiO3浓度明显降低。
2.3 营养盐结构特征及影响因素
2.3.1 营养盐结构的时空变化 海水中氮磷等营养物质的含量及结构组成, 不仅影响着海域的初级生产力, 对浮游植物群落组成也起着至关重要的作用。调查发现N/P比值表现出远离河口而逐渐减小的特点。在2.2中讨论了调查海域内DIN主要来自于长江冲淡水, 而磷酸盐主要来自于颗粒物分解以及外海输入, 由于从口门内至外海区长江冲淡水的影响逐渐减弱而外海水影响增强, 因此N/P比值表现出远离河口而逐渐减小的特点(图 4)。浑浊带较低的透光率限制了浮游植物的生长, 由此也抑制了浮游植物对营养盐的吸收利用, 因此该区域N/P比值无明显周年变化特征(诸大宇等, 2008)。
在口门内、浑浊带及外海区, Si/N平均值的最高值均出现在夏季(图4), 差异性非常显著(P<0.01); 而在调查全海域中, 可以发现秋季的 Si/P比值相对较低(图 4), 该季节减弱的长江水输送可能是导致此特征的重要原因。

表2 长江口及邻近海域营养盐浓度(μmol/L)变化Tab.2 Variations of nutrients concentrations (μmol/L) in the Changjiang River estuary and adjacent area

图4 长江口及邻近海域营养盐结构变化特征Fig.4 Variations of nutrients structures in the Changjiang River estuary and adjacent area
2.3.2 浮游植物生长的营养盐限制 N、P、Si等元素作为浮游植物生长繁殖的重要营养成分, 其在海水中的含量组成可以直接影响海洋初级生产力与生态系统结构和功能(Hodgkisset al,1997; Flynn,2002)。早在1963年Redfield等就提出浮游植物吸收海水中营养盐按照 N∶P∶Si = 16∶1∶16的浓度比例进行(Redfieldet al, 1963); 此后, Justić等(1995)定义一般情况下海水中的营养盐 SiO3= 2μmol/L, DIN=1μmol/L, PO4= 0.1μmol/L 可作为浮游植物生长的最低阈值。依据此限制条件, 发现调查期间仅在春季33号站位存在绝对P限制(PO4= 0.03μmol/L)。
由于长江口海域营养盐常年处于较高浓度的状态, 因此对浮游植物生长形成绝对限制的情况很少。但营养盐比例失衡将影响并改变海域浮游植物的群落结构和组成, 所以我们进一步考察了调查海域浮游植物生长潜在营养盐限制的状况。通常认为, 当N∶P>22、Si∶P>22时, 处于潜在磷限制状态; 当 N∶P<10、Si∶N>1 时, 处于潜在氮限制状态; 当 Si∶N<1、Si∶P<10 时, 处于潜在硅限制状态(Justićet al,1995)。依据该标准, 发现冬季有超过60%的站位处于潜在磷限制状态(图 5), 春季、夏季和秋季这一比例分别为90%、92%和41%(表3), 这一特征与chla的时空变化相似, 说明浮游植物的吸收是引起潜在营养盐限制的重要原因之一。而出现P限制特征的站位均位于外海区, 推测输送至外海区的磷酸盐被浮游植物进一步吸收利用, 引起这一海域大范围的潜在P限制特点。与潜在P限制不同, 调查期间的所有海域基本未发现潜在 N限制特征; 仅在秋季发现四个位于长江口外海区的站位有潜在Si限制的可能性。

图5 长江口及其邻近海域潜在营养盐限制特征Fig.5 Potential limitations of nutrients in the Changjiang River estuary and adjacent area a: P 限制; b: N 限制; c: Si限制
2.3.3 潜在限制特征的年际变化 与文献报道(王奎等, 2013)的 2006—2007年期间调查结果比较, 发现本次调查中春季和夏季潜在 P限制的站位比例呈现增加的特点, 而潜在 N限制和潜在 Si限制的站位则明显减少(表 3)。这一特征可能是由于营养盐输入增加和浮游植物种类演替所致。一方面, 根据之前的对比结果, 发现近 10年来长江输送的氮营养物质均有一定程度增加, 导致调查区域存在较高浓度的 N营养盐(表2), 因此基本无潜在N限制。另一方面, 研究还发现(王江涛等, 2012), 随着长江冲淡水输送营养盐的增多, 赤潮暴发的频率也大大增加, 而且优势种由硅藻逐渐向甲藻转变。另据报道, 2006年在长江河口赤潮发生海区优势种为中肋骨条藻和东海原甲藻赤潮(国家海洋局, 2011), 而在2012年赤潮优势种则为东海原甲藻和米氏凯伦藻(国家海洋局, 2013)。优势种的转变导致对 Si营养盐的吸收降低, 可能是这一区域潜在Si限制减少的原因之一。
3 结论
(1) 在调查期间, DIN、SiO3和TN等营养盐主要受长江冲淡水的影响, 平均浓度表现出从长江口门内至外海区逐渐减小的特征, 且均与盐度呈现显著负相关特征。PO4除受冲淡水影响外, 可能还受外海水输送和泥沙吸附的作用, 因此与盐度的相关性相对较低。由于浑浊带富含的泥沙吸附了较高含量的PP, 使得 TP在这一区域表现出最大浓度。受浮游植物生长吸收作用的影响, DIN和TN在长江径流量最高的夏季全海域出现最低平均浓度。

表3 长江口及邻近海域营养盐限制特征Tab.3 Characteristics of nutrients limitations in the Changjiang River estuary and adjacent area
(2) N∶P比值呈远离河口而逐渐减小的特征, 其中浑浊带区域因浮游植物光合作用低, 因此各季节间无明显氮磷比变化。受到长江径流高营养盐输入的影响, 除春季外海区有一个站位出现绝对 P限制外,调查海域均未表现出绝对营养盐限制。然而调查海域中多数站位存在潜在P限制特征, 春季和夏季的比例超过90%; 潜在N限制站位比例减小, 可能是受到长江输送氮营养盐增多所致; 而潜在 Si限制站位比例降低, 推测是由于调查海域浮游植物种类演替所致。
王 奎, 陈建芳, 金海燕等, 2011. 长江口及邻近海域营养盐四季分布特征. 海洋学研究, 29(3): 18—35
王 奎, 陈建芳, 金海燕等, 2013. 长江口及邻近海区营养盐结构与限制. 海洋学报, 35(3): 128—136
王江涛, 曹婧, 2012. 长江口海域近 50a来营养盐的变化及其对浮游植物群落演替的影响. 海洋环境科学, 31(3):310—315
杨 红, 王春峰, 2008. 长江口海域营养盐通量的估算. 上海海洋大学学报, 17(6): 700—706
沈志良, 1991. 三峡工程对长江口海区营养盐分布变化影响的研究. 海洋与湖沼, 22(6): 540—546
沈志良, 1997. 长江干流营养盐通量的初步研究. 海洋与湖沼,28(5): 522—528
陈 野, 李青云, 曹慧群, 2014. 河流泥沙吸附磷的研究现状与展望. 长江科学院院报, 31(5): 12—16
陈慧敏, 孙承兴, 仵彦卿, 2011. 近 23a来长江口及其邻近海域营养盐结构的变化趋势和影响因素分析. 海洋环境科学, 30(4): 551—553, 563
国家海洋局, 2011. 2006年中国海洋灾害公报. 中国海洋信息网, http://www.coi.gov.cn/gongbao/zaihai/201107/t20110729_17723.html
国家海洋局, 2013. 2012年中国海洋灾害公报. 中国海洋信息网, http://www.coi.gov.cn/gongbao/zaihai/201303/t20130311_26237.html
周名江, 颜 天, 邹景忠, 2003. 长江口邻近海域赤潮发生区基本特征初探. 应用生态学报, 14(7): 1031—1038
俞志明, 沈志良, 2011. 长江口水域富营养化. 北京: 科学出版社, 549
徐开钦, 林诚二, 牧秀明等, 2004. 长江干流主要营养盐含量的变化特征-1998~1999年日中合作调查结果分析. 地理学报, 59(1): 118—124
徐明德, 韦鹤平, 李 敏等, 2006. 长江口泥沙与沉积物对磷酸盐的吸附和解吸研究. 太原理工大学学报, 37(1):48—50, 54
诸大宇, 郑丙辉, 雷 坤等, 2008. 基于营养盐分布特征的长江口附近海域分区研究. 环境科学学报, 28(6):1233—1240
黄自强, 暨卫东, 1994. 长江口水中总磷、有机磷、磷酸盐的变化特征及相互关系. 海洋学报, 16(1): 51—60
Chai C, Yu Z M, Shen Z L et al, 2009. Nutrient characteristics in the Yangtze River Estuary and the adjacent East China Sea before and after impoundment of the Three Gorges Dam.Science of the Total Environment, 407(16): 4687—4695
Chen C T A, Wang S L, 1999. Carbon, alkalinity and nutrient budgets on the East China Sea continental shelf. Journal of Geophysical Research: Oceans, 104(C9): 20675—20686.
Diaz R J, Rosenberg R, 2008. Spreading dead zones and consequences for marine ecosystems. Science, 321(5891):926—929
Fang T H, 2004. Phosphorus speciation and budget of the East China Sea. Continental Shelf Research, 24(12): 1285—1299
Flynn K J, 2002. How critical is the critical N: P ratio. Journal of Phycology, 38(5): 961—970
Froelich P N, 1988. Kinetic control of dissolved phosphate in natural rivers and estuaries: a primer on the phosphate buffer mechanism. Limnology and Oceanography, 33(4 Part 2):649—668
Gao L, Li D J, Zhang Y W, 2012. Nutrients and particulate organic matter discharged by the Changjiang (Yangtze River): seasonal variations and temporal trends. Journal of Geophysical Research: Biogeosciences (2005—2012),117(G4), doi: 10.1029/2012JG001952
Glibert P M, Magnien R, Lomas M W et al, 2001. Harmful algal blooms in the Chesapeake and coastal bays of Maryland,USA: Comparison of 1997, 1998, and 1999 events. Estuaries,24(6): 875—883
Hodgkiss I J, Ho K C, 1997. Are changes in N: P ratios in coastal waters the key to increased red tide blooms. Hydrobiologia,352(1—3): 141—147
Hodgkiss I J, Lu S H, 2004. The effects of nutrients and their ratios on phytoplankton abundance in Junk Bay, Hong Kong.In: Ang Jr P O ed. Asian Pacific Phycology in the 21st Century: Prospects and Challenges. Netherlands: Springer,215—229
Hori Y, Miyahara K, Nagai S et al, 1998. Relationships between the dominant phytoplankton and DIN: DIP ratios in Osaka Bay and Harima-Nada. Nippon Suisan Gakkaishi, 64(2):243—248
Justić D, Rabalais N N, Turner R E, 1995. Stoichiometric nutrient balance and origin of coastal eutrophication. Marine Pollution Bulletin, 30(1): 41—46
Li M T, Xu K Q, Watanabe M et al, 2007. Long-term variations in dissolved silicate, nitrogen, and phosphorus flux from the Yangtze River into the East China Sea and impacts on estuarine ecosystem. Estuarine, Coastal and Shelf Science,71(1—2): 3—12
Liu S M, Zhang J, Chen H T et al, 2003. Nutrients in the Changjiang and its tributaries. Biogeochemistry, 62(1):1—18
Mallin M A, Parsons D C, Johnson V L et al, 2004. Nutrient limitation and algal blooms in urbanizing tidal creeks.Journal of Experimental Marine Biology and Ecology,298(2): 211—231
Parsons T R, Maita Y, Lalli C M, 1984. A Manual of Chemical and Biological Methods for Seawater Analysis. Oxford,USA: Pergamon Press
Redfield A C, Ketchum B H, Richards F A, 1963. The influence of organisms on the composition of seawater. In: Hill M N ed. The Sea, Vol. 2. New York: Wiley Interscience, 26—79
Wang B D, Wang X L, Zhan R, 2003. Nutrient conditions in the Yellow Sea and the East China Sea. Estuarine, Coastal and Shelf Science, 58(1): 127—136
Wang W T, Yu Z M, Song X X et al, 2016. Characteristics of thedistribution and its drivers in the Changjiang River estuary and adjacent waters. Chinese Journal of Oceanology and Limnology, doi: 10.1007/s00343-016-5276-x
猜你喜欢
海洋信息技术与应用(2021年3期)2022-01-17
海洋通报(2020年2期)2020-09-04
建筑热能通风空调(2018年5期)2018-07-09
中国工程咨询(2016年9期)2016-02-13
应用海洋学学报(2015年3期)2015-11-22
应用海洋学学报(2015年3期)2015-11-22
应用海洋学学报(2015年1期)2015-11-22
西安建筑科技大学学报(自然科学版)(2014年2期)2014-11-12
中国海洋大学学报(自然科学版)(2014年12期)2014-02-28
中国海洋大学学报(自然科学版)(2014年12期)2014-02-28
