
3种马尾藻周年生长和成熟时间以及形态学研究
2015-12-22 07:49:28曲元凯谢恩义孙立伟林巨华全建安陆志款
安徽农业科学 2015年23期
曲元凯,谢恩义*,孙立伟,林巨华,全建安,何 况,陆志款
(1.广东海洋大学水产学院,广东湛江524088;2.湛江市海洋与渔业发展研究中心,广东湛江524039)
马尾藻是广东省重要的经济藻类,可以作为鲍鱼、海参等重要经济动物的饵料,还可以提取褐藻胶等工业原料。由于沿海地区人们对马尾藻价值认识的提高,对马尾藻的利用和资源依赖程度逐步提高,又加上海区环境恶化、水质下降等因素,导致大量天然马尾藻床退化或消失。
半叶马尾藻中国变种(S.hemiphyllum var.chinense)、全缘马尾藻(S.integerrimum)、亨氏马尾藻(S.henslowianum)皆为马尾藻属中个体较大种类。半叶马尾藻中国变种,长度大约0.8~1.0 m,藻体黄褐色,在浪大的海区能够达到4 m以上;全缘马尾藻,长度可以达到2 m以上,黄褐色;亨氏马尾藻,黑褐色,在浪大的海区能够达到2 m以上。目前对这3种海藻的研究报道不多,已有文献主要集中于多糖提取以及保健方面的应用[1-7]。笔者通过野外调查和室内观察的方法研究了3种马尾藻周年的生长情况及其形态结构、成熟时间和繁殖盛期,旨在为3种马尾藻生物学、生态学、人工育苗以及藻场构建的研究提供基础资料。
1 材料与方法
1.1 研究区概况 硇洲岛位于广东省湛江市东南部,岛周围海底遍布岩礁,海水透明度高,十分利于海藻生长,马尾藻是该海区的优势种类。由于3种马尾藻分布潮带基本相同,生境相似,在硇洲岛东海头皆有分布,因此被选定为试验调查点(20°55'07.3″N,110°38'21.4″E),如图 1 所示。
1.2 仪器与耗材 采样铲(金属部分长3.5 cm、4 cm、6.5 cm的灰铲各1把);温度计(酒精温度计,温度测量范围-2~52℃,最小刻度为1℃);海水比重计(余姚市黄家埠玻璃仪表厂);卷尺(最小刻度为1 cm,最大测量距离15 m);游标卡尺(上海量具刃具厂,0~300 mm,最小刻度0.02 cm);架盘药物天平(上海医疗器械八厂,最大称量2 000 g,精确到1 g);尼康数码相机(800万象素);GPS仪;25 cm×25 cm自制采样框;标本袋;标签纸;塑料桶;采样封口袋等。
1.3 方法 于2012年6月至2013年5月平均每月1次(视潮水与台风情况而定)的调查都是在大潮期最低潮位时进行,采集3种马尾藻样品带回实验室进行外形观察、体长和生物量测量,并记录3种马尾藻生殖托形成及成熟的时间。
采用Origin7.5和SPSS18.0进行试验数据统计与分析,用One-way ANOVA检验差异的显著水平,P<0.05表示差异显著。
2 结果与分析
硇洲岛东海头马尾藻生长原生态的水温、海水比重周年各月变化如表1所示。3种马尾藻形态学指标如表2所示。
2.1 亨氏马尾藻的周年生长调查 从图2可以看出,亨氏马尾藻的生活周期可以划为休止期、生长期、成熟期和衰老期4个生活史阶段,5月底至10月为亨氏马尾藻的休止期。据调查,此阶段未能见亨氏马尾藻藻体,仅保留假根附着于低潮带岩石上,以此来度过夏季高温期;11月至次年3月初为生长加速期,此阶段亨氏马尾藻藻体生长迅速,次年1月底到3月初藻体可从原来的20 cm长至200 cm,2月底至3月初为整个生长期中生长最快的阶段,日平均生长速度可达5~10 cm;3月至4月初为亨氏马尾藻的成熟期,藻体因生殖托逐渐形成到成熟生物量增加较大(P<0.05),雄生殖托圆柱状,表面光滑,多单生,长1.5~5 cm,直径1 mm,雌生殖托纺锤形,单生,表面光滑,少数上部分叉,长约0.4~1 cm,直径1.0~1.5 mm;4月底至5月底为亨氏马尾藻的衰老期,藻体排卵后开始腐烂,叶先脱落腐烂,然后是顶端侧茎和主茎。6月,亨氏马尾藻藻体全部腐烂流失,仅剩假根保留在岩石上,此时海区水温较高,藻体进入新一轮的休止期。
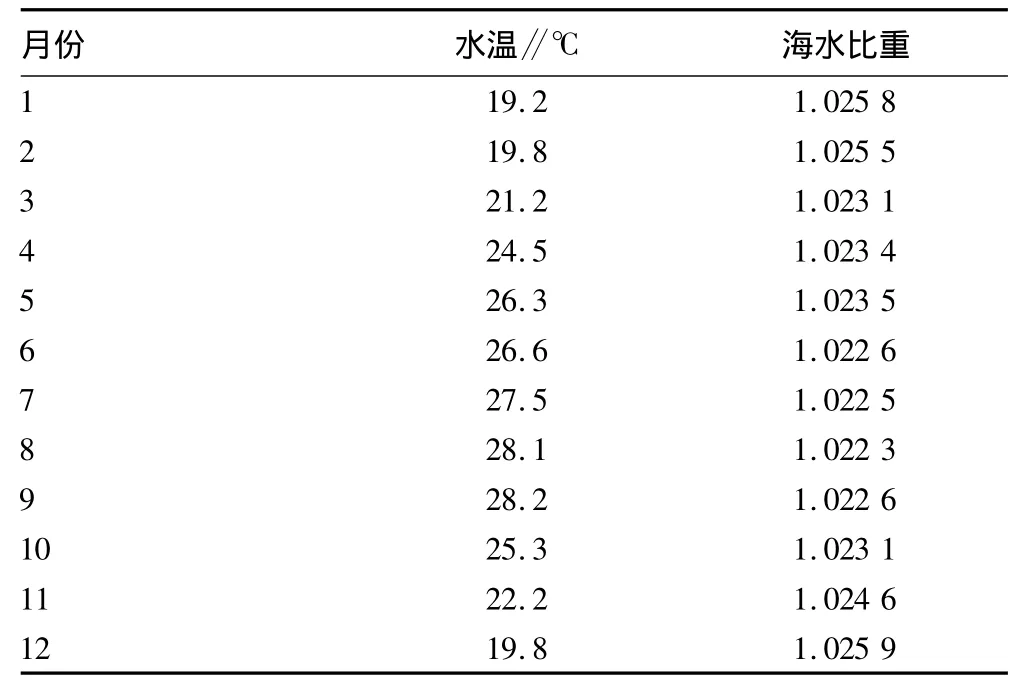
表1 硇洲岛东海头马尾藻生长原生态的周年各月水温和海水比重
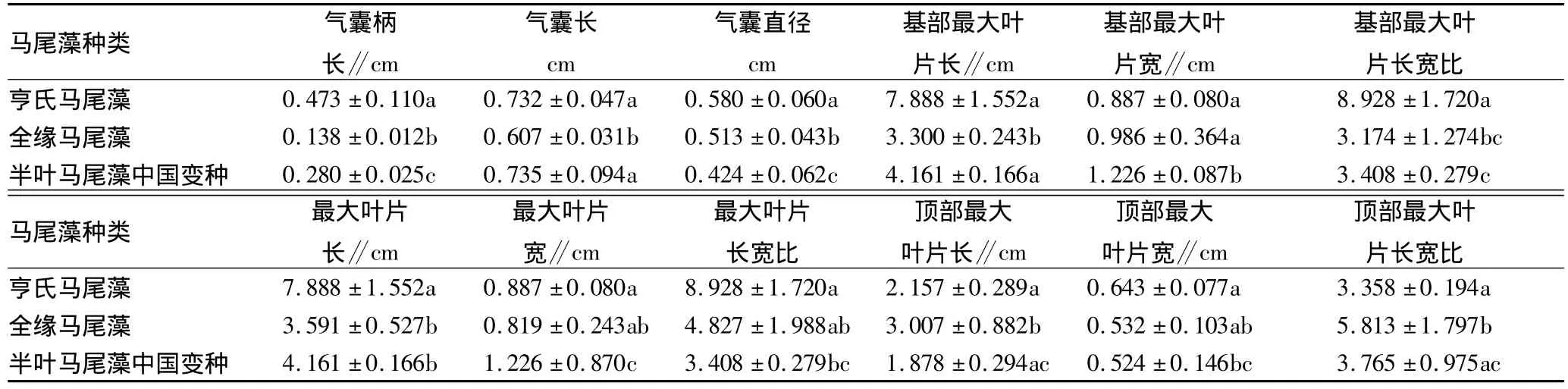
表2 3种马尾藻的形态学指标比较
亨氏马尾藻成熟期较早,3月底至4月初为繁殖盛期,据调查种群靠假根度过高温休止期,其幼孢子体容易遭受退潮干躁、日晒以及篮子鱼等敌害生物危害,因此野外亨氏马尾藻有性繁殖幼孢子体成活率极低,低于10%。绝大多数野生资源靠假根的无性繁殖来维持。有性繁殖的幼孢子体一般呈单株状,1个假根上生长1棵藻苗,而无性繁殖的幼孢子体一般为簇状,共同生长于一个假根上,假根较大,有性繁殖的藻体靠假根度过休止期后,第2年进行假根营养繁殖,假根开始增大,长出的藻体为簇状。
2.2 半叶马尾藻中国变种的周年生长调查 从图3可以看出,半叶马尾藻中国变种的生活周期同亨氏马尾藻也可以划分为休止期、生长期、成熟期和衰老期。6~10月为藻体休止期,水温高,藻体长度和生物量全年最低,仅保留假根来度夏。11月至次年4月上旬为生长期,此时海区水温降低,藻体度夏期结束,生长加速,长度和生物量增加迅速。4月中旬至5月中旬为成熟期,藻体开始出现生殖托,并逐渐成熟,繁殖盛期藻体长度和生物量最大,平均藻体长度为(140.0±26.6)cm。5月下旬至6月底为衰老期,藻体的叶片开始腐烂,紧接着藻体自顶端至基部的主侧枝也腐烂脱落,长度和生物量较低,最后藻体仅保留假根固着在岩石上度过夏天高温期,11月假根开始萌生幼孢子体进入新的生长期。
半叶马尾藻中国变种藻体是二倍体的孢子体,单倍体的配子体阶段仅在产生精子和卵细胞时存在,其繁殖可分为有性生殖和营养生殖2种方式。①有性生殖:孢子体的藻体产生精子囊和卵囊,囊内的母细胞通过减数分裂形成精子和卵,卵细胞由生殖托排出体外并粘附于雌性生殖托上,等待精子和卵细胞结合为受精卵,受精卵不休眠直接萌发之后形成新藻体。②营养生殖:二叉状分枝的假根度夏之后逐渐萌生出新的幼苗,幼苗生长迅速成为孢子体,假根直径较大约2~5 cm,假根发出的幼苗经常簇生,多株幼苗生长于一个假根上。
2.3 全缘马尾藻的周年生长调查 从图4可以看出,全缘马尾藻的生活周期同样分为休止期、生长期、成熟期和衰老期。全缘马尾藻休止期从7月中旬至10月底,此阶段为全缘马尾藻度夏阶段,调查期间未见该藻幼苗,仅假根附着在岩石上;11月至次年5月中旬为藻体的生长期,此阶段相对夏季水温较低,全缘马尾藻藻体生长迅速,从1月进入生长加速期,至5月中旬由原来的10~15 cm长至约200 cm,其中以3月中旬至5月初的生长速度最快,日生长速度最高可达10~15 cm;5月底到6月底为全缘马尾藻的成熟期,此期间藻体生长速度相对较慢,但藻体增重明显,生殖托较大,占藻体湿重的20%~30%;7月为全缘马尾藻的衰老期,藻体在成熟期结束后逐渐腐烂流失,至8月初仅剩假根保留在岩石上度过高温期。
野生全缘马尾藻资源主要靠营养繁殖维持,有性繁殖靠卵细胞受精发育而形成二倍体藻体,受精卵不休眠直接萌发之后形成新藻体,刚萌发的幼体抗逆能力弱,经受不了潮间带复杂的环境因子影响,因此有性繁殖的成活率极低。同样,全缘马尾藻营养繁殖的幼孢子体往往呈簇状生长,同一个假根上生出多个藻体,假根形状为假根状愈合,表面常可见假根凸起,中央略高,假根耐高温能力较强。
综上所述,3种马尾藻虽然繁殖时间存在差异,但其生活史都可分为休止期、生长期、成熟期和衰老期4个阶段。3种马尾藻都在繁殖盛期藻体长度和生物量达到最大。衰老期藻体逐渐腐烂流失,仅剩下假根。休止期未见小苗萌发,仅假根保留下来度夏。野生种群繁衍的维持以营养繁殖为主。
2.4 3种马尾藻的形态结构和成熟时间 根据对东海头试验点3种马尾藻的调查,3种马尾藻大多生长在岩石的迎浪面,背浪面较少。3种马尾藻中亨氏马尾藻成熟时间最早,在2月底、3月初形成生殖托,生殖托刚刚形成时比较短,雌雄难辩,至成熟时可达1.5~5.0 cm,生殖托成熟时由基部至顶端逐步成熟。雄性生殖托圆柱状,单生,成熟后生殖托顶端钝圆,雌生殖托成熟时顶端三分叉,分批排卵,雌雄生殖托表面光滑(图5A~B)。气囊通常为圆形,直径5~7 mm,顶端圆,顶端具有细尖,通常有圆柱状柄。初生分枝下部叶片较厚,披针形,顶端尖锐,中肋明显,到顶,长达6~8 cm,边缘具浅齿,毛窝分散在中肋的两边。上部藻叶呈窄披针形,与下部藻叶相似,但较窄,次生分枝上的藻叶呈窄披针形或线状,顶部尖,基部楔形,边缘具有锯齿。
半叶马尾藻中国变种生殖托于3月上旬(水温22℃左右)发生在叶腋间,丛生,一般为2~3个,多则5~6个(图5C~D)。生殖托发育成熟时,雌、雄区分明显:雌生殖托较粗,呈较短的圆柱形,前端钝圆,通常2~3个连在一起,表面凹凸不平,生殖窝所在的地方呈突出状态;雄生殖托呈较长的圆柱状,雄生殖托明显比雌托长而细。雌性生殖托在生殖窝中形成卵,雄性生殖托在生殖窝中形成精子囊产生精子。生殖托成熟时间为4月中旬,成熟雌生殖托2~3次分叉,长约4~7 mm,直径约0.7 mm。成熟雄生殖托圆柱形长达7~10 mm,直径0.6~0.8 mm,单条或2~3次分叉排成总状。气囊为倒卵形,顶端圆形,藻体上部的气囊常为纺锤形或椭圆形,顶端尖,边缘和顶端有翼状部分。藻体下部分的叶子左右不对称,一侧向外弧形弯曲,无中肋,基部叶片呈平行四边形,叶柄圆柱状,叶缘有粗齿,可长达5~7 cm。
全缘马尾藻成熟时间最晚,6月初,且同一地区生长的全缘马尾藻成熟并不同步。生殖托约在5月份形成,雌性生殖托为三棱形,长度为5~8 mm,顶端和边缘具有棘刺(图5E);雄性生殖托为圆柱状,顶端和边缘光滑。藻叶较厚,基部藻叶宽披针形,顶部略尖,中肋明显到顶,毛窝不规则的分散在中肋两侧,呈波状。次生分枝上的藻叶更细长,长披针形,线形,边缘全缘。
3 讨论
了解马尾藻各个时间段的生长情况及成熟时间对于构建人工藻场至关重要[8]。3种马尾藻虽然成熟时间不相同,但藻体成熟时排卵时间比较集中,这给人工育苗带来了很大的便利,影响马尾藻生长和繁殖的因子很多[9-13]。姜宏波等报道,浪大海区的海藻叶片平直呈流线型,分枝健壮,固着器较大,平静海区的海藻藻体长分枝浓密[14]。谢恩义等[9,15]研究表明风浪大水流畅通的海区有利于马尾藻的栽培和产量提高。王飞久等[16]研究发现鼠尾藻生殖托的成熟主要受到温度的控制。孙修涛等[17]研究表明鼠尾藻的成熟具有极大的不同步性,除温度外,光照和干露时间也是重要的影响因素。3种马尾藻中亨氏马尾藻繁殖时间最早,藻体的休止期也最长;全缘马尾藻繁殖时间最晚,藻体的休止期也最短。该研究结果表明,3种马尾藻的休止期都是于10月底结束,至11月便开始进入生长期,这可能与海区的海水温度有关,11月海区的海水温度约小于26℃,温度低于25℃时,有利于3种马尾藻幼苗的生长。3种马尾藻繁殖期过后马上开始进入衰老期,藻体逐渐腐烂流失,这可能是藻体基因调控决定的。3种马尾藻的体长在繁殖盛期时达到最长,生长速度在繁殖前一个月内较快,成熟藻体越长,其生长速度越快,这可能是藻体越大,接受光合作用的面积越大,藻体吸收的营养成分也越多。繁殖季节,藻体重量有较大增长,这是因为形成的生殖托重量占藻体鲜重比例较大。3种马尾藻的有性繁殖在生态学上属于R对策型,即将主要能量用于后代繁殖上,但对于迫害的规避能力较弱[18]。
整个休止期未见到3种马尾藻小苗萌发,仅假根保留下来度夏,营养繁殖以假根为主,因此3种马尾藻均为假根多年生海藻。由于多年生的假根固着能力强,假根生出的幼苗抗风浪能力也较强,在长出幼苗的同时假根也在不断生长,多年生的假根往往直径较大有些连成片状。生长出的幼苗往往簇生,更加强了幼苗的抗逆性。但是,近年来由于人类破坏,大量的马尾藻被连“根”拔起,导致资源量锐减。有性繁殖的幼孢子体第1年一般为单株状,一个假根上只生长一株苗,此类藻体抗逆性相对较弱,假根较小固着力相对较小,容易被风浪冲掉,至第2年无性繁殖时,假根也进一步生长,生出的苗也为簇生,与多年生的假根无异。因此,通过人工繁殖的方法构建海藻场,第1年藻体的存活尤为重要,第1年藻体存活后并保留假根度夏至第2年便可通过假根的营养繁殖生成新的幼苗,形成永久性的海藻场。
[1]傅明辉,佘纲哲,许素斌.亨氏马尾藻多糖的分离、纯化和鉴定[J].中国生化药物杂志,2000,21(2):66 -69.
[2]谌素华,王维民,刘辉,等.亨氏马尾藻化学成分分析及其营养学评价[J].食品研究与开发,2010,31(5):154 -156.
[3]卢睿春,侯振建.亨氏马尾藻硫酸多糖抗肿瘤活性的研究[J].海洋科学,1998,22(3):63 -64.
[4]谌素华,王维民,刘辉,等.亨氏马尾藻岩藻聚糖硫酸酯提取工艺的研究[J].食品工业科技,2011,32(8):269 -272.
[5]卢虹玉,杨小青,谢恩义,等.全缘马尾藻的主要营养成分分析与评价[J].食品研究与开发,2013(7):120 -122.
[6]孟庆勇,刘志辉,徐美奕,等.半叶马尾藻多糖的提取和分析[J].光谱学与光谱分析,2004,24(12):1560 -1562.
[7]刘秋英,孟庆勇.半叶马尾藻多糖体内抗肿瘤作用及其机制探讨[J].第一军医大学学报,2004,24(4):434 -436.
[8]章守宇,孙宏超.海藻场生态系统及其工程学研究进展[J].应用生态学报,2007,18(7):1647 -1653.
[9]谢恩义,贾柽,陈秀丽,等.硇洲马尾藻的繁殖特性及体长生物量的季节变动[J].水产学报,2011,35(7):1015 -1022.
[10]徐金根,陈素文,张汉华,等.温度和光照对瓦氏马尾藻繁殖的影响[J].广东农业科学,2013,40(9):121 -123.
[11]李伟新,朱仲嘉,刘凤贤.海藻学概论[M].上海:上海科学技术出版社,1982.
[12]YABG Y,FEI X,SONG J,et al.Growth of Gracilaria lemaneiformis under different cultivation conditions and its effects on nutrient removal in Chinese coastal waters[J].Aquaculture,2006,254(1/4):248 -255.
[13]WONG C P S.Biomass production of two Sargassum species at Cape Rachado,Malaysia[J].Hydrobiologia,2004,512(1):79 -88.
[14]姜宏波,田相利,董双林,等.温度和光照强度对鼠尾藻生长和生化组成的影响[J].应用生态学报,2009,20(1):185 -189.
[15]谢恩义,陈秀丽,申玉春,等.流沙湾7种海藻栽培比较及其对栽培海区水质的影响[J].热带海洋学报,2011,30(3):69 -75.
[16]王飞久,孙修涛,李锋.鼠尾藻的有性繁殖过程和幼苗培育技术研究[J].海洋水产研究,2006,27(5):1 -6.
[17]孙修涛,王飞久,王义民,等.基于有性繁殖的鼠尾藻规模化繁育试验[J].渔业科学进展,2010,31(3):84 -91.
[18]沈国英,黄凌风,郭丰,等.海洋生态学[M].北京:科学出版社,2010.
猜你喜欢
林业勘察设计(2021年4期)2021-04-21 08:58:58
大自然探索(2019年9期)2019-12-13 07:25:02
中国中医药现代远程教育(2019年23期)2019-01-31 04:13:12
儿童故事画报·自然探秘(2017年10期)2018-03-15 17:38:02
食品工业科技(2016年9期)2016-09-12 05:25:18
水产养殖(2016年4期)2016-08-25 08:39:16
广东海洋大学学报(2015年4期)2016-01-13 08:39:29
看世界(2014年24期)2014-12-23 18:15:38
科学养生(2014年11期)2014-11-18 10:45:53
海洋开发与管理(2012年11期)2012-11-07 05:46:14
