
菹草石芽萌发及其生长过程的初步探究
2015-12-20 08:02尹传宝张翠英2申云飞王龙刘自强张敏沈应时
湿地科学与管理 2015年1期
尹传宝张翠英,2申云飞王龙刘自强张敏沈应时
(1 徐州工程学院环境工程学院,江苏 徐州 221111;2 中国矿业大学环境与测绘学院,江苏 徐州221116)
菹草石芽萌发及其生长过程的初步探究
尹传宝1张翠英1,2申云飞1王龙1刘自强1张敏1沈应时1
(1 徐州工程学院环境工程学院,江苏 徐州 221111;2 中国矿业大学环境与测绘学院,江苏 徐州221116)
菹草Potamogeton crispus为眼子菜科多年生沉水草本植物,分布广泛,是我国很多湖沼、池塘水体的优势种,菹草可作草食鱼类及家禽、家畜的饲料,具有较高的营养价值和经济价值(简永兴等,2001)。在自然环境中菹草主要依靠植株顶端的繁殖鳞茎——石芽(又叫休眠芽、芽苞、殖芽等)进行繁殖。石芽坚硬边缘具有锯齿,形如松果,待水温适宜即开始萌发生长(Barko et al,1981)。
目前有关沉水植物繁殖体研究的报道很多,国外已在繁殖体的萌发生态方面作了大量工作(Baskin et al,1996; Lombard i et al,1997; Brenchley et al,1998;Balestri et al,1998),国内在以资源利用为目的的沉水植物种植方面、繁殖体的萌发生态及其影响因子方面已进行了一些研究(连光华等,1996;黄运平等,2001;崔心红等,2000),而对菹草石芽萌发机理的研究报道相对较少。菹草兼有无性繁殖和有性繁殖两种繁殖方式(Rogers et al,1980;任久长等,1997),但菹草种子在自然状况下极少萌发,主要靠特化的石芽进行繁殖(陈小峰等,2006),而石芽是在强光、长日照、水温高的季节产生的(陈小峰等,2006)。由于水体受到污染,水体透明度降低,菹草在萌发期和苗期得不到适宜的光照,影响其正常的光合作用,甚至生长发育受阻。由于近年来水体富营养化日趋严重,各地均有菹草爆发的事例。研究菹草石芽的萌发和幼苗生长机理能够为人工湿地及自然水体生态修复提供预测和控制菹草爆发的理论依据,为渔业和人工池塘菹草的栽培、育种以及健康生态系统的建立和稳定提供数据支持。
1 材料与方法
1.1 实验材料
实验所用石芽皆为当年芽,石芽大小均匀,均重0.7 g,采自徐州工程学院校园景观湖。景观湖水体营养水平高,夏季生长有菹草、金鱼藻,狐尾藻等水生植物。2013年9月6日实测水体各项指标见表1。
1.2 实验设计
1.2.1 光照对石芽萌发的影响 一般认为菹草石芽秋后萌发,本实验用培养箱模拟秋冬季水温条件,设置白天温度15 ℃,夜间10 ℃,白天和黑夜各12 h,白天光照强度6级,夜晚无光照。白天设置不同光照条件,以玻璃杯外加玻璃隔板层数实现光照强度的梯度,分别设置光照强度梯度为2 000 l x、1 400 l x、900 l x、500 l x、0 l x。共5个光照梯度,
实验在9月2日开始,在室内光照培养箱进行实验,共选取新采集石芽300颗,石芽放在玻璃杯(直径12 cm,高12 cm)内,每个玻璃杯内放置石芽30颗,根据光照梯度实验分5组,每组两个平行共用10个玻璃杯,无土栽培,向杯内加水至2/3杯高,实验期间适当补充蒸发损耗的水,实验用水为景观湖水。
1.2.2 温度对石芽萌发的影响 本实验共分A组、B组、低温组和常温组4组,其中A组、B组和低温组与上组实验同时于9月2日在培养箱内进行,温度和光照条件相同。各组中每个玻璃杯放置石芽30颗,用玻璃隔板设置光照强度1 400 lx。培养箱内温度控制白天15℃,夜间10℃,白天光照强度6级,夜间光照为0。9月5日取出A组、B组放置在室内正常温度下(9月初温度26~30℃),使光照强度为1 400 lx,观察萌发情况。低温组培养箱内继续观察。常温组同时于9月2日在室内正常温度26~30℃下开始,白天光照强度1 400 lx,夜晚为0。
1.2.3 石芽在自然状态下的生长机理 选取大小均匀石芽60颗,称重后,根据景观湖形状,沿湖布置6个实验点,每点放置10颗石芽。每颗石芽均用尼龙细线系住置于湖水中,在细线上做好每个石芽的标记,细线足够长,石芽分布散开避免细线缠绕,线头拴在岸边以便观察。
1.3 实验方法
以石芽萌发出的芽苗长于1 cm时为标准,即认定已萌发。对每组实验每天观察、统计萌发情况,直至各组石芽萌发率趋于稳定,实验终止。第3组实验菹草石芽全部下沉后取回实验室称重。
2 结果与分析
2.1 光照对石芽萌发的影响
光照对石芽萌发率统计如图1所示,石芽在有光照和无光照条件下均能萌发,从图1的萌发率曲线来看,从9月2日至9月17日,石芽基本全部萌发,前4组萌发率均为100%,第5组为95%。各组萌发情况近似,与光照梯度无明显相关性,即在较低的温度条件下弱光环境对菹草石芽的萌发影响不大。

图1 不同光照强度条件下石芽萌发率

图2 低温和常温下石芽萌发率

表1 实验水体各项指标含量
2.2 温度对石芽萌发的影响
1)石芽在低温条件和常温条件下的萌发率。从图2可看出,9月初正常温度下菹草石芽在8-10 d 萌发率就可达到100%,而在温度低的情况之下大约需14 d 的时间才可达到,低温条件下菹草石芽的萌发速度低于较高温度条件的实验组。
2)温度交叉条件下石芽的萌发情况。根据各组实验的对比发现,9月2日至9月5日A组、B组和低温组的萌发率均低于常温组,低温条件下萌发速
度慢、萌发率较低;而在回到日常温度后9月5日至9月7日,A组和B组萌发迅速上升至93%和97%,9月8日A组97%,B组达到100%,至完全萌发比常温组提前1-2天,对比前一组实验完全萌发比低温组提前约7天(图3)。这说明石芽在一段时间的低温培养后,恢复常温使石芽的萌发率迅速升高,萌发时间缩短。

图3 温度交叉条件下石芽萌发率
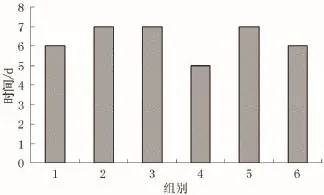
图4 自然状态下石芽吸附悬浮物下沉的时间

图5 石芽在自然状态下吸附悬浮物情况
2.3 自然状态下石芽的生长过程
1)根据实验观察发现,菹草石芽密度小于水,漂浮于水面,而由于湖面波动,石芽在随水面波动中会吸附湖水中的悬浮颗粒,使自身重量增加沉入水底,并生根于底泥中。本次实验所选取湖水悬浮物含量为86 mg/L,实验发现石芽5-7 d (图4)就能吸附自身重量一倍以上的悬浮物颗粒(图5),使自身重量增加并沉入水底。
2)一般在7-8月菹草植株衰退腐烂后即可形成许多独立的冬芽,冬芽密度小于水,浮于水面,但冬芽特殊的结构(鳞状,边缘有齿)使它易于粘附水体中的悬浮泥沙,大量的悬浮泥沙被吸附后,冬芽重量增加并沉入水底,待环境合适生根扎于底泥中。底泥中含有丰富的营养物质,能保证冬芽顺利萌发生长。菹草的生长过程如图6所示。
3)9-10月我国北方大部分地区秋高气爽,湖水温度条件较好,实验调查发现,此时菹草石芽已开始萌发(图7),但随着天气逐渐转凉,萌发后的菹草石芽并不会大规模生长,这时的菹草一般叶片稀少,但茎部纤细且长,纤细的茎不能支撑菹草直立生长,倒伏下的茎叶触到河流基质时就会生根(图8),形成新的植株。菹草的无性生殖是导致后期菹草大规模繁殖,且草群密集分布的主要原因。

图6 菹草的生长过程

图7 菹草石芽萌发图

图8 菹草茎部的无性增殖
3 结论与讨论
1)普遍认为石芽多数在秋季萌发,萌发温度15~25 ℃,温度越低越有利于石芽的萌发( Rogers et al,1980)。本实验模拟秋末冬初温度条件,研究光照和温度对石芽萌发的影响。结果证实石芽在该温度下可以萌发并且萌发情况较好,根据图1萌发率曲线,在较低的温度条件下,光照对菹草石芽的萌发无显著影响,这一结论与杨文斌等(2009)、李永函等(1989)的研究结果一致。
2)李强等(2009)研究认为,秋冬季节是菹草石芽萌发的主要季节,自然条件下9-11月石芽大量萌发,幼苗生长发育快,有利于其在翌年春季形成大量的水生植被。本实验用的菹草石芽于9月初采集于校园人工湖,9月初个别石芽已出现萌发现象,且实验石芽在14-17 d 内萌发率就可达到100%。100%的萌发率显著高于野外调查获得的值(简永兴等,2001),这可能与实验所用的石芽个体较大、发育好、差异小有关。
3)低温和高温的交叉实验证实,在一段时间的低温条件下,恢复较高的温度会提高石芽的萌发速率,缩短正常萌发所用的时间。这也很好地验证了越冬后的菹草适应性强,生长旺盛。菹草的这一特性和通过自身茎部的无性拓殖能力可能是造成后期菹草大规模繁殖,且草群密集分布的主要原因。
4)调查发现,菹草石芽密度小于水,正常状态下漂浮在水面上,而石芽是沉水植物菹草的繁殖方式。本实验证实由于湖面水体的波动,菹草石芽能在短时间里吸附大量悬浮在水体中的颗粒物,并使自身重量增加并沉入水下。石芽的锯齿边缘、松果形状结构也间接证明了它有足够的空间附着悬浮物,并能粘附底泥。实验证明一般石芽吸附自身重量一倍左右的悬浮物质,便能够使自身下沉至湖泊底部,石芽吸附的悬浮物及底泥均含有一定的营养成分,能促进石芽的萌发。
陈小峰,陈开宁,肖月娥,等. 2006.光和基质对菹草石芽萌发、幼苗生长及叶片光合效率的影响[J].应用生态学报,17(8):1413-1418
崔心红,熊秉红,蒲云海,等. 2000. 5种沉水植物无性繁殖和定居能力的比较研究[J].植物生态学报,24(4):502-505
黄运平,蒲云海. 2001.竹叶眼子菜冬芽萌发的生态研究[J].中南民族学院学报:自然科学版,20(3):85-88
简永兴,王建波,何国庆,等. 2001.水深、基质、光和去苗对菹草石芽萌发的影响[J] 水生生物学报,25(3):224-229
连光华,张圣照. 1996.伊乐藻等水生高等植物的快速营养繁殖技术和栽培方法[J].湖泊科学,8(增刊):11-16
李强,王国祥.2009.秋冬季光照、水温对菹草萌发和幼苗生长发育的影响[J].重庆文理学院报:自然科学版,28(1):9-15
李永函,金送笛,史进禄,等. 1989.几种生态因子对菹草鳞枝形成和萌发的影响[J].大连水产学院学报,4(3):1-9
任久长,乔建荣,董巍,等.1 9 9 7.菹草(Potamogeton crispus)的生态习性和在京密引水渠的发生规律研究)[J].北京大学学报:自然科学版,33(6):749-754
杨文斌,王国祥,郑海洋,等. 2009.菹草石芽萌发及幼苗生长对光、温因子的响应[J].生态与农村环境学报,25(1):96-100,105
Balestri E, Piazzi L, Cinelli F.1998. In vitro Germination and Seed ing D evelopment of Posid onia Oceanica [J].Aquatic Botany, 60:83-93
Barko J M, Smart R M. 1981.Comparative influences of light and temperature on the growth and metabolism of selected submerged freshwater macrophytes [ J ]. Ecology Monograph, 51: 219 -235
Baskin C C, Baskin J M, Chester E W. 1996. Seed Germina-tion Ecology of the Aquatic Winter Annual Hottonia Inflata[J].Aquatic Botany, 54:51-57
Brenchley J L, Probert R J. 1998. Seed Germination Re-sponse to Some Environmental Factors in the Seagrass Zostera Capricorni from Eastern Austalia[J]. Aquatic Botany,62:177-188
Rogers K H, Breen C M.1980. Growth and reprod uction of Potamogeton crispus in a South African lake[J]. Journal of Ecology,68:561-571
Lombard i T, Fochetti T, Bertacchi A.1997. Germination Requirements in a Population of Typha Latifolia [J].Aquatic Botany,56:1-10
A Preliminary Study of Germination and Growth Process of the Turion of Potamogeton crispus
YIN Chuan-Bao1ZHANG Cui-Ying1,2SHEN Yun-Fei1WANG Long1LIU Zi-Qiang1ZHANG Min1SHEN Ying-Shi1
(1 School of Environment Engineering, Xuzhou Institute of Technology, Xuzhou 221111; 2 School of Environment Science and Spatial Informatics, China University of Mining and Technology, Xuzhou 221116)
研究了光照和温度对菹草石芽萌发的影响及菹草的生长机理。结果表明:①在模拟秋冬季温度条件下不同光照强度对石芽萌发率的影响无规律性,即光照并未对菹草石芽的萌发起促进作用;②石芽在一定时间的低温作用后,恢复常温使石芽的萌发速度和萌发率明显提高,说明石芽的萌发受温度的影响,这也更好地解释了菹草的越冬生长习性及春季部分水体菹草大规模爆发的现象;③石芽随水面波动可吸附大量悬浮物,一般5-7天即可增加足够的重量下沉至水体底部,石芽吸附的悬浮物含有一定的营养成分,能促进石芽的萌发。石芽的萌发机制可为菹草石芽繁殖的影响因素研究提供依据,对栽培菹草具有重要的实践意义。
菹草;石芽;萌发率;悬浮物
Influence of light and temperature on turion germination of Potamogeton crispus and the growth mechanism of P. crispus were studied. Results showed that: 1) In simulated of autumn and winter temperature conditions the effects of different light intensities on germination rate of turions of P. crispus displayed no regularity, indicating that light did not play a catalytic role in turion germination 2) Treated with low temperature for a certain time, germination speed and germination rate were evidently increased by restoring the normal temperature, suggesting that turion germination was influenced by temperature, which provided a better explanation for the winter growth habit and the phenomenon of outbreak of P. crispus in spring; 3) The of P. crispus floating in the water adsorbed a large amount of suspended solids, usually 5-7 days to add enough weight to sink to the bottom of water bodies, the adsorbed suspension containing certain nutrients can promote the germination of turions. The mechanism of turion germination provides a basis for studies of factors influencing reproduction of P. crispus and is of important practical significance for cultivation of P. crispus.
Potamogeton crispus; Turion; Germination rate; Suspended solids
10.3969/j.issn.1673-3290.2015.01.10
2014-07-16
国家住房和城乡建设部科学技术计划项目(2012-K-79),国家级大学生创新创业训练计划项目(201311998017),江苏省高校自然科学基金项目(13KJD 610003),徐州市科技计划项目(XM12B087)
尹传宝(1991-),男,汉族,安徽宿州人,本科,主要从事水生态修复方面的研究。E-mail:411727715@qq.com
张翠英,讲师,主要从事水生态修复方面的研究。E-mail:cuiyingzhang@163.com
猜你喜欢
真空与低温(2022年6期)2023-01-06
化工管理(2022年14期)2022-12-02
真空与低温(2022年5期)2022-10-13
中国机械工程(2022年8期)2022-05-09
中国特种设备安全(2021年4期)2021-10-13
儿童时代·幸福宝宝(2021年1期)2021-03-29
音乐教育与创作(2019年8期)2019-05-16
劳动保护(2018年8期)2018-09-12
中国卫生标准管理(2015年3期)2016-01-14
油气田环境保护(2015年4期)2015-12-28
