
小麦分子标记辅助育种研究进展
2015-12-25 12:21:54于庆祥雷小利马海财
甘肃农业科技 2015年6期
于庆祥,雷小利,张 静,马海财
(宁夏回族自治区固原市种子工作站,宁夏 固原 756000)
小麦是世界上约35%人口的主粮,随着人口的不断增长,未来我们将面临着小麦产量不能满足人口增长需求的严峻挑战[1]。按照目前人口的增速,在未来几十年粮食产量需要翻番[2],但近10年小麦产量未能持续增长,仅是适度增长,引起了全世界小麦领域专家的关注[3]。尽管社会经济因素和基础设施缺乏在很大程度上阻碍了小麦产量和品质上的提高,但最主要的瓶颈是没有进一步利用遗传改良突破传统的育种模式,这在很大程度上是由于缺乏最复杂性状的遗传结构知识、在一个简单的育种计划中对某些性状中一些复杂的数量性状没有能力进行有效选择所致。因此,小麦育种家正在寻求可选择的技术来补充传统的小麦育种所面临困难。近年来,作为这些技术中的分子标记辅助选择,已经在世界上许多国家作为小麦常规育种的补充被越来越多的育种家采用[3],一些被开发的且与控制主要经济性状的数量性状位点(Quantitative trait locus,QTL)相关联的标记,已经被应用到分子标记辅助选择中(表1),并已取得良好的效果[4]。分子标记辅助育种初期大多数被用到一些单基因的难以评价的性状中,而随着技术不断的发展和成熟,育种家开始把分子标记轮回选择和全基因组选择等用到更适合复杂的数量性状,例如控制产量、分蘖等多基因的数量性状[5]。我们主要阐述了小麦分子标记辅助选择育种技术以及该技术当前利用的现状和局限性。
1 分子标记辅助育种的诞生
分子标记辅助选择技术最早由Sax于1923年提出[6]。小麦分子标记辅助育种技术是指在小麦上利用与目标性状紧密连锁的分子标记对目标性状加以间接的选择,在选育的早代就能够对目标基因进行准确、稳定的选择,并且能够克服一些隐性基因再度利用时识别的困难,从而加快小麦育种的进程,提高育种的效率,选育出抗病、优质和高产的新品种或新品系。与形态标记、细胞标记、生化标记育种相比较,分子标记辅助育种的优点一是在植物体的多个组织及生育阶段均可检测到,不受时空限制,数量多,遍及整个基因组。有许多标记表现为共显性,能够鉴别基因型纯合与否,提供完整的基因型。二是在标记小麦抗病害基因方面,分子标记可以在更深层次上揭示小麦抗病遗传机制。通过找到与抗病基因紧密连锁的分子标记,不但能在遗传背景不同的育种材料中特异性的检测目的基因,而且可以在任何一个生育阶段同时对多个抗性基因进行筛选,这为了解抗源和抗病品种中所含有的抗性基因提供了更为迅速、稳定和可靠的方法。三是为了提高选择效率,越来越多的分子标记被用到回交育种了,主要在前景选择(主要跟踪目标基因的等位基因)、背景选择(回交育种计划中主要进行轮回亲本基因组的选择)和重组体的筛选(用目标基因和发生在目标位点和标记两侧的重组事件来鉴定回交后代,而减少连锁累赘)层面进行选择。截止目前已开发和定位的简单遗传性状位点见表2,定位的基因或QTL进行聚合的结果见表3。

表1 小麦中定位到的与重要性状相关联的QTLs(引GyanaRanjan Rout,2013)

表2 小麦中用各种分子标记定位到的简单遗传性状位点(引GyanaRanjan Rout,2013)
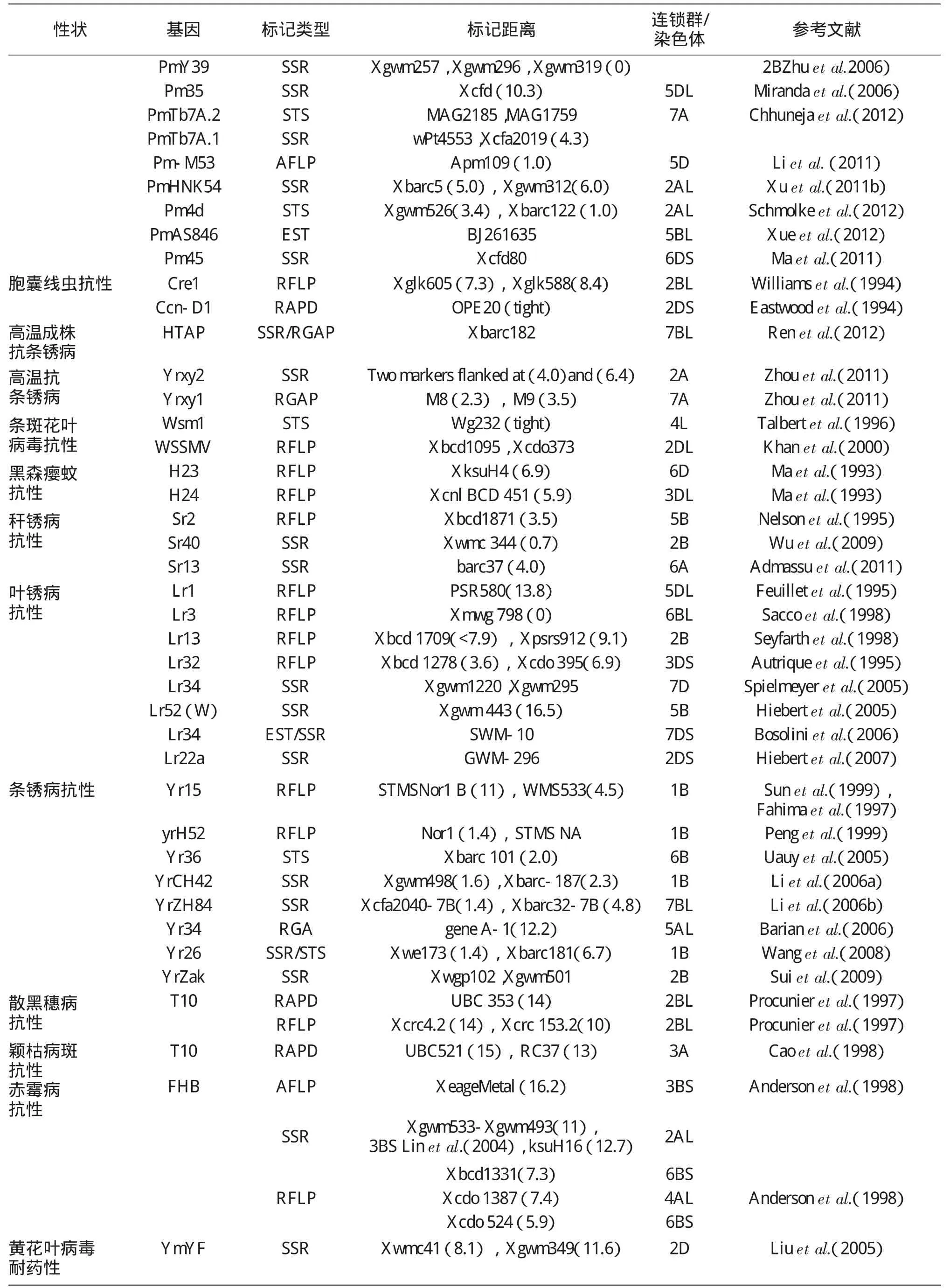
表2 续
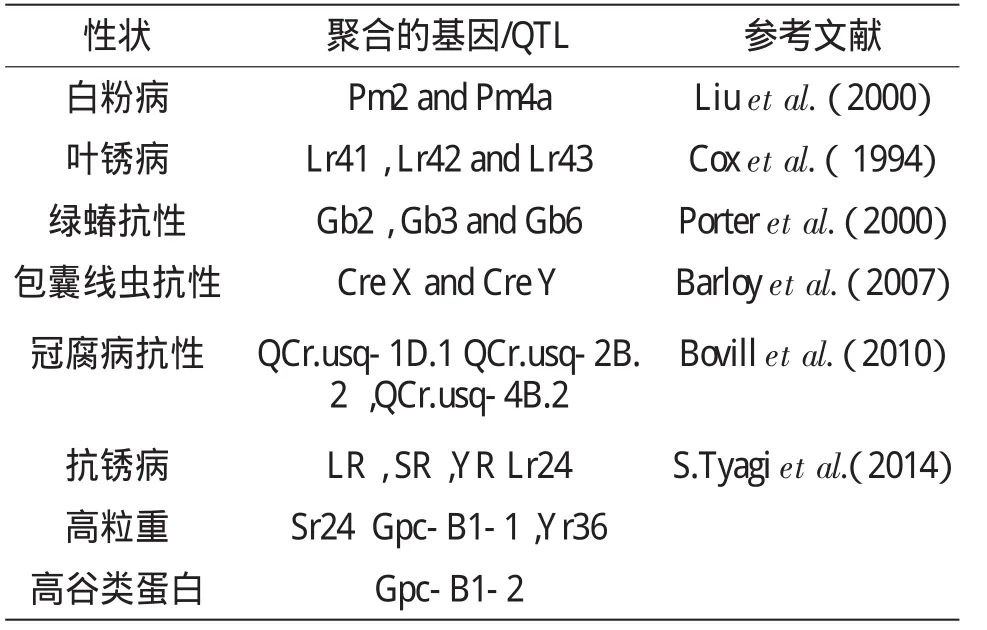
表3 小麦中通过标记辅助聚合的基因和QTL(部分引 GyanaRanjan Rout,2013)
2 分子标记辅助技术在小麦育种中的应用
过去的10 a里,利用分子标记对小麦进行了遗传多样性的调查,构建了小麦分子遗传图谱,对小麦的某些质量性状和数量性状(QTLs)进行了定位,同时利用分子标记辅助技术进行了小麦商用品种和品系的选育工作(表4)。虽然DNA标记在作物育种中有很多用途,但是育种中最有前景的是标记辅助选择育种。目前利用标记辅助选择育种已经取得显著效果[7]。

表4 利用标记辅助回交育种所选育的新品种和新品系(部分引GyanaRanjan Rout,2013)
2.1 国外应用情况
在国外,小麦分子标记辅助育种主要集中在澳大利亚、美国、加拿大、墨西哥、阿根廷、乌克兰、法国、土耳其、俄罗斯和印度等国家。标记辅助小麦育种,无论是在国家的科研单位和高校,还是在私营公司都已经产生了积极效果[8]。
1996年,澳大利亚开始了国家小麦分子标记计划(NWMMP),2001年与大麦计划合并,将其命名为Australian Winter Cereal Molecular Marker Program’(AWCMMP)。这2个计划的重要研发资金由为了在小麦育种计划中开发和利用分子标记的谷类研发公司(GRDC)提供。该计划涉及到小麦20个不同性状的改良,包括抗病性、抗非生物胁迫和质量性状等。澳大利亚联邦科学与工业研究组织植物工业部植物工程小麦改良计划用MAS来筛选一个提高蒸腾效率的QTL[9],在澳大利亚的西部,小麦育种计划有约42个性状或基因,包括条锈病抗性的范围已经被选择用到通过MAS进行品种的选育和种植资源创新[9]。
在美国,一个名叫“将基因组学引进小麦领域”的计划于2001年开始,形成了一个国家小麦MAS团体,类似于澳大利亚的“NWMMP”。该项目延续了4 a(2001—2005年),后来延伸为在小麦农业合作计划下的应用小麦基因组学或叫小麦合作农业计划。2个项目都由美国农业部提供资金资助,同时与加利福尼亚州立大学戴维斯分校的Jorge Dubcovsky教授合作[10]。把基因组引入小麦计划的目标是用MABS整合27个不同的病虫害抗性基因和20个在面包制作和匹萨饼的品质方面有效的等位基因聚合到180个适应美国小麦原产地的品系中。
在加拿大,发起了一个重要计划,即整合分子标记技术到他们的常规小麦育种计划中去。在该计划的最初阶段,分子标记与多个农艺性状相关的特性,如抗病性、抗虫性和面包品质以及淀粉的特异性和穗发芽被鉴定。第二个阶段和第一个阶段被鉴定的标记被用到转移目标性状到栽培品种中去,被用来通过MAS跟踪不同基因,包括Fhb1, Fhb2(赤霉病)、Lr34/Yr18(叶锈病)、Sr30,Sr24/Lr24(根锈病)、Bt10(腥黑穗病)、GPC-B1(谷类蛋白)含量)、Sm-1(麦黄吸浆虫)、and Utd1(散黑穗病)。这项工程的实施,开发了2个品种,即拥有高蛋白质含量基因(GPC-B1)的Lillian和携带香橙花小麦蚊抗性基因(Sm1)的Goodeve。
在欧洲,多数小麦育种是在商业部门进行[11]。分子标记已经被广泛应用,用大于50个不同位点的标记辅助的计划已经进行多年。其中,俄罗斯在抗病育种方面已取得显著的效果,例如抗病基因 Lr、Yr、Sr、Rht、Ppd、Vrn和 Vp等的定位和应用[12]。法国、土耳其等国在小麦分子标记辅助育种方面的进步也很快[13]。
在印度,基因聚合方面已经取得了显著的效果。基因聚合是指将一个或多个性状的多个QTL或基因渗入一个在该性状上有缺陷的优良品系或品种的过程。例如S.Tyagi等将抗条锈病、抗穗发芽和高粒重等4个性状的8个QTL聚合到一个新品系PBW343中,并应用到新品种选育中[14]。
2.2 国内应用情况
我国利用分子标记技术辅助育种选育了一批新的抗病品系和种质,如中国科学院遗传与生物发育研究所农业资源研究中心利用分子标记辅助选择选育出了抗条锈病和白粉病的新品系0-123-1-1[15],南京农业大学应用植物基因组实验室精细定位到了抗白粉病基因pm45、抗赤霉病基因Fhb5和一些产量性状有关的QTL并已经提供给育种者进行育种[16-17];江苏省农业科学院农业生物遗传生理研究所筛选得到2份高抗赤霉病材料,研制出抗赤性超过“苏麦3号”的新抗源,育成了70多个优质、高产、抗赤霉病新品系[18];张海泉等将粗山羊草[Aegilop s tauschii(Coss.)Schmal]Y189中鉴定出1个显性抗小麦白粉病基因,暂定名为PmAeY2,PmAeY2是一个新的抗白粉病基因,可用于分子标记辅助选择育种[19];中国农业科学院将Bdv2、Pm-X转育到“中麦16”、“宛7107”和优质小麦“中优9507”、“郑州9023”中;西北农林科技大学利用分子标记将中国小麦抗赤霉病基因(Yr5,Yr9,Yr10,Yr15,Yr17,Yr18,Yr26)的抗病效果进行了全面评价(Zeng Q.D.et al,2014);山东农业大学与中国农业科学院合作实现了小麦一粗山羊草双二倍体抗白粉病基因定位及其遗传转移[20]。欧巧明等采用优化的花粉管通道法将高粱(Sorghum bicolor L.)基因组DNA导入高感条锈病、籽粒粉质的稳定小麦品系,并借助幼胚培养挽救加代、早代变异筛选等技术,经多代连续选择优良变异系,获得2个稳定的优良变异新品系[21]。
3 存在的问题
3.1 多态性或辨别的标记
为了增加选择效果,育种者应有与感兴趣的基因紧密连锁的标记。所选的标记群体有时候与想要的种植资源不符。认识到的多态性常常使用价值小,在大多数情况下,便利可用的标记不适合育种计划的筛选。低水平的标记多态性在多数育种计划中是需要解决的问题,因为种植资源常常建立在窄的基因库上。这种现象在D基因组中尤为重要[4]。
3.2 标记辅助选择育种的成本
虽然最近转向 SNP和DArT(diversity array technology)平台将帮助解除成本限制,然而标记实验的成本仍然是一个焦点。随着基因组测序工作的完成,分子标记育种的成本将会有逐渐降低的趋势。
3.3 基因组结构
小麦基因组相对比较复杂,详细的了解小麦基因组中目标性状的遗传学机理对在育种计划中有效应用MAS很关键,而且小麦改良中决定很多重要的性状QTL或基因出现在外源片段上,因此在分离策略中加强这些外源片断的渗透和重组尤为重要。这就需要一些管理大片段的方法来处理那些在互相排斥中连锁的性状。
3.4 标记辅助选择的位点数量
在育种计划中,寻求扩大分子标记的应用时,处理大量的位点同样也是一个问题。当用标记跟踪的位点多于60个时,在很大程度上增加了育种的复杂性。当保持群体大小在育种资源的范围内,多个性状可以被渗入同一个品系中。处理这种问题的策略需要进一步研究和探索,同时,有些农艺性状,例如产量等性状由多个数量性状QTL决定[22],选择起来比较困难。
4 结束语
在分子标记产生以来,标记性状关联性应用方面和标记辅助选择在小麦育种方面都取得了显著的成就,而且,在标记辅助选择的帮助下,大量的新品系和种质被选育,这些种质或品系被全世界的育种家利用。然而通过标记辅助选择改良的大多数性状是简单的性状,包括病虫害抗性和质量性状等,而利用分子标记辅助选择育种技术改良多基因的数量性状刚刚开始。这些性状将是未来育种者利用MAS要去努力的聚焦。小麦育种未来成功的一个关键组成是在于通过应用分子技术所进行的策略和创新。因此,育种者需要直接精细定位目标基因和用来分析重要的种植资源。在育种策略上,利用分子标记创新将在未来分子标记育种中成为关键的动力。小麦复杂的基因组结构、外源的种植资源和染色体片断在改良中的应用,是目前面临着特殊的挑战。然而,随着小麦基因组测序工作的完成,分子育种成本逐渐降低,分子育种水平将得到快速发展。
[1] WENW,DENG QY,JIA H Y,et al.Sequence variations of the partially dominant DELLA gene Rht-B1c in wheat and their functional impacts[J].Journal of Experimental Botany,2013,11(64):3 299-3 312.
[2] FOLEY JA,RAMANKUTTY N,BRAUMAN KA,et al.Solutions for a cultivated planet[J].Nature,2011(478):337-342.
[3] GUPTA PK,LANGRIDGE PETER,MIR R R.Markerassisted wheat breeding:present status and future possibilities[J].Molecular Breeding,2010(26):145-161.
[4] JIN FENG,ZHANG D D,BOCKUS,et al.Fusarium head blight resistance in US winter wheat cultivars and elite breeding lines[J].Crop science,2012,53:2 006-2 013.
[5] JIA H Y,WAN H S,YANG SH,et al.Genetic dissection of yield-related traits in a recombinant inbred line population created using a key breeding parent in China’s wheat breeding[J].Theor.Appl.Genet.,2013,126:2 123-2 139.
[6] MA ZQ,WEIJB,CHENGSH.PCR-based markers for the powdery mildew resistance gene Pm 4a in wheat[J].Theor.Appl.Genet.,2004,109:140-145.
[7] LEONOVA I N.Molecular markers:Implementation in crop plant breeding for identification、introgression and gene pyramiding.R.J.G[J].Applied Research,2013,3(6):464-473.
[8] GUPTA P K,LANGRIDGE PETER,MIR R R.Marker-assisted wheat breeding:present status and future possibilities[J].Molecular Breeding,2010,26:145-161.
[9] FINKEL E RICHARD.making every drop count in the buildup to a blue evolution[J].Science,2009,323:1 004-1 005.
[10] KUCHEL H,FOX R,HOLLAMBY G,et al.The challenges of integrating new technologies into a wheat breeding programme[J].Molecular Breeding,2008,9:1-5.
[11] DUBCOVSKY J,MARKER.Aassisted selection in public breeding programs:the wheat experience[J].Crop Science,2004,44:1 895-1 898.
[12] BESPALOVA L A,VASSILYEV A V,ABLOVA I B,et al.The use of molecular markers in wheat breeding at the Lukyanenko Agricultural Research Institute.R.J.G[J].Applied Research,2012,2(4):286-290.
[13] MA H Q,KONG Z X,FU B S,et al.Identification and mapping of a new powdery mildew resistance gene on chromosome 6D of common wheat[J].Theor.Appl.Genet.,2011,7:1-8.
[14] TYAGI S,MIR R R,KAUR P H,et al.Marker-assisted pyramiding of eight QTLs/genes for seven different traits in common wheat (Triticumaestivum L.)[J].Molecular Breeding,2014,34(6):167-175.
[15] JIA J I,DEVOS K M,CHAO S,et al.RFLP based maps of the homoeologous group-6 chromosomes of wheat and their application in the tagging of Pm12,a powdery mildew resistance gene transferred from Aegilopsspeltoides to wheat[J].Theoretical and Appllied Genetics,2013,92:559-565.
[16] XUE SL,XU F,TANG M Z,et al.Precise mapping Fhb5,a major QTL conditioning resistance to Fusarium infection in bread wheat(Triticumae stivum L.)[J].Theor.Appl.Genet.,2011,123:1 055-1 063.
[17] SU X H,XU X W,JIANG G,et al.Fine mapping TaFLW1,a major QTL controlling flag leaf width in bread wheat(Triticumae stivum L.)[J].Theor.Appl.Genet.,2013,126:1 941-1 949.
[18] 马鸿翔,陆维忠.小麦赤霉病抗性改良研究进展[J].江苏农业学报,2010(1):197-203.
[19] 张海泉.粗山羊草Y189抗小麦白粉病基因SSR标记[J]. 河南大学学报,2007(37):177-180.
[20] 孔令让,董玉深,贾继增.小麦一粗山羊草双二倍体抗白粉病基因定位及其遗传转移[J].植物保护学报,1999,2(26):116-120.
[21] 欧巧明,崔文娟,王 炜,等.花粉管通道法导入高粱DNA创造优良小麦新品系的分子聚合育种[J].干旱地区农业研究,2013(2):2-12.
[22] COLLARD B C Y,JAHUFER M Z Z,BROUWER J B.An introduction to markers,quantitative trait loci(QTL)mapping and marker-assisted selection for crop improvement: The basic concepts[J]. Euphytica,2005,142:169-196.
猜你喜欢
贵州畜牧兽医(2023年3期)2023-06-29 07:07:28
农技服务(2023年2期)2023-03-15 00:43:08
今日农业(2021年11期)2021-08-13 08:53:24
世界科学技术-中医药现代化(2020年2期)2020-07-25 02:06:08
园林科技(2020年2期)2020-01-18 03:28:18
临床医药文献杂志(电子版)(2017年11期)2017-05-17 04:47:56
中学生物学(2016年8期)2016-01-18 09:08:21
作物研究(2014年6期)2014-03-01 03:39:03
遗传(2014年3期)2014-02-28 20:58:49
新疆农垦科技(2014年6期)2014-02-28 19:20:08
