
南京近郊麻栎林冠层蒸腾时间变化及降温效应
2015-12-29 11:15张金池谢德晋庄家尧邵永昌张水锋
浙江农林大学学报 2015年4期
刘 鑫,张金池,谢德晋,庄家尧,邵永昌,张水锋,2
(1.南京林业大学 水土保持与生态修复重点实验室,江苏 南京 210037; 2.南京森林警察学院,江苏南京210023)
南京近郊麻栎林冠层蒸腾时间变化及降温效应
刘 鑫1,张金池1,谢德晋1,庄家尧1,邵永昌1,张水锋1,2
(1.南京林业大学 水土保持与生态修复重点实验室,江苏 南京 210037; 2.南京森林警察学院,江苏南京210023)
2012年9月和10月及2013年5-8月,利用树干液流仪和小气象站对南京近郊东善桥林场的麻栎Quercus acutissima树干液流速率及环境因子进行连续观测,分析麻栎林冠层蒸腾量及其对周边环境的降温效应。结果表明:①各月麻栎液流密度均在13:00左右达到最大值,其中8月峰值最高(18.78 g·cm-2·h-1),6月峰值最低(13.49 g·cm-2· h-1);②5-10月麻栎林冠层蒸腾总量为237.52 mm,7月冠层蒸腾量最高(50.46 mm),10月冠层蒸腾量最低(29.86 mm);③7月的蒸腾耗能量与太阳辐射能量值最高,分别为154.97 MJ·m-2,489.65 MJ·m-2,8月冠层蒸腾耗能系数最高(35.82%),10月的冠层蒸腾耗能系数最低(25.76%);④麻栎林7月冠层蒸腾降温值最高(3.07℃),生长期平均可降温(2.35±0.53)℃。图6表1参28
森林水文学;麻栎林;冠层蒸腾;降温
森林具有显著的调控水源与调节气候的作用,森林植被通过蒸腾作用向大气中输送大量的水分,增加空气湿度,同时吸收周围空气中的热量以降低空气温度。随着社会发展,全球变暖一直都是人们广泛关注的热点[1-2],从1990年至今政府间气候变化专门委员会(IPCC)已经发布了4次气候变化问题的综合性评估报告[3-6],城市热岛效应是全球变暖内容中的重要部分,而植被在调节热岛效应里扮演着重要角色[7-9]。目前,热技术被广泛应用于研究植被蒸腾作用,多数学者通过热技术测定树干液流来研究植被对水资源的输出调控,如Falko等[10],Liu等[11]和Hentschel等[12]分别研究了挪威云杉Picea abies,砂梨Pyrus pyrifolia与山毛榉Fagus sylvatica的液流变化特征。植被在蒸腾的同时吸收了热能,降低了空气温度。Zhu等[13]得出木荷Schima superba通过蒸腾作用可以降低10 m3空气柱4.5℃左右的温度,Nakazato等[14]研究得出黄金葛Epipremnum aureum通过蒸腾可以降低约1.0℃。本研究采用热扩散探针,对南京城郊典型树种麻栎Quercus acutissima林树干液流速率进行连续定位观测,并结合各环境因子的同步观测,实现对各月冠层蒸腾量的准确估算,并分析对周边环境的降温效应,为该区域树种选择及缓解城市热岛效应提供基础资料。
1 试验区概况
试验区位于南京市近郊国有东善桥林场铜山分场(31°35′~31°39′N,118°50′~118°52′E),属亚热带季风气候区,生长季节日间平均气温为18.06℃,无霜期229.0 d,年日照时间2 199.5 h。区内气候温和湿润,四季分明,雨热资源比较丰富,生长季长,年平均降水量为1 100.0 mm,时间分布上多存在2个多雨期:一是春夏之交的梅雨,二是夏季的台风雨,为中国雨期最长的地区之一,十分有利于农作物和林木的生长。地形以丘陵为主,海拔为38~388 m,森林类型以杉木Cunninghamia lanceolata,马尾松Pinus massoniana,麻栎和毛竹Phyllostachys edulis为主。
2 材料与方法
2.1 试验材料
试验区的麻栎林,平均树高为13.8 m,平均胸径为25.8 cm,郁闭度为0.81,林分密度为425株· hm-2。平均坡度为18°。土壤类型均为黄棕壤。麻栎林林下植被为悬钩子Rubus corchorifolius和山胡椒Lindera glauca。选择6株干形通直,冠型良好,生长健康的样木。具体特征见表1。

表1 麻栎样树的基本特征Table 1 Features of Quercus acutissima sample trees
2.2 试验方法
2.2.1 探针式树干液流仪安装与测定 首先,在树干距地1.3 m处用刀将树皮清除掉,形成1个宽4 cm,高10 cm的矩形框,将钻模平放在矩形框上,使用电钻打2个圆孔,其距离为4 cm,然后将FLGS-TDP热扩散探针安装进去,用橡皮泥封住接口处,并用固定泡沫和胶带固定探针。之后,用防辐射护罩将探针所在位置完全包裹,最后将探针与主机相连。隔10 s获取1次数据,并存储1 h的平均值。比较6株样木液流速率变化特征,可以明显看出其变化趋势基本相同,主要差异在于其值的大小。这主要是由于胸径等因素的影响,因此,此次对6株样树测定的液流速率数据进行加权平均处理。
树干液流速率计算公式[15-16]如下:

式(1)~式(3)中:α为参数;dTM为上下探针之间的最大昼夜温差,℃;dT为瞬时温差,℃;v为平均液流速率,cm·s-1;As为树干边材面积,cm2;vs为树干液流速率,kg·h-1。
每个样地选择除测定样木之外的20株树木进行边材面积的直接读取,利用生长锥和胸径尺获取心材、边材与树皮的厚度,进而求出边材面积。通过相关性分析得出,麻栎胸径与边材面积存在着显著的幂函数关系。公式如下:
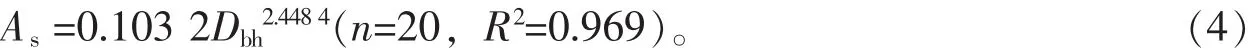
式(4)中:Dbn为胸径,cm。
2.2.2 各环境因子测定 使用小气象站进行各环境因子的连续测定,存储30 min的均值。此次测定的环境因子有太阳辐射(Rs,W·m-2),空气相对湿度(RHa,%),空气温度(Ta,℃),土壤体积含水率(SMC,%),土壤温度(Ts,℃)。其中空气温湿度分别测定了林内和林外值,土壤体积含水率、土壤温度为林内土壤测定值,太阳辐射为林外测定值。饱和水汽压差(VPD)是当时温度下空气中的饱和水汽压(es)与实际水汽压(ea)之间的差值,由以下公式[17]计算得出:

式(5)~式(6)中:VPD为饱和水汽压差,kPa;RHa为空气相对湿度,%;Ta为空气温度,℃。
2.2.3 能量计算 蒸腾耗能量是指单位面积白天冠层蒸腾作用所消耗的能量,计算时段为太阳辐射不为0时段,由以下公式计算得出:

式(7)中:Etr为单位面积蒸腾耗能量,kJ·m-2;Q为液流量,kg;λ为汽化潜热,气温为Ta时,λ=2 498.9-2.33Ta,kJ·kg-1;ρ为麻栎林林分密度,值为425株·hm-2。太阳辐射能量是指单位面积太阳辐射所释放的能量,由以下公式计算:

式(8)中:Esr为单位面积太阳辐射能量,kJ·m-2;Rs为太阳辐射,W·m-2;t为时间,s。耗能系数是指麻栎林日间蒸腾耗能量占太阳辐射能量的百分比,计算公式如下:

2.2.4 蒸腾降温计算 考虑到空气的湍流、对流和辐射作用,空气与叶面之间及空气微气团之间不断地进行热量扩散和交换,取底面积为10 m2,厚度为100 m的空气柱[13]作为计算单元,在此空气柱体中,因植物蒸腾消耗热量是取自于周围1 000 m3的空气柱体,故使气柱温度下降。气温下降值用下式[13]表示:

式(10)中:T为蒸腾所降低的温度,℃;ρair为空气密度(ρ=1.283 7-0.003 9Ta,kg·m-3);c为空气的比热容,1 005 J·kg-1·℃-1;V为1 000 m3空气柱体体积。
2.2.5 数据处理 使用Excel 2007和SPSS 19.0进行数据处理和统计分析。
3 结果与分析
3.1 研究区环境因子
图1描述了2012年9-10月及2013年5-8月麻栎生长期内环境因子的月变化。图1A显示7月降水量较大,10月降水量较低,生长期总降水量为778.6 mm。图1C显示了生长期内林外空气温度与空气相对湿度的月变化。8月空气温度较高,10月空气温度较低,分别为30.3℃和17.0℃;6月的空气相对湿度较大,8月的空气相对湿度较低,分别为89.4%和79.0%,生长期内平均空气温湿度分别为23.7℃和82.7%。图1B显示8月饱和水汽压差较大,值为0.854 kPa,6月饱和水汽压差较低,值为0.319 kPa,平均饱和水汽压差为0.524 kPa。土壤含水率变化与雨量变化较一致,6月平均土壤含水率较高,值为17.6%,8月平均土壤含水率较低。主要是因为空气温度高,土壤蒸发与植被蒸腾消耗水量较高,因此即使雨量高于10月,但土壤含水率则较低。
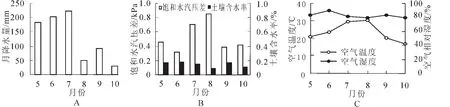
图1 环境因子月变化Figure 1 Monthly variations of environment factors
3.2 液流密度日变化
如图2所示:各月麻栎液流密度具有显著的昼夜变化规律,各月均在13:00左右达到最大值,5月至10月最大值分别为13.69,13.49,18.71,18.78,15.86,14.99 g·cm-2·h-1,其中8月峰值最高,6月峰值最低。6月降水量仅次于7月份,但其降水历时长,空气相对湿度高于其他月份,而7月降水历时较短,降水量集中,因此6月虽然降水量小于7月,但液流密度最低。6月与7月液流密度在6:00开始增加,其他月份从7:00开始增加。

图2 液流密度日变化Figure 2 Daily change of sap flux density
3.3 冠层蒸腾量月变化
图3描述了麻栎林冠层蒸腾量月变化特征。7月冠层蒸腾量较高,值为50.46 mm,10月冠层蒸腾量较低,值为29.86 mm。8月虽然饱和水汽压差、空气温度均高于7月,但其冠层蒸腾量低于7月。分析环境因素可以得出其可能是因为7月降水量和土壤含水率高于8月份。5-10月麻栎林冠层蒸腾量为237.52 mm,日均蒸腾量为1.29 mm。
3.4 冠层蒸腾降温效应
通过公式(6)~式(8)分别计算了麻栎林冠层蒸腾耗能量、太阳辐射能量及冠层蒸腾耗能系数。图4描述了2012年9月和10月及2013年5-8月的麻栎林单位面积冠层蒸腾耗能量、太阳辐射能量与耗能系数的月变化。由图4可以看出:7月的蒸腾耗能量与太阳辐射能量值较高,分别为154.97 MJ·m-2和489.65 MJ·m-2,其次是8月,10月的蒸腾耗能量较低,值为84.74 MJ·m-2,6月的太阳辐射能较低,值为0.69 MJ·m-2。从5-10月,耗能系数先升后降,在8月达到峰值,值为35.82%,10月的耗能系数最低,为25.76%,平均冠层蒸腾耗能系数为30.13%。
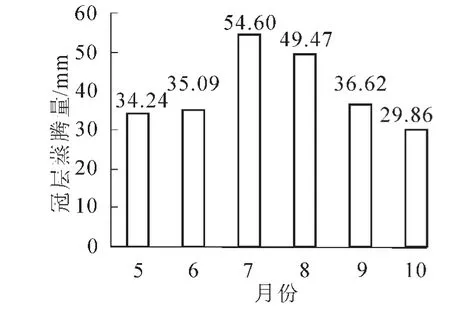
图3 林分冠层蒸腾量月变化Figure 3 Monthly change of stand canopy transpiration
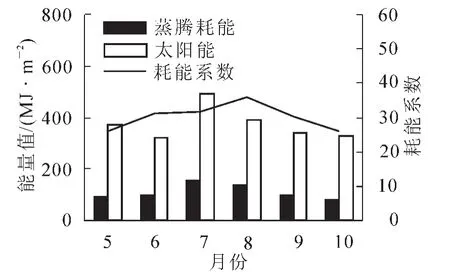
图4 月尺度下蒸腾耗能、太阳能及耗能系数变化特征Figure 4 Changes of transpiration heat flux,total solar radiation and consume energy coefficient at the temporal scale of a month
通过式(6)和式(9)算出日间麻栎林冠层1 000 m3空气柱内每小时可以降低的空气温度。图5分别描述了2012年9月、10月与2013年5-8月日尺度(图5A)与月尺度(图5B)的麻栎林冠层日间蒸腾平均每小时降低的空气温度。另外,由于数据缺乏,仅分析了2012年9月和10月麻栎林林内与林外的温度差值。
随着冠层蒸腾量增加,麻栎林冠层每天约在6:00-7:00左右发挥其降温效应。由图5A可以看出:5月13日的冠层蒸腾降温最高,达到3.95℃,其次是7月13日,降温3.88℃,9月与10月冠层蒸腾降温与麻栎林内外温差之间变化较一致,相关性达到0.70(F=105.9,P=0.000<0.01)。由图5B可以得出,7月的降温最高(3.07℃),显著高于5月、6月、10月降温值,与8月、9月降温值之间差异不显著,其次是8月(2.88℃),10月的降温最低(1.89℃),5月、6月、8月、9月、10月蒸腾降温值之间差异不显著。在此次的试验区内,麻栎林生长期内通过冠层蒸腾作用日间平均降温为(2.35±0.53)℃·h-1。

图5 麻栎林冠层1 000 m3空气柱蒸腾降温效应Figure 5 Cooling effect of canopy transpiration of Quercus acutissima
3.5 冠层蒸腾降温影响因素分析
冠层蒸腾降温效应随各月份蒸腾量大小而变化,同时受到各月份环境因子的影响,如图6所示,同时可以说明蒸腾量与各环境因子的关系。比较蒸腾降温值与各环境因子的关系可以看出:其与空气温度(P=0.04<0.05)、饱和水汽压差(P=0.04<0.05)具有显著的正相关关系,与叶面积指数存在正相关关系,但不显著(P=0.09>0.05),与土壤含水率、空气相对湿度和降雨之间的关系不显著。
4 讨论
众所周知,森林具有涵养水源、保持水土的作用,但森林同时通过蒸腾作用消耗大量的水分,不合理的造林设计将会对地区的水资源利用造成威胁。本研究得出5-10月期间南京城郊麻栎林冠层蒸腾总量为237.52 mm,约占该时段降水量的43.53%,其中,7月冠层蒸腾量较高(50.46 mm),10月冠层蒸腾量较低,日均蒸腾量为1.29 mm,略低于阮宏华等[18]估算的苏南区域6-8月次生栓皮栎Quercus variabilis日均蒸腾量,其结果为1.69 mm。另外,分别总结中国华北、西北、华南、西南区域关于林分蒸腾耗水量的研究进展得出,不同区域不同林分蒸腾耗水存在一定的差异性,如:肖辉杰等[19]测定得出6-9月北京山区侧柏Platycladus orientalis林、油松Pinus tabuliformis林日均蒸腾量分别为2.63和1.58 mm;王鹤松等[20]测定华北石质山区杜仲Eucommia ulmoides人工林4-9月日均蒸腾量约为1.31 mm;刘彩凤等[21]研究山西黄土丘陵沟壑区油松、刺槐Robinia pseudoacacia混交林蒸腾耗水情况得出7-10月日均蒸腾量为0.81 mm;田风霞等[22]利用改进的Penman-Monteith修正式估算出祁连山区云杉Picea asperata 5-9月平均日蒸腾量为0.97 mm;赵平等[23]测定华南丘陵区成熟马占相思Acacia mangium林6-7月日均蒸腾量约为1.26 mm;Huang等[24]研究广西桂林青冈栎Cyclobalanopsis glauca林蒸腾量得出7月日均值5.1 mm。比较以上各区域的林分蒸腾量可以看出,西北区域的降水量为365~600 mm,并且多集中在6-8月,日均蒸腾量普遍低于1.00 mm,降水严重影响该区域的蒸腾特征;降水较为充分的华北地区,降水量为600~700 mm,蒸腾耗水量显著高于西北区域,而降水丰沛的华南地区各林分的蒸腾量差异性较大,年均降水量为1 500~1 900 mm,各月最高日蒸腾量为1.26~5.10 mm,间接可以说明在该区域蒸腾耗水不受到降水量的限制。本研究区域位于华东范围内,降水较为充沛,但降水量是否限制其蒸腾量还不能通过此阶段的比较得出。
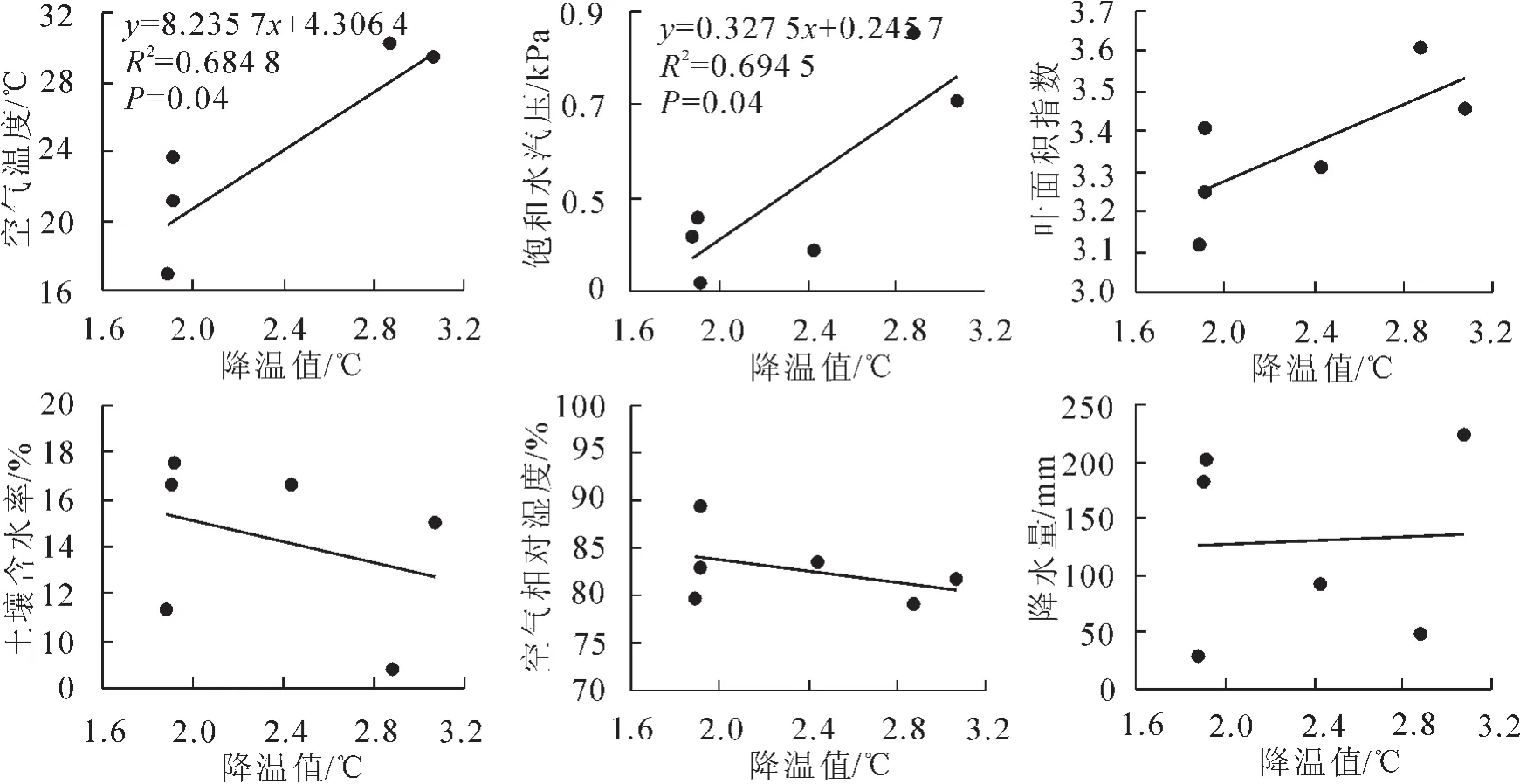
图6 蒸腾降温值与环境因子的关系Figure 6 The relationship between temperature reduction value and environment factors
图6 得出蒸腾降温值与土壤含水率、空气相对湿度和降水之间的关系不显著,说明降水量不是限制该区麻栎蒸腾及降温的环境因子,同华南地区的分析结论一致。蒸腾量与土壤含水率、空气相对湿度之间大致呈现负相关关系,主要因为其随着降水而增加,降水时期由于太阳辐射、空气温度降低,蒸腾量显著下降。另外,比较6月和7月各环境因子与耗水量变化特征得出,6月空气温度、降水量与7月相差较小,如图1,但其蒸腾量及其降温值却显著低于7月,如图5,分析其主要原因是由于6月降水次数显著多于7月,7月降水较集中导致的,这与赵平等[23]关于马占相思林的研究结果相一致。综合以上比较分析得出,降水量虽然不限制该区域麻栎蒸腾耗水,但降水过程历时较长,会直接影响饱和水汽压差、太阳辐射、空气温度等因素,进而降低某一时间段的蒸腾耗水量。
生长的树木是天然的冷却器,而水则就是冷却剂[25]。苏泳娴等[26]利用遥感影像广州城区对周边公园降温效应指出城市绿地通过植被的蒸腾、蒸散、光合作用降低温度增加湿度,以缓解城市热岛效应,张艳丽等[27]利用光合测定系统测定叶片蒸腾速率进而研究成都市主要绿化树种的降温效应,研究指出垂柳Salix babylonica的降温能力最强,日均降温0.33℃,其次是水杉Metasequoia glyptostroboides和山杜英Elaeocarpus sylvestris。欧阳旭等[28]通过观测林内外小气候变化特征得出混交林内年均气温与地温分别比空旷地低2.3℃和4.0℃,表现出良好的降温效应,Zhu等[13]通过测定蒸腾速率得出木荷通过蒸腾作用可以降低10 m3空气柱4.5℃左右的温度,比较不同方法及不同地区及树种均得出蒸腾作用具有显著的降温生态效应,但定量研究森林蒸腾对周围环境的降温效应还是很少的,而且随着全球气候变暖、城市热岛效应及地区水资源紧缺等问题的加剧,如何定量准确地评价地区森林蒸腾的降温值及其对城市环境的影响将变得更加重要。本研究通过准确测定冠层蒸腾量,分析得出南京城郊麻栎林冠层7月份日间蒸腾降温值最高,达到3.07℃,生长期平均可降温(2.35±0.53)℃。
本研究目前只针对麻栎进行了为期1 a的研究,在未来还需增加研究树种,并选择不同密度、不同林龄样地的测定,并进行多年的持续观测,最终实现对该区域林木冠层蒸腾量及其降温效应的准确估测与评价。
[1] LOBELL D B,HAMMER G L,McLEAN G,et al.The critical role of extreme heat for maize production in the United States[J].Nat Clim Change,2013,3(5):497-501.
[2] GAO Xuejie,ZHAO Zongci,DING Yihui,et al.Climate change due to greenhouse effects in China as simulated by a regional climate model[J].Adv Atmos Sci,2001,18(6):1224-1230.
[3] IPCC.Climate Change 1990:the IPCC Scientific Assessment[M].Cambridge:Cambridge University Press,1990.
[4] IPCC.Climate Change 1995:the Science of Climate Change:Contribution of Working Group I to the Second Assessment Report of the Inter Governmental Panel on Climate Change[M].Cambridge:Cambridge University Press,1996.
[5] IPCC.Climate Change 2001:the Scientific Basis:Contribution of Working Group I to the Third Assessment Report of the Intergovernmental Panel on Climate Change[M].Cambridge:Cambridge University Press,2001.
[6] IPCC.Climate Change 2007:the Physical Science Basis:Contribution of Working Group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change[M].Cambridge:Cambridge University Press,2007.
[7] OKE T R,CROWTHER J M,McNAUGHTON K G,et al.The micrometeorology of the urban forest[J].Philos Trans Royal Soc Ser B Biol Sci,1989,324(223):335-349.
[8] PETERS E B,McFADDEN J P,MONTGOMERY R A.Biological and environmental controls on tree transpiration in a suburban landscape[J].J Geophys Res Biogeosci,2010,115(G4):G04006.DOI:10.1029/2009JG001266.
[9] GILLA S E,RAHMANB M A,HANADLEY J F,et al.Modeling water stress to urban amenity grass in Manchester UK under climate change and its potential impacts in reducing urban cooling[J].Urban For&Urban Green,2013,12(3):350-358.
[10] CLAUSNITZER F,KÖSTNER B,SCHWÄRZEL K,et al.Relationships between canopy transpiration,atmospheric conditions and soil water availability:analyses of long-term sap-flow measurements in an old Norway spruce forest at the Ore Mountains/Germany[J].Agric For Meteorol,2011,151(8):1023-1034.
[11] LIU Xiaozhi,KANG Shaozhong,LI Fusheng.Simulation of artifcial neural network model for trunk sap flow of Pyrus pyrifolia and its comparison with multiple-linear regression[J].Agric Water Manage,2009,96(6):939-945.
[12] HENTSCHEL R,BITTNERB S,JANOTT M,et al.Simulation of stand transpiration based on a xylem water flow model for individual trees[J].Agric For Meteorol,2013,182:31-42.
[13] ZHU Liwei,ZHAO Ping.Temporal variation in sap-flux-scaled transpiration and cooling effect of a subtropical Schima superba plantation in the urban Area of Guangzhou[J].J Integr Agric,2013,12(8):1350-1356.
[14] NAKAZATO T,INAGAKI T.Analysis of plant function as bio-thermal-conditioner using Pothos (Epipremnum aureum)[J].Thermochimica Acta,2012,532:49-55.
[15] GRANIER A.A new method of sap flow measurement in tree stems[J].Ann For Sci,42(2):193-200.
[16] GRANIER A.Evaluation of transpiration in a Douglas fir stand by means of sap flow measurements[J].Tree Physiol,1987,3(4):309-320.
[17] CAMPBELL G S,NORMAN J M.An Introduction to Environmental Biophysics[M].2nd ed.New York:Springer-Verlag,1998:5-9.
[18] 阮宏华,郑阿宝,钟育谦.次生栎林蒸腾强度与蒸腾量的研究[J].南京林业大学学报,1999,23(4):32-35.
RUAN Honghua,ZHENG Abao,ZHONG Yuqian.A study on the transpiration intensity and total transpiration calculation of the secondary oak forest[J].J Nanjing For Univ,1999,23(4):32-35.
[19] 肖辉杰,余新晓,贾瑞燕,等.北京山区典型人工林的耗水规律[J].中国水土保持科学,2014,12(2):72-77.
XIAO Huijie,YU Xinxiao,JIA Ruiyan,et al.Water consumption of typical artificial forests in Beijing mountainous area[J].Sci Soil Water Conserv,2014,12(2):72-77.
[20] 王鹤松,张劲松,孟平,等.华北石质山区杜仲人工林蒸腾特征及水分供求关系[J].林业科学研究,2008,21(4):475-480.
WANG Hesong,ZHANG Jinsong,MENG Ping,et al.Transpiration variation and relationship between supply and demand of water for Eucommia ulmoides plantations in the rocky mountainous area of north China[J].For Res,2008, 21(4):475-480.
[21] 刘彩凤,张志强,郭军庭,等.晋西黄土丘陵沟壑区油松刺槐混交林蒸腾耗水[J].中国水土保持科学,2010,8(5):42-48.
LIU Caifeng,ZHANG Zhiqiang,GUO Junting,et al.Transpiration of a Pinus tabulaeformis and Robinia pseudoacacia mixed forest in hilly-gully region of the Loess Plateau,West Shanxi Province[J].Sci Soil Water Conserv,2010,8(5):42-48.
[22] 田风霞,赵传燕,冯兆东.祁连山区青海云杉林蒸腾耗水估算[J].生态学报,2011,31(9):2383-2391.
TIAN Fengxia,ZHAO Chuanyan,FENG Zhaodong.Model-based estimation of the canopy transpiration of Qinghai spruce(Picea crassifolia)forest in the Qilian Mountains[J].Acta Ecol Sin,2011,31(9):2383-2391.
[23] 赵平,邹绿柳,饶兴权,等.成熟马占相思林的蒸腾耗水及年际变化[J].生态学报,2011,31(20):6038-6048.
ZHAO Ping,ZOU Lüliu,RAO Xingquan,et al.Water consumption and annual variation of transpiration in mature A-cacia mangium plantation[J].Acta Ecol Sin,2011,31(20):6038-6048.
[24] HUANG Yuqing,LI Xiankun,ZHANG Zhongfeng,et al.Seasonal changes in Cyclobalanopsis glauca transpiration and canopy stomatal conductance and their dependence on subterranean water and climatic factors in rocky karst terrain[J].J Hydrol,2011,402(1/2):135-143.
[25] PRAŽÁK J,ŠÍR M,TESARRˇM.Estimation of plant transpiration from meteorological data under conditions of sufficient soil moisture[J].J Hydrol,1994,162(3):409-427.
[26] 苏泳娴,黄光庆,陈修治,等.广州市城区公园对周边环境的降温效应[J].生态学报,2010,30(18):4905-4918.
SU Yongxian,HUANG Guangqing,CHEN Xiuzhi,et al.The cooling effect of Guangzhou City parks to surrounding environments[J].Acta Ecol Sin,2010,30(18):4905-4918.
[27] 张艳丽,费世民,李智勇,等.成都市沙河主要绿化树种固碳释氧和降温增湿效益[J].生态学报,2013,33(12):3878-3887.
ZHANG Yanli,FEI Shimin,LI Zhiyong,et al.Carbon sequestration and oxygen release as well as cooling and humidification efficiency of the main greening tree species of Sha River,Chengdu[J].Acta Ecol Sin,2013,33(12):3878-3887.
[28] 欧阳旭,李跃林,张倩媚.鼎湖山针阔叶混交林小气候调节效应[J].生态学杂志,2014,33(3):575-582.
OUYANG Xu,LI Yuelin,ZHANG Qianmei.Characteristics of microclimate in a mixed coniferous and broadleaf forest in Dinghushan Biosphere Reserve[J].Chin J Ecol,2014,33(3):575-582.
Temporal variation for canopy transpiration and its cooling properties in a Quercus acutissima forest of suburban Nanjing
LIU Xin1,ZHANG Jinchi1,XIE Dejin1,ZHUANG Jiayao1,SHAO Yongchang1,ZHANG Shuifeng1,2
(1.Jiangsu Key Laboratory of Soil and Water Conservation and Ecological Restoration,Nanjing Forestry University, Nanjing 210037,Jiangsu,China;2.Nanjing Forest Police College,Nanjing 210023,Jiangsu,China)
Global warming and regional water resources shortage have become the focus of the global.Climate warming affects global hydrological cycles,and tree transpiration also affects climate warming.This study was conducted to analyze canopy transpiration of Quercus acutissima and to determine its cooling effect on surrounding environments.Continuous observation of the sap flow density and environmental factors,such as solar radiation,air temperature,air relative humidity,soil temperature,soil water content,used a sap flow meter and a mini weather station in Nanjing Dongshanqiao Forest Farm.Maximum sap flux density,peak daily sap flux density,transpiration heat flux,solar energy,consumed-energy coefficient,and canopy transpiration cooling was conducted according these data.Six trees of different diameter grade were observed.Results indicated that 1)the maximum sap flux density appeared at 13:00;the greatest peak daily sap flux density was 18.78 g·cm-2·h-1in August,and the lowest was 13.49 g·cm-2·h-1in June.2)Canopy transpiration summed from May to October was 237.52 mm with the highest month being July.3)July also had the greatest transpiration heat flux(154.97 MJ·m-2)and solar energy(489.65 MJ·m-2).The coefficient of consumed-energy for canopy transpiration was highest in August(35.8%)and lowest in October(25.8%).4)Canopy transpiration cooling from Q. acutissima was highest in July(3.07°C),and during the growing season,canopy transpiration reduced the av-erage temperature (2.35±0.53)°C·h-1per 10 m3air.According to the transpiration and its cooling effect,we could choose suitable afforestation design in order to alleviate global warming and heat island effect [Ch,6 fig.1 tab.28 ref.]
forest hydrology;Quercus acutissima forest;canopy transpiration;cooling
S715.4
A
2097-0756(2015)04-0529-08
10.11833/j.issn.2095-0756.2015.04.006
2014-11-20;
2014-12-24
长三角水源区面源污染林业生态修复技术研究项目(201104055-1);国家林业公益性行业重大专项(201104005);江苏高校优势学科建设工程资助项目(PAPD)
刘鑫,博士,从事森林水文研究。E-mail:731845212@qq.com。通信作者:张金池,教授,博士,博士生导师,从事林业生态工程、水土保持与荒漠化防治研究。E-mail:zhang8811@njfu.edu.cn
猜你喜欢
气候与环境研究(2022年4期)2022-08-15
干旱区地理(汉文版)(2022年3期)2022-05-30
中南林业科技大学学报(2021年9期)2021-10-12
环保科技(2020年2期)2020-05-21
浙江农林大学学报(2019年6期)2019-11-13
装备环境工程(2019年9期)2019-10-24
湖南大学学报·自然科学版(2015年10期)2015-11-30
建筑科学与工程学报(2015年3期)2015-07-25
小小说月刊·下半月(2015年11期)2015-05-14
储能科学与技术(2014年5期)2014-02-27
