
西南喀斯特典型树种光合特性的季节变化及主要影响因子
2015-12-18 09:35:16付威波宋敏彭晚霞杨钙仁杜虎文丽李莎莎
农业现代化研究 2015年5期
付威波,宋敏,彭晚霞,杨钙仁,杜虎,文丽,李莎莎
(1.广西大学林学院,广西 南宁530004;2.中国科学院亚热带农业生态研究所亚热带农业生态过程重点实验室,湖南 长沙410125;3.中国科学院环江喀斯特生态系统观测研究站,广西 环江547100;4.湖南农业大学农学院草业系,湖南 长沙410128)
我国西南喀斯特地区面积达50 多万km2,是全球喀斯特集中分布区面积最大、岩溶发育最强烈、景观类型复杂、生物多样性丰富、生态系统极为脆弱的典型地区,在全球喀斯特生态系统中占有重要地位[1]。西南喀斯特区土壤贫瘠,水土流失严重,人地矛盾尖锐,“喀斯特”贫困现象严重,石漠化现象日益严峻[2-3],而植被恢复是退化生态系统重建的主要措施[4]。多年生植物在生命周期中要经历各种各样环境因子的季节变化,而植物的生态适应性是对长期变化的环境因子适应的结果。不同季节植物对环境因子的适应特征、生物学特征及生理过程反映了植物各自的持续生长能力、适应能力和遗传特性。光合作用是植物最重要的生理过程之一,且对环境变化极其敏感,光合作用决定了树木碳代谢和生物量之间的关系。所以,研究植物光合作用季节变化有助于了解其水分利用特性、碳同化、物质积累和持续生长能力,为植物生态适应性的研究提供相应的科学依据。
几十年来,国际上对中亚热带地区常绿阔叶林在区系组成、物种多样性、外貌结构、演替、生态系统的结构和功能、物质和能量的流动等方面进行了大量的研究,并积累了许多资料[5-11]。但有关光合生理生态方面的研究还不多。近年来我国对喀斯特地区的植物光合生理的研究多集中在造林树种的日变化特征[12-13]、不同生活型植物幼苗对干旱生境的生理生态适应[14]、石山植物的光响应[15]、青冈栎(Cyclobalanopsis glauca)或九龙藤(Bauhinia championii)等的光合生理特征[16-17]等方面,对喀斯特地区典型植物光合作用的季节变化研究尚少,而植物的演替状态与其内在的光合生理特性有密切的联系[18]。
本文以西南喀斯特滇南安息香、女贞和岩樟3种主要典型植物为材料,用LI-6400 光合测定系统测定光合生理的季节变化,从植物光合生理特征长期季节变化和光合特性中探寻植物的生态适应性对策,为喀斯特地区人工育种造林、管理等提供科学参考,为喀斯特地区退化生态系统的重建和恢复提供相应的理论指导。
1 研究区概况与研究方法
1.1 自然概况
试验设置在中国科学院环江喀斯特农业生态系统观测站森林植物资源引种 园内(24°43′58.9″-24°44′48.8″N,108°18′56.9″-108°19′58.4″E),地势四周高,中间低,海拔最高为647.2 m,最低为272.0 m,坡度较陡,≥25°的坡面占62%,属于典型的桂西北峰丛洼地。属亚热带季风气候区,气候温和、雨水充沛、日照充足、雨热同季。年平均降雨量1 389.1 mm。年均日照1 451 h,年均太阳总辐射量为413.8 kJ/cm2,有效辐射量为211.9 kJ/cm2。研究区域在上世纪70、80 年代受人为活动影响较严重,砍伐、放牧、耕种、火烧等人为干扰导致植被的覆盖率较低。
1.2 实验材料
试验材料选择滇南安息香(Styrax benzoinoides Craib)、女贞(Ligustrum lucidum)和岩樟(Cinnamomum saxatile)3 种喀斯特典型植物,所有试验对象均为自然生长、立地条件均一、长势良好的植株。
1.3 研究方法
采用1、4、7、10 月份的数据进行光合作用季节变化特征分析。季节变化的测定分别于2009 年10月及2010 年1、4、7 月选择晴朗、无风的天气进行,利用美国LI-6400 便携式光合仪于8:00-17:30 时测定,每个树种选取3 株长势相近的植株,每株选取顶端第2 到第5 片完全展开的4 个健康成熟叶作为待测叶片,每小时测定一个轮回。鉴于一次测量周期较长,为了尽量消除不同植物在同一次测量中的光照、温度、空气湿度、CO2浓度差异,采用3 种植物交叉测定的方法。测定净光合速率(Pn,μmol CO2/(m2·s))、胞间CO2浓度(Ci,μmol/mol)、光合有效辐射强度(PAR,μmol/(m2·s))、大气CO2浓度(Ca,μmol CO2/mol)、大气温度(Ta,℃)、大气相对湿度(RH,%)。
1.4 数据处理
测定结果用Excel 2003 进行处理计算, 并用SPSS 16.0 对实测数据进行显著性检验。
2 结果与分析
2.1 植物光合作用的季节变化
喀斯特3 种典型植物净光合速率季节变化特征不同。从图1 来看,滇南安息香、女贞、岩樟的净光合速率季节变化在1 月均处于相对较低的状态;4 月均有所升高,滇南安息香和岩樟达到峰值;7 月女贞达到峰值,其叶片的光合作用也最强,而滇南安息香和岩樟略有下降;10 月女贞和岩樟的净光合速率有所下降,低于1 月份,而滇南安息香有所上升,仅低于4月。整体而言,滇南安息香的净光合速率四季均为最低,且季节变化较小,4 月、10 月光合速率较高,1 月、7 月光合速率较低。说明季节变化对女贞、岩樟光合能力的影响明显大于滇南安息香。女贞和岩樟的净光合速率季节变化相似,岩樟的净光合速率略低于女贞。
2.2 净光合速率日变化

图1 不同树种净光合速率季节变化Fig.1 Seasonal dynamics of Pn for different species
选取叶片已发育成熟又属于生长旺季的7 月观察数据,对滇南安息香、女贞、岩樟3 种典型喀斯特植物光合作用日进程进行分析(图2)。在阳光强烈、高温的环境条件下,3 种植物的净光合速率日变化总的趋势差异较大,11:00 左右光合速率的降低原因主要是云层的遮挡导致光强降低,从11:00 后开始3 种植物都呈明显“双峰”型,出现了明显的光合“午休”现象,但各树种的变化曲线不同。滇南安息香第1 峰出现在12:00,第2 峰则在16:00 出现,第2 峰值比第1峰值高33%左右,而谷底则出现在15:00,最低值是第2 峰值的31%。女贞的第1 峰在12:00 出现,第2峰在16:00,第1 峰值比第2 峰值高近1 倍,基本上属于“一睡不起”型,光合午间降低的程度也较大(仅为第1 峰值的29%)。岩樟的第1 峰也出现在12:00,第2 峰在14:00,第2 峰值与第1 峰值接近。
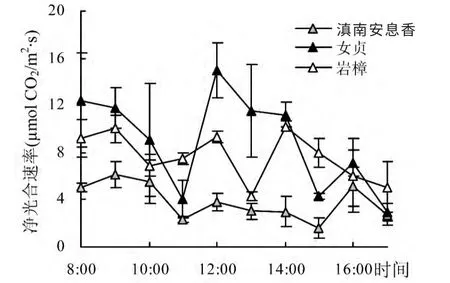
图2 7 月不同树种净光合速率日变化Fig.2 Diurnal variation of Pn for different species in July
2.3 环境因子的季节变化
研究区光合有效辐射、空气相对湿度、空气温度季节变化趋势一致,呈倒“V”型,1 月三者均最低,分别为294.41 μmol/(m2·s)、50.3%和17.6 ℃,随着时间的推移,逐渐升高,7 月达到峰值,分别达到797.27 μmol/(m2·s)、55.41%和36.7 ℃,随后降低。1 月和10月的光合有效辐射、4 月和7 月的空气温度相差幅度不大,空气相对湿度整年变化幅度不大。空气CO2含量在1 月最高,为406.30 μmol/mol,随着时间的推移,呈先降低、后升高、再下降的趋势,4 月最低,为391.76 μmol/mol(图3)。

图3 光合有效辐射、空气温度、空气相对湿度和空气CO2含量的季节变化Fig.3 The seasonal dynamics of PAR、Ta、RH and Ca
2.4 不同季节净光合速率与生理生态因子的关系
本文采用1 月、4 月、7 月、10 月测定的净光合速率数据,并对相关生理生态因子与净光合速率的关系进行相关分析。不同季节,不同树种净光合速率的影响因子不同。由表1 可看出,1 月女贞和岩樟的净光合速率与蒸腾速率和光合有效辐射极显著相关(P<0.01),滇南安息香的净光合速率与蒸腾速率、光合有效辐射显著相关(P<0.05),岩樟还与气孔导度显著相关(P<0.05)。胞间CO2含量、空气湿度与净光合速率均为负相关,但均未达到显著水平。这表明春季的光合有效辐射、蒸腾速率、气孔导度等生理生态因子与滇南安息香、女贞及岩樟的光合作用的关系都很密切。
在4 月,岩樟的净光合速率与光合有效辐射、蒸腾速率极显著相关(P<0.01);女贞的净光合速率与光合有效辐射极显著相关(P<0.01),与胞间CO2含量显著相关(P<0.05);而滇南安息香的净光合速率与各因子相关度均不高。各植物的蒸腾速率与净光合速率相关系数均不一样,差异较大,岩樟净光合速率的相关系数最大,为0.765,而滇南安息香的相关系数最小,仅为-0.007。说明4 月光合有效辐射、蒸腾速率是岩樟光合作用的主要影响因子,光合有效辐射和胞间CO2含量对女贞光合作用的影响最大,而相比其他因子,光强对滇南安息香光合作用的影响较大。
在7 月,3 种植物的净光合速率与胞间CO2含量均极显著相关(P<0.01),女贞的净光合速率还与气孔导度显著相关(P<0.05),岩樟与气孔导度为极显著相关(P<0.01);其他因子与净光合速率的相关度不高。这表明7 月滇南安息香、女贞、岩樟的净光合速率主要受胞间CO2含量的影响,而女贞和岩樟的净光合速率还受气孔导度的影响。
在10 月,滇南安息香、女贞和岩樟的净光合速率与光合有效辐射、蒸腾速率极显著相关(P<0.01),与胞间CO2含量显著相关(P<0.05),岩樟的净光合速率还与气孔导度极显著相关(P<0.01),女贞与气孔导度显著相关(P<0.05)。这说明在10 月,蒸腾速率、光合有效辐射和胞间CO2含量是3 种植物净光合速率共同的主要影响因子,气孔导度对岩樟净光合速率的影响大于女贞和滇南安息香。
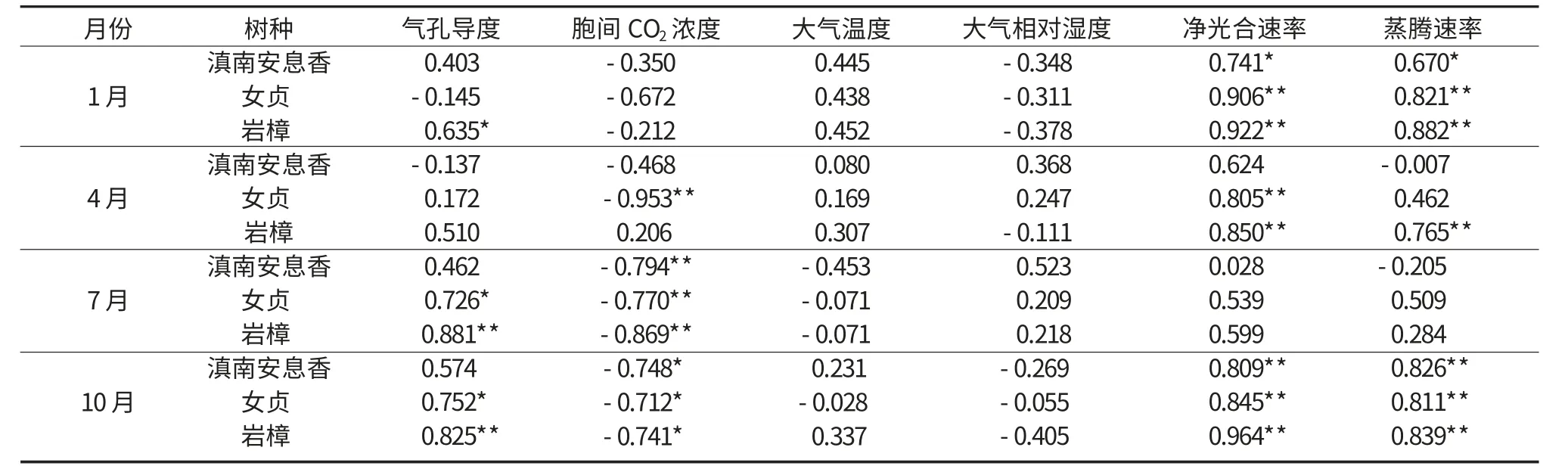
表1 净光合速率与生理生态因子的相关系数Table 1 Correlation coefficients between net photosynthetic rate and physioecological factors
3 讨论
光合作用是植物生长发育中非常重要的生理过程之一,受遗传和环境条件的影响,某些光合特性参数已逐渐成为植物速生丰产及制定栽培措施的科学依据[19]。滇南安息香、女贞和岩樟3 种喀斯特典型植物的净光合速率季节变化差异明显,女贞和岩樟的光合速率季节变化表现为4 月、7 月较高而1 月、10月低,滇南安息香的光合速率季节变化则不明显。不同季节滇南安息香的净光合速率均最低,女贞和岩樟的净光合速率明显高于滇南安息香。树种之间净光合速率的不同,反映出3 种植物光合作用能力的强弱顺序为:女贞>岩樟>滇南安息香。
滇南安息香、女贞和岩樟3 种植物都出现了光合“午休”现象,但各树种的变化曲线不同。11:00 左右光合速率的降低原因主要是云层的遮挡导致光强降低。滇南安息香和女贞两个峰值时间相近,滇南安息香的第二峰高于第一峰,而女贞的第一峰高于第二峰。岩樟的两个峰值出现较早,第一峰和第二峰相近。关于光合午休的成因,不同研究者至今还有不同的看法,主要归纳为气孔限制因素和非气孔限制因素[20-21],哪种因素起主导作用与植物种类及其所处的环境有关[22]。
不同季节影响3 种植物净光合速率的因子不同。1 月光强和蒸腾速率为3 种植物共同的主要影响因子,4 月3 种植物的主要影响因子各不相同,胞间CO2浓度和光合有效辐射是影响女贞的主要影响因子。光强和蒸腾速率是影响岩樟的主要影响因子,而各因子对滇南安息香的影响不显著。7 月影响3 种植物净光合速率的因子主要是气孔导度和胞间CO2含量,10 月蒸腾速率和光合有效辐射仍是3 种植物共同的主要影响因子。结合环境因子的季节变化来看,4 月、7 月的平均气温(31.73 ℃、36.74 ℃) 及光强(490.48 μmol/(m2·s)、797.27 μmol/(m2·s)) 高 于1月、10 月平均气温(17.60 ℃、28.77 ℃) 和光强(294.41 μmol/(m2·s)、286.63 μmol/(m2·s)),气温和光强是导致4 月、7 月影响因子差异的主要原因。在各因子中,光强是不同季节影响3 种植物净光合速率最为主要的因子。
4 结论
光合作用是植物十分复杂的生理过程,与自身遗传因素密切相关,又受到气温、空气、相对湿度、光照强度等外界因子的影响。西南喀斯特地区的气候具有明显的季节特点和分化,使得生长在此的植物的光合特性也发生了季节变化。女贞和岩樟不同季节的光合速率差异较大,而滇南安息香在不同季节下光合速率变化不明显。而在光照、温度、湿度和大气CO2浓度各环境因子中,光照是不同季节影响3种植物净光合速率最为主要的因子。因此植物光合作用也呈现出4、7 月高,1 月、10 月低的季节变化规律。从3 种植物的光合季节变化来看,女贞和岩樟的光合能力明显大于滇南安息香。虽然3 种树种均为喀斯特地区的典型树种,但女贞和岩樟能较好地与西南喀斯特环境相适应,其中女贞的适应性最好,其次为岩樟,而滇南安息香对西南喀斯特环境适应性较差。通过对比3 种西南喀斯特地区典型植物的光合季节变化能够为揭示西南喀斯特地区植物群落的构建机制、喀斯特植被迅速恢复以及喀斯特地区的生态重建提供理论支持。值得指出的是,本试验对3种喀斯特地区典型植物的光合季节变化进行了研究,其光合季节差异的植物自身因素是今后研究的目标。
[1]刘丛强.生物地球化学过程与地表物质循环——西南喀斯特流域侵蚀与生源要素循环[M].北京:科学出版社,2009.
[2]彭晚霞,王克林,宋同清,等.喀斯特脆弱生态系统复合退化控制与重建模式[J].生态学报,2008,28(2):811-820.
[3]刘璐,曾馥平,宋同清,等.喀斯特木论自然保护区土壤养分的空间变异特征[J].应用生态学报,2010,21(7):1667-1673.
[4]张厚华,傅德志,孙谷畴.森林植被恢复重建的理论基础[J].北京林业大学学报,2004,26(1):97-99.
[5]洪伟,林成来,吴承桢,等.福建建溪流域常绿阔叶防护林物种多样性特征研究[J].生物多样性,1999,7(3):208-213.
[6]金则新.浙江仙居俞坑森林群落优势种群结构与分布格局研究[J].武汉植物学研究,2000,18(5):383-389.
[7]贺金生,陈伟烈,李凌浩.中国中亚热带东部常绿阔叶林主要类型的群落多样性特征[J].植物生态学报,1998,22(4):303-311.
[8]Cornelissen JHC.Seedling growth and morphology of the deciduous tree Cornus controversa in simulated forest gap light environments in subtropical China[J].Plant Species Biology,1993,8(1):21-27.
[9]Zhang H C,Hong TC,Bo SW.Studies on biomass and production of the lower subtropical evergreen broad-leaved forest in Heishiding Nature Reserve,China.VI.Distribution,biomass and production of roots[J].Journal of Tropical Ecology,1994,10(2):273-279.
[10]An SQ,Liu M S,Wang Y J,et al.Forest plant communities of the Baohua Mountains,Eastern China[J].Journal of Vegetation Science,2001,12(5):653-658.
[11 殷庆仓.桂西北喀斯特主要先锋树种的光合生理生态特性研究[D].南宁:广西大学,2011.
[12]殷庆仓,宋同清,彭晚霞,等.西南喀斯特地区四种造林树种光合作用日变化及生态适应性研究[J].农业现代化研究,2011,32(3):367-370.
[13]柯世省.天台山3种常绿阔叶树光合特性的季节变化[J].浙江林业科技,2004(5):8-12.
[14]刘长成,刘玉国,郭柯.四种不同生活型植物幼苗对喀斯特生境干旱的生理生态适应性[J].植物生态学报,2011,35(10):1070-1082.
[15]张中峰,黄玉清,莫凌,等.岩溶区4种石山植物光合作用的光响应[J].西北林学院学报,2009,24(1):44-48.
[16]张中峰,黄玉清,莫凌,等.桂林岩溶区青冈栎光合速率与环境因子关系初步研究[J].广西植物,2008,28(4):478-482.
[17]莫凌,黄玉清,覃家科,等.岩溶植物青冈和九龙藤叶片光学特性日变化研究[J].广西科学,2008,15(2):189-194.
[18]梁春,林植芳,孔国辉.不同光强下生长的亚热带树苗的光合-光响应特性的比较[J].应用生态学报,1997,8(1):7-11.
[19]苏培玺,张立新,杜明武,等.胡杨不同叶形光合特性、水分利用效率及其对加富CO2的响应[J].植物生态学报,2003,27(1):34-40.
[20]Xu D Q,Shen Y G.Diurnal variations in the photosynthetic efficiency in plants[J].Acta Phytophysiologica Sinica,1997,23(4):410-416.
[21] Farquhar G D, Sharkey T D. Stomatal conductance and photosynthesis[J]. Annual Review of Plant Physiology, 1982(33): 317-345.
[22]陈贤田,柯世省.茶树光合“午休”的原因分析[J].浙江林业科技,2002,22(3):80-83.
猜你喜欢
读者(2023年2期)2023-02-20 08:10:40
云南社会科学(2021年4期)2021-08-14 01:47:10
云南医药(2021年3期)2021-07-21 05:40:30
Asian Herpetological Research(2021年2期)2021-06-30 14:03:34
热带亚热带植物学报(2020年5期)2020-10-09 14:32:02
化学教与学(2019年4期)2019-05-14 04:46:40
基层中医药(2018年1期)2018-03-01 07:36:25
现代园艺(2017年11期)2017-06-28 11:32:18
上海农业学报(2016年2期)2016-10-27 00:49:49
应用化工(2014年1期)2014-08-16 13:34:08
