
西南地区木棉居群种子变异及萌发特性研究
2015-12-21 18:15王小玲伍建榕唐军荣马焕成
中南林业科技大学学报 2015年10期
高 柱,王小玲,伍建榕,唐军荣 ,马焕成
(1. 江西省科学院生物资源研究所 江西省重金属污染生态修复工程技术研究中心,江西 南昌 330096;2. 西南林业大学 国家林业局西南地区生物多样性保育重点实验室,云南 昆明 650224)
西南地区木棉居群种子变异及萌发特性研究
高 柱1,2,王小玲1,伍建榕2,唐军荣2,马焕成2
(1. 江西省科学院生物资源研究所 江西省重金属污染生态修复工程技术研究中心,江西 南昌 330096;2. 西南林业大学 国家林业局西南地区生物多样性保育重点实验室,云南 昆明 650224)
为明确木棉居群种子对环境长期适应产生的变异规律,对西南地区不同居群种子形态指标、种子萌发特性进行调查分析。结果表明:木棉不同居群种子变异系数为10.45%~28.85%,种子形态存在丰富的变异,平均种子长为4.44~7.30 mm,平均种子高为3.11~5.46 mm,勐堆乡居群种子最长,勐罕镇居群种子最高,物茂乡居群种子千粒重最重;东面种子较北方种子圆,年平均温度越高种子越高,西南方种子最重;木棉种子第4 d开始发芽,最高发芽势为49.25%,最高发芽率为57.05%,种子萌发变异系数为53.20%~93.32%,居群间种子萌发差异显著;木棉种子越大,萌发时间越长,但发芽率越高和发芽较整齐;根据种子形态指标可将西南地区木棉种子分为6个种群,种子形态未随居群种子地理位置连续或就近原则分类。可见,木棉局域性居群环境因子形成种子特有形态,影响种子萌发及幼苗生长,温度影响种子长、高及千粒重。
木棉;居群;种子性状;变异分析
丰富的生物种类和所拥有的基因,以及生存环境组成的生态系统构成了生物的多样性,对维持生态平衡、稳定环境以及保持持续发展具有重要作用。遗传多样性是生物多样性的核心和物质基础,林木变异主要有林木间的遗传变异、树木基因型与环境的相互作用,以及林木生长的环境变异[1]。种内遗传变异层次有地理种群变异,同一种源不同林分变异,同一林分不同个体变异及个体内变异[2]。植物形态特征受遗传基础、结构基因与调控基因、环境的综合作用,居群受地理阻隔和自然条件的长期作用,种子特征存在较大变异[3]。种子是被子植物更新生活史的重要阶段,也是植物向后代传递基因的重要途径[4],种子性状的变异是对复杂环境的一种适应[5]。
木棉Bombax ceibaL.别名英雄树、攀枝花、红棉等,为木棉科木棉属落叶大乔木,花大美丽,是南方重要行道树之一[6]。木棉适宜在光、热充足地区生长,是干热河谷植被恢复先锋树种之一,是西南地区重要的生态林、景观林、经济林树种[6-9];其果内棉毛为天然纤维原料,纤维具有中空率高、保暖性能好、抗菌等优点在纺织上备受青睐。木棉随海拔每增加80~150 m,花期推迟7~10 d,西南坡光照充足较东南坡光照弱的开花早[10];印度东部不同气候带的30个木棉种质选系的遗传分化测定显示,居群间存在丰富遗传变异[11];云南干热河谷地区木棉居群具有138个位点,PPB为89.86%,Nei’s基因多样性指数为0.255,居群遗传分化系数为0.117,同样具有丰富的遗传多样性,但居群间遗传分化低[12];吉贝果实性状指标变异系数为12.73%~25.26%,种子个数与果实长度存在显著相关性,种子个数还影响纤维产量[13]。通过对西南地区木棉种子变异及萌发特性的比较分析,掌握木棉种子的变异规律,可对木棉资源评价提供重要参考。
1 材料与方法
1.1 材料来源
试验收集材料来自西南地区各乡镇的地理单株/居群,共收集21个县、市,34个乡镇木棉居群种子,均为当年采集的种子,种源地概况、自然条件及编号见表1、表2。
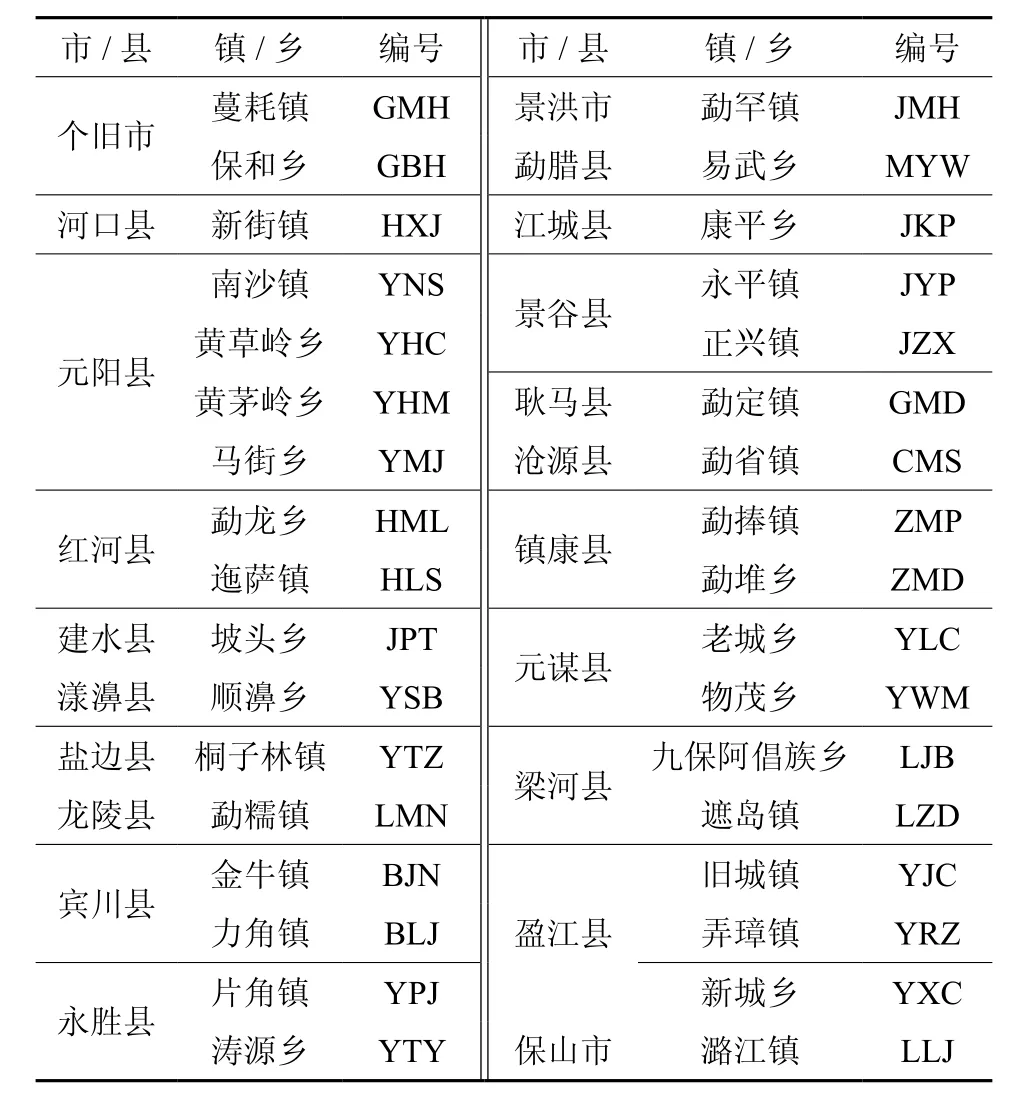
表1 木棉参试居群及编号Table 1 The tested population and No. of Kapok

表2 木棉居群地理、气候概况Table 2 Geography and climate overview of Kapok populations
1.2 试验方法
以红河、怒江、澜沧江、金沙江四流域为主线,每到一个地理居群,记录生态因子,目测选择生境条件水平相似、树龄相近的植株采种,每个居群10~12株,从不同方位选取正常成熟的果实,每株30个。
采用电子读书游标卡尺测定种子长、宽,用电子天平称量种子重、千粒重,不同产地种子形态采用产地均值与全部产地均值比较差异,采用欧氏距离聚类法确定种子形态分布区划。收集完种子后,为确定观测种子性状合适样本数,先将每个单株采集的种子取出混合均匀,然后在所有采集点中随机抽取10个点,每点100粒种子,分别测定种子长(胚根伸长方向)、高(垂直子叶方向)和籽重,计算不同样本数下各观测性状的变异系数,根据变异系数变化的曲线(图1)确定种子形态性状观测的样本数[14-15],木棉取30粒为种子形态观测样本数。

图1 木棉种子样本与性状变异系数关系Fig.1 Relationship between trait variance coefficient and number of Kapok seeds
每个居群取300粒种子,分成3组,为3次重复。播种前用清水浸泡24 h,然后用1%高锰酸钾溶液消毒10 min,冲洗后播种在装有表层红心土的营养袋中,常规管理。出苗后每天统计出苗数,记录开始发芽天数、发芽势(3 d)、发芽率(15 d),60 d后统计保存率、苗高及地茎。
2.1.3 统计分析
所有数据处理在Microsoft Excel 2007和SPSS 16.0 for Windows软件下完成,图在Sigmaplot 10.0软件下完成。采用系统
类聚分析相关性,LSD法分析差异显著性并进行Duncan多重比较,其中百分数经反正弦转换。
2 结果与分析
2.1 不同居群种子形态变异分析
将收集到的居群种子混合后,测定种子长(l)、高(h),千粒重,种子比重(籽重/长×高),结果见表3。从表3可以看出,木棉不同居群种子形态存在差异,平均长度为4.44~7.3 mm,平均种子高为3.11~5.46 mm,平均千粒重为20.49~50.99 g,种子重、长、宽比为1.05~3.11。其中,种子最长居群为ZMD(勐堆乡),较平均值长2.06 mm,是最短居群YXC(新城乡)种子长度的1.64倍,其次为居群JMH(勐罕镇)、YNS(南沙镇)、YWM(物茂乡)、JZX(正兴镇)、ZMP(勐捧镇);种子最高居群为勐罕镇,分别较平均值、最短居群JPT(坡头乡)种子高1.75 mm、2.35 mm,其次为居群勐堆乡、物茂乡、勐捧镇、GMD(勐定镇);千粒重最重居群为物茂乡,较平均值重14.39 g,是最轻居群YHM(黄茅岭乡)千粒重的2.49倍,其次为居群GMH(蔓耗镇)、LMN(勐糯镇)、BJN(金牛镇)。变异系数大小依次为种子比重、千粒重、种子高、种子长,其中种子比重和千粒重变异系数大于20%,说明木棉不同地理居群种子比重和千粒重存在较丰富的遗传多样性。LSD方差分析及多重比较显示,不同地理居群种子长、种子高、种子比重方差检验Sig.值为0.000,小于0.01,差异达极显著,千粒重差异显著(P<0.05)。
2.2 不同居群种子形态与生态因子相关分析
种子大小在同一物种内部被认为是相对稳定的,不同地理种群种子质量差异与种源所处地理位置、生境条件及气候特征有着重要相关性。木棉居群种子性状与环境生态因子相关系数见表4。由表4可知,种子长度与纬度、经度呈负相关,但未达显著水平,与海拔、年降雨量、年均温呈正相关,说明种子长度随纬度增加变短,越往东越矮,增加降雨和温度能够促进种子的增长;种子宽与纬度、年降雨量、年均温呈负相关,与经度、海拔呈正相关,相关性均未达显著水平,说明越往北种子越短,越往南种子越高,增加温度能够促进种子高生长;千粒重仅与年降雨量呈正相关,与其它生态因子呈负相关,说明西南地区木棉千粒重东北方低于西南方,温度越低越有利于种子增重;种子比重与千粒重相反,说明体积的增大较重量增加要快;种子形状往北越椭圆(与纬度呈正相关),往东越圆(与经度呈负相关)。综合相关系数认为,木棉不同居群种子形态与生态因子存在一定相关性,但相关性不显著,木棉种子的变异是由居群特有局域性气候综合因素决定。
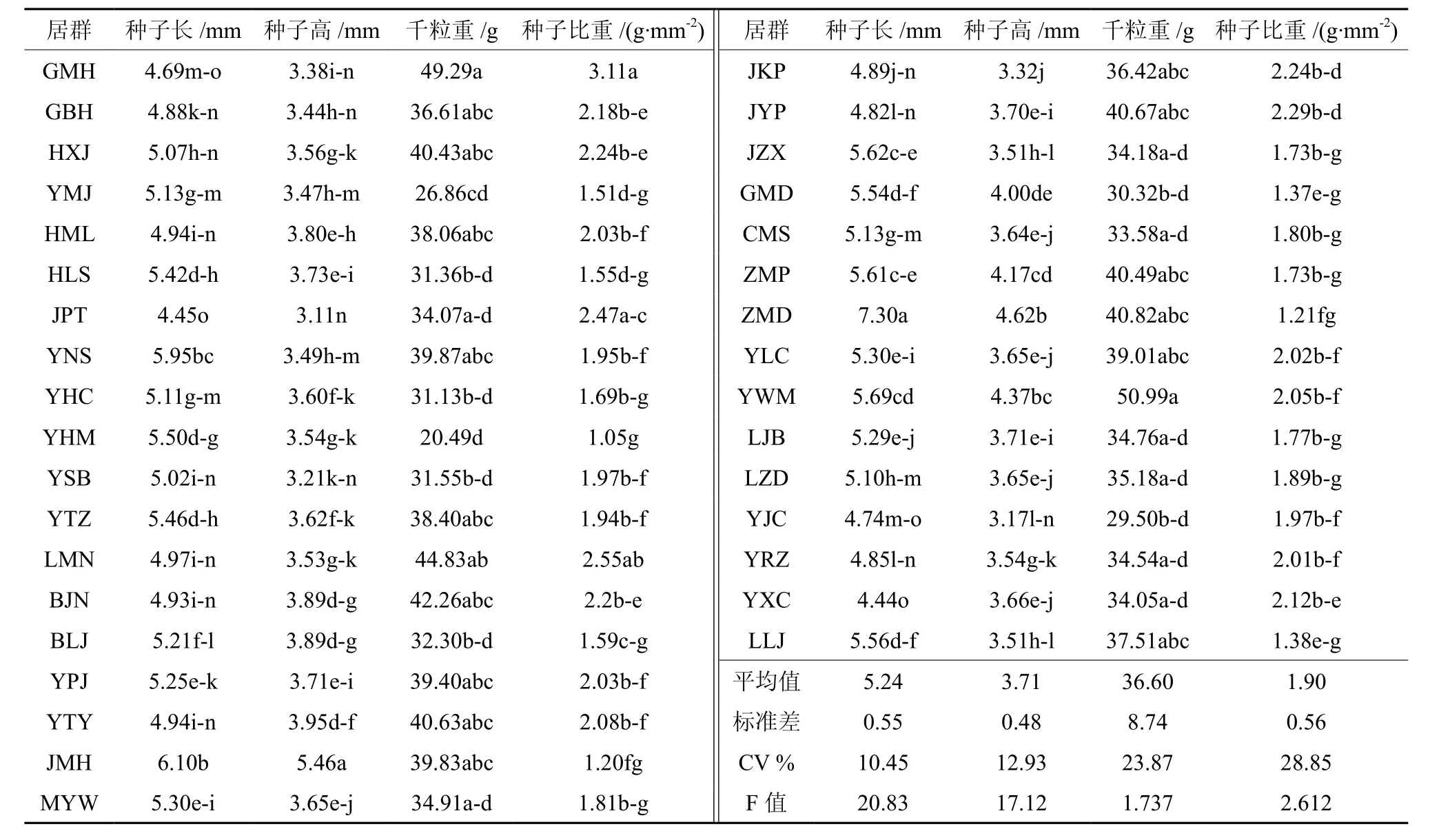
表3 木棉不同居群种子性状变异†Table 3 The variation of seed properties in different Kapok populations
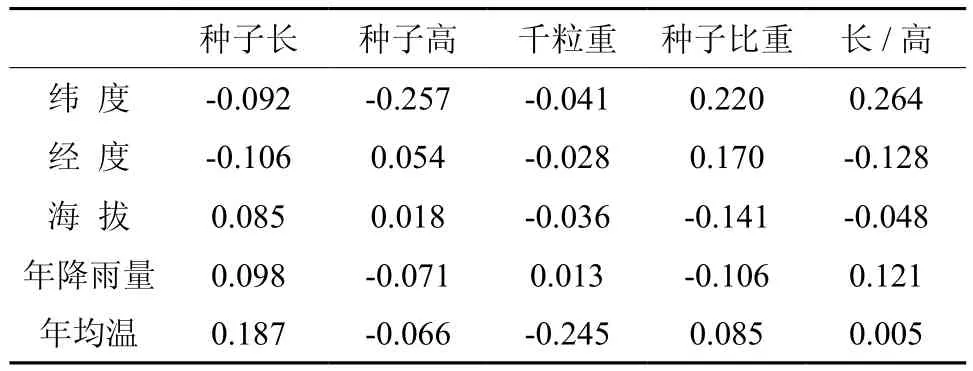
表4 木棉居群种子性状与地理、气候的相关性Table 4 Correlation analysis between seed properties and population geography, climate of Kapok populations
2.3 不同居群种子萌发特性
林木种子的出苗率及保存率在林业生产中直接关系到种子生长用量及苗木的质量。对收集的木棉居群种子,进行场圃播种发芽试验,自发芽开始每天统计萌发率,最终确定发芽后第3 d统计发芽势,第15 d统计发芽率,60 d统计保存率,结果见表5。从表5可以看出,除GMH(蔓耗镇)、YRZ(弄璋镇)居群种子没有发芽无法比较外,其它居群种子发芽性状均存在一定差异,种子开始发芽平均天数为4.00~9.00 d,平均发芽势为0.57%~49.25%,平均发芽率为0.29%~57.05%,平均保存率为12.24%~70.97%。发芽最早前5位为居群LJB(九保阿倡族乡)、GMD(勐定镇)、BLJ(力角镇)、BJN(金牛镇)、JMH(勐罕镇),较平均天数早1.97 d以上;居群JZX(正兴镇)、ZMD(勐堆乡)、JPT(坡头乡)、YHC(黄草岭乡)出苗最整齐,居群HML(勐龙乡)、HXJ(新街镇)、YTZ(桐子林镇)出苗最不整齐;最高发芽率为居群正兴镇,较平均发芽率高23.27%,较最低发芽率居群九保阿倡族乡高56.76%,其次为JYP(永平镇)、勐堆乡、GMD(勐定镇)、CMS(勐省镇)、YTY(涛源乡),最差的居群为九保阿倡族乡、GBH(保和乡)、新街镇、HML(勐龙乡)、YSB(顺濞乡)、YTZ(桐子林镇),平均发芽率低于10.00%,木棉居群种子发芽率均较低,应进行种子培育及播种时采用合理的种子催芽技术;苗保存率前5位依次为YMJ(马街乡)、保和乡、YXC(新城乡)、HLS(迤萨镇)、勐堆乡居群,最低为居群LZD(遮岛镇)、力角镇,不同居群保存率均较低,要加强种苗的管护。变异系数为53.20%~93.32%,发芽势变异系数最高,说明居群间种子萌发差异大,生产上要进行种子催芽处理以获得出苗的整齐度;其次为发芽率,说明居群间种子遗传差异显著。

表5 木棉不同居群种子萌发特性Table 5 The seed germination characteristics in different Kapok populations
LSD方差分析及Duncan比较显示,P值均小于0.000,表明4项统计指标内差异极显著,其中居群JYP(永平镇)、正兴镇与其它居群差异显著(P<0.05),居群马街乡保存率与其它居群差异显著(P<0.05)。由此说明,攀枝花居群种子萌发差异显著,萌发性好的居群可作为生产用种采集地。
2.4 不同居群种子形态与萌发特性及幼苗生长的相关性
林木种子形态对种子发芽及苗期生长存在显著相关性,但已有的报道认为树种不同,种子形态的影响也不同。采集到的木棉居群种子形态特征与种子发芽性状相关系数见表6。由表6可知,木棉种子长、高与发芽天数呈负相关,说明种子越大,萌发的时间越长;种子长、高与发芽率、发芽势呈正相关,说明种子越大,发芽率越高,发芽越整齐;种子比重与保存率呈正相关,与其它指标呈负相关,但未达显著水平。种子质量反映了居群种子营养物质的积累,相对饱满的种子营养物质充足,种子萌发快、发芽率高。
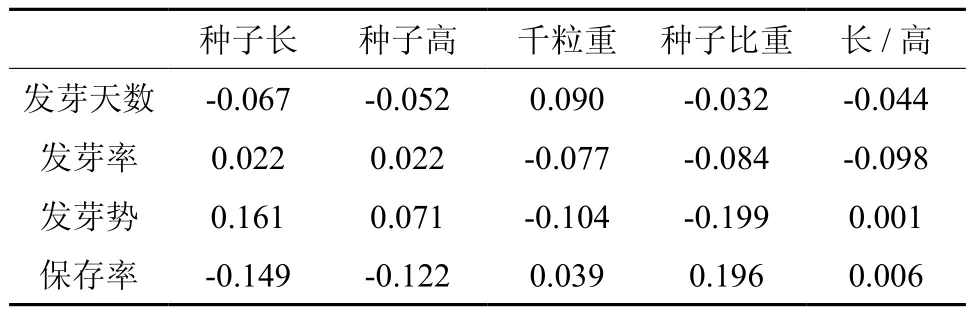
表6 木棉不同居群种子性状与萌发及苗期生长表型相关Table 6 The relationship between seed properties and its germination rate or seedling growth
2.5 种子特征分布区区划
根据种子长、高、长/高、长×高、千粒重、籽重/长×高6个表型特征,对西南地区木棉34个居群进行欧式距离聚类分析(图2)。从图2中可以看出,34个居群按种子性状指标可明显划分为8种群(特征值见表7),第1种群有ZMD(勐堆乡)、JMH(勐罕镇)2个居群,表现为种子长、高最大,长/高比最小,长×高最大,说明该种群种子较圆、较大;第2种群有YWM(物茂乡)、LMN(勐糯镇)、GMH(蔓耗镇)3个居群,表现为千粒重最大,说明该种群种子最重;第3种群有JPT(坡头乡)、YNS(南沙镇)2个居群;第4种群有10个居群;第5种群有YSB(顺濞乡)、YJC(老城乡)、YMJ(马街乡)3个居群,表现为长/宽最大,说明该种群种子呈长扁型;第6、7、8种群分别含有4、3、7个居群。同一种群内各居群种子表型特征基本相近,而种群间变异较大(表5)。木棉种子表型特征并未随居群地理位置的连续或最近原则分类,这与云南独特小区域气候有关,同时也说明木棉种子表型特征是居群生态因子综合作用影响的结果。

图2 木棉居群种子产地欧氏聚类分析Fig.2 The cluster analysis for quality traits of Kapok species

表7 各区组木棉种子表型特征Table 7 Seed phenotypes in different district groups of Kapok
3 结论与讨论
3.1 木棉种子形态与环境因子相关性
不同树种遗传特性、地理环境气候及生长条件的变化造成了种子性状趋变模式的差异,受地理阻隔和自然条件长期作用,种子在居群间可以发生较大的差异[3]。种子形态在同一植物种内或品种内认为是相对稳定的,而在种群内、种群间或者个体间,会存在一定的差异。另外,种子作为果实重要组成之一,其发育与果实生长存在密切关系,山葡萄Vitis amurensis含有种子的果实较没有种子的果实大,种子生长发育的同时可能产生内源激素促进浆果膨大[16];菠萝蜜Artocarpus heterophyllus果皮包刺密度与果肉可溶性固形物含量密切相关,可通过外形选择优质品种[17];芒果Mangifera indica可食率随果实增大而提高,而小果型可溶性固形物含量高[18]。因此,果实和种子形态指标不仅可以作为种源差异评判标准,还可作为不同物种选育指标实现选育目的。
木棉种子形态变异系数为10.45%~28.85%,不同居群种子存在丰富的变异,该结论与Chaturvedi等[11]、汪书丽[12]采用分子技术测定居群遗传变异结果相似。种子长度与纬度、经度呈负相关,与海拔、年降雨量、年均温呈正相关,该变异与厚朴种子随经纬变化具有相似性[19];千粒重与年降雨量呈正相关,降雨量增加和温度升高能够促进种子的增长,北面种子较南面圆,西面较东面圆,东北方种子较西南方轻。综合分析认为,木棉不同地理居群种子形态存在丰富的变异,种子性状与气候因子存在一定的关系,西南地区特殊的局域性气候影响了种子变化趋势的连续性,种子的变异是地理分布区局域性气候综合影响的结果,体现变异的空间趋势。
3.2 木棉种子形态对种子萌发及幼苗生长的影响
种子大小和重量影响种子萌发速度和发芽率,进而影响苗木早期生长[20]。林木种子出苗率在林业生产中直接关系到生产用量及苗木的质量,而种子形态特征与种子发芽及苗期生长的影响,已有的报道存在差异。樟树不同产地种子场圃出苗率存在极显著差异,种子形态指数对出苗率有影响,但不显著,而种子长/宽比与苗木生长密切相关[15];乌药种子形态指数对出苗率有不显著的影响,但种子宽与一年生苗高呈极显著正相关[21]。小桐子种子千粒重越大,出苗率越高,而种子形态及千粒重与苗高、地茎生长特性无关。栎属种子性状对苗期生长无显著关系,种子重生长反而差[22]。本试验场圃发芽试验显示,木棉不同居群种子发芽性状指标表型变异系数为53.20%~93.32%,不同居群种子发芽性状差异显著。开始发芽天数为4.00~9.00 d,出苗最快为发芽后3 d,正兴镇居群发芽最整齐,发芽率最高。种子越大,发芽率越高,出苗越整齐;种子越长,发芽后苗高生长也越快;千粒重对攀枝花种子发芽率影响不显著;种子比重越大,苗期保存率也越高。综合认为,种子性状对木棉发芽率及苗期生长存在影响,但影响较小。
3.3 木棉种子形态及其种群区划特性
聚类分析可将相似性状的种源或种群进行分组,相同分组具有相似或相近的性状。木棉不同居群按种子性状可分为8个种群,第1种群种子最圆、大,第2种群种子最重,第3~8种群各项指标均处于中间水平。木棉种子表型特征并未随地理位置的连续或就近原则分类,这与不同地理居群小区域气候差异有关,同时也说明木棉种子变异是地理生态因子综合影响的结果。
[1]毛爱华. 河南郏县侧柏种源、家系及无性系遗传变异与选择[D]. 北京: 北京林业大学, 2009.
[2]王秋玉. 红皮云杉地理种源的遗传变异[D]. 哈尔滨: 东北林业大学, 2003.
[3]Greipsson S, Davy A J. Seed mass and germination behaviour in populations of the dune building grass Leym usarenarius[J].Annals of Botany, 1995, 76: 493-501.
[4]张大勇. 植物生活史进化与繁殖生态学[M]. 北京: 科学出版社, 2004.
[5]舒 枭, 杨志玲, 杨 旭, 等. 不同产地厚朴种子性状的变异分析[J]. 林业科学研究, 2010, 23(3): 457-461.
[6]高 柱, 王小玲, 马焕成, 等. 木棉栽培技术研究进展[J]. 江西科学 , 2009, 27(5): 761-766.
[7]李文华, 熊定国. 木棉产业导引: 首届中国木棉产业发展研讨会文集[C]. 中国纺织出版社, 2009.
[8]高 柱, 伍建榕, 马焕成. 红河干热河谷2年生吉贝人工幼林施肥效应研究[J]. 水土保持研究, 2012, 19(2): 95-100.
[9]高 柱, 伍建榕, 严 毅, 等. 3种配方处理吉贝插穗生根过氧化物酶(POD)活性变化的研究[J]. 东北农业大学学报,2012, 43(10): 151-155.
[10]胡 亮, 陈邦美, 何和明. 海南西南地区尖峰岭木棉开花生物学特性[J]. 蜜蜂杂志 , 2006, 2(3): 39-40.
[11]Chaturvedi O P, Pandey N. Genetic Divergence inBombax ceibaL. Germplasms[J]. Siline Genetion, 2001, 50(3-4): 99-102.
[12]汪书丽. 云南干热河谷地区木棉群居的遗传多样性研究[D].西双版纳: 中国科学院, 2007.
[13]高 柱, 田向楠, 郑 元, 等. 云南吉贝种质资源调查及果实性状相关性分析[J]. 经济林研究, 2012, 30(1): 88-94.
[14]George W. Snedecor, William G. Statistical Mehtods[M]. The Iowa State University Press, Ames Iowa. USA, 1980: 215-237.
[15]姚小华. 樟树遗传变异与选择的研究[D]. 长沙: 中南林学院,2002.
[16]葛玉香, 沈育杰, 李晓红. 山葡萄种子性状与果实品质的关系[J]. 特产研究 , 1999, (2): 36-39.
[17]叶春海, 吴 钿, 丰 锋, 等. 菠萝蜜种质资源调查及果实性状的相关分析[J]. 热带作物学报, 2006, 27(1): 28-32.
[18]李国鹏, 张林辉, 解德宏, 等. 怒江干热河谷芒果种质资源果实性状相关性及多样性分析[J]. 西南农业学报, 2010, 23(3):1225-1229.
[19]杨志玲, 杨 旭, 谭梓峰, 等. 厚朴不同种源及家系种子性状的变异[J]. 中南林业科技大学学报, 2009, 29(5): 49-55.
[20]姜景民, 虞沫奎, 胡世才, 等. 湿地松种子分级育苗的研究[J]. 林业科学研究 , 1996, 9(3): 290-295.
[21]陈丽华, 姜景民, 栾启福, 等. 乌药种子性状产地表型变异研究[J]. 浙江林业科技, 2005, 25(1): 9-11.
[22]汪企明, 李晓储, 黄利斌, 等. 美国栎属种源引种、变异研究:种子及苗期生长变异[J]. 江苏林业科技, 1998, 26(1): 1-6.
Study on Seed Variation and Germination Characteristics of Kapok Populations in Southwest of China
GAO Zhu1,2, WANG Xiao-ling1, WU Jian-rong2, TANG Jun-rong2, MA Huan-cheng2
(1. Jiangxi Engineering Research Center For Eco-Remedication of Heavy Metal Pollution /Institute of Biological Resources, Jiangxi Academy of Sciences, Nanchang 330096, Jiangxi, China; 2. Key Laboratory of Biodiversity Conservation in Southwest China, State Forestry Administration, Southwest Forestry University, Kunming 650224, Yunnan, China)
In order to reveal the variation law generated long-term adaptation to the environment on different populations Kapok, seed morphology and germination characteristics of different population Kapok in the southwest of china were investigated and analyzed.The results showed that the seeds in different populations coefficient of variation was 10.45% to 28.85%, there were abundant variations between different populations of Kapok. The average seed length was 4.44 mm to 7.30 mm, and the average height of seed was 3.11 mm to 5.46 mm. The longest seed was population of Mengdui town, the highest seed was population of Menghan town, and the grain weight was population of Wumao town. It was rounder the seed in east than in north. The seed live in the higher annual average temperature was taller, however the heaviest seed was in southwest. The seed was began to germinate in the 4th day, and the highest germination potential and germination rate were 49.25% and 57.05%. Seed germination coefficient of variation was 53.20% to 93.32%, it has significantly different among population of Kapok. The larger seed the germination times was longer, but the germination rate was higher and the germination was more neat. According to the seed morphological indicators can be divided into six populations of Kapok in southwest of china, but seed morphology classification principles were not continuous or nearest location with different populations. As a result,Seed-specific forms, seed germination and seedling growth were affected by environment factors which Kapok population was localized,temperatures was affected seed length, seed height and grain weight.
Kapok; populations; seed characters; analysis of variance
S722.3+1
A
1673-923X(2015)10-0006-07
10.14067/j.cnki.1673-923x.2015.10.002
2014-02-25
国家自然基金项目(31260175);江西省科学院科研开发专项基金(2012-YQC-06);江西省科学院产学研合作资金项目(2013-09)
高 柱,硕士,副研究员
马焕成,教授;E-mail:mahuancheng@yahoo.com.cn
高 柱,王小玲,伍建榕,等. 西南地区木棉居群种子变异及萌发特性研究[J].中南林业科技大学学报,2015,35(10):6-12.
[本文编校:吴 彬]
猜你喜欢
植物研究(2022年5期)2022-10-12
河北科技师范学院学报(2022年2期)2022-08-26
浙江中医药大学学报(2021年6期)2021-07-12
歌海(2021年6期)2021-02-01
中国粮油学报(2020年12期)2021-01-09
幼儿园(2020年15期)2020-11-24
艺术家(2020年5期)2020-07-09
海峡姐妹(2019年5期)2019-06-18
植物研究(2019年2期)2019-03-19
种子科技(2018年11期)2018-09-10
