
山羊ZP3基因的cDNA克隆、序列分析及组织表达研究
2015-12-17 03:10卢建远杨珂伟字向东
西南民族大学学报(自然科学版) 2015年4期
刘 霜,卢建远,马 力,夏 威,胡 亮,罗 斌,杨珂伟,字向东
(西南民族大学国家民委动物科学重点实验室,四川 成都 610041)
山羊ZP3基因的cDNA克隆、序列分析及组织表达研究
刘 霜,卢建远,马 力,夏 威,胡 亮,罗 斌,杨珂伟,字向东
(西南民族大学国家民委动物科学重点实验室,四川 成都 610041)
对高繁金堂黑山羊和低繁藏山羊ZP3基因cDNA进行克隆、序列分析,并采用Real-time PCR技术对其在发情前期母羊卵巢组织的表达进行研究.结果表明:山羊ZP3基因编码区全长为1269bp,共编码422个氨基酸,两品种间有2处碱基差异,但未导致氨基酸的差异.山羊的ZP3基因在卵巢表达量高,与其他组织差异极显著(P<0.01),但两个品种间差异不显著(P>0.05).说明ZP3基因主要在卵巢中表达,但可能不是影响山羊多羔性状的主基因.
山羊;ZP3;克隆;序列分析;qPCR
透明带(zona pellucida,ZP)是卵母细胞周围一层糖蛋白基质,该家族有ZP1、ZP2、ZP3和ZP4四种糖蛋白[1-2],在哺乳动物精卵识别、结合、穿透、胚胎着床等过程中有着关键的作用[3-5].人、鼠、鸡等中已证实,ZP3蛋白是精子第一受体[6-8],精卵结合与ZP3糖蛋白的O-糖基化位点息息相关[9].重组人的ZP3同样能诱导顶体反应,使人的精子和仓鼠透明带融合[10-11].同时,研究表明敲除ZP3基因的小鼠不能形成透明带[12].可见ZP3基因对卵透明带在精卵结合过程具有极为重要的作用.
目前,国内外有关山羊多胎机制的研究主要集中在绵羊上,已证实与卵泡发育和排卵数的突变基因,如骨形态发生蛋白-15(BMP15)和及其受体-1B (BMPR1B)等,但结果表明这些基因不是控制山羊多胎的主基因[13-14].对与受精过程密切相关的山羊ZP3基因的研究则未见报导.金堂黑山羊是优良的多胎(平均产羔率240%)肉山羊品种[15],而藏山羊则是单胎肉山羊品种[16].因此,本研究以多胎金堂黑山羊和单胎藏山羊为研究对象,首次对不同产羔率的山羊品种的ZP3基因进行cDNA克隆、序列分析和组织表达研究,以期从受精环节探讨山羊多胎性状的分子调控机制奠定一定的理论基础.
1 材料与方法
1.1 试验动物及样品采集
选用5只健康成年雌性四川金堂黑山羊和5只四川理县藏山羊,在同期发情取出阴道栓塞(CIDR)后40h,屠宰羊只,即刻采集卵巢、输卵管、子宫、垂体等组织,液氮冻存备用.
1.2 主要试剂
反转录试剂盒购自FERMENTAS(MBI)公司;2 ×Taq PCR Master Mix、感受态细胞均购自购自康迪生物技术有限公司;氨苄青霉素(Ampr)、克隆载体pMD-19 Vector购自大连宝生物公司;Sso Advanced TMSYBR©Green Super mix,购自伯乐公司.
1.3 总RNA的提取和cDNA的合成
采用Trizol法提取金堂黑山羊和藏山羊卵巢、输卵管、子宫、垂体组织中的总RNA,按Fermentas (MBI)反转录试剂盒进行cDNA的合成,保存备用.
1.4 引物设计及目的基因PCR扩增
根据NCBI上已收录的牛ZP3基因mRNA序列(NM173974),设计克隆引物和qPCR引物(表1). PCR扩增程序:95.0℃预变性5 min;94.0℃变性40s,60.0℃、退火40s;72.0℃延伸1 min,32个循环;最后72.0℃延伸5 min;4.0℃保存.

表1 ZP3克隆及定量引物序列Table 1 Primer pairs of ZP3 for cloning and qPCR
1.5 目的基因的克隆、鉴定及测序
PCR产物电泳检测,连接,转化.取菌液PCR鉴定,挑选阳性克隆菌液2mL送上海英俊生物技术有限公司测序.
1.6 山羊ZP3基因的生物信息学分析
利用DNAman软件对获得的上下游序列进行拼接.从NCBI上检索的人(NM_001110354)、牛(NM_ 173974)、牦牛(GQ856646)、猪(NM_213893)、鼠(NM _011776)、鸡(NM_204389)的ZP3基因相应的核苷酸序列进行多重对比,构建物种分子进化树.用NCBI在线程序(http://www.ncbi.nlm.nih.gov/gorf/ gorf.html)对所得两种山羊ZP3序列进行ORF查找,将CDS区翻译成氨基酸序列.ProtParam程序分析ZP3蛋白的一级结构,理化参数;ProtScal程序分析山羊ZP3蛋白疏水性;TMHMM2.0 Server程序进行跨膜区预测;SignalP 4.0 Server程序预测信号肽;NetOGlyc4.0程序分析O-糖基化位点;GOR程序预测蛋白质二级结构;Phyre2程序预测蛋白质三级结构.
1.7 山羊ZP3基因系统发育树的构建
通过DNAman、DNAstar等生物分析软件对两品种ZP3基因序列进行同源性比对,并利用生物软件MAGA 6.0构建ZP3基因的分子系统进化树.
1.8 qPCR
qPCR反应体系为10μL:ZP3上下游引物(表1)各0.8μL,SsoAdvancedTM SYBR©Green Super mix 5μL,ddH2O 2.9μL,cDNA模板0.5μL.反应程序: 95.0℃预变性3min,95.0℃/10s、57.6℃/20s,35个循环.每个样品为3个重复,并且设置不加模板为对照,取平均值进行数据分析.
1.9 统计分析
qPCR所得数据用Pfaffl法[17]分析ZP3在金堂黑山羊和藏山羊卵巢组织及其他组织的相对表达量,RE=(1+E ref)Ctre/(1+E target)Cttarget.两品种ZP3基因表达量用t检验法进行显著性差异分析.
2 结果
2.1 藏山羊和金堂黑山羊ZP3基因PCR扩增结果
以金堂黑山羊和藏山羊总cDNA为模板,扩增ZP3基因,得到约1300bp左右的片段,与预期相符,并以菌液作为模板进行PCR扩增,所得片段大小也与预期相符(图1).
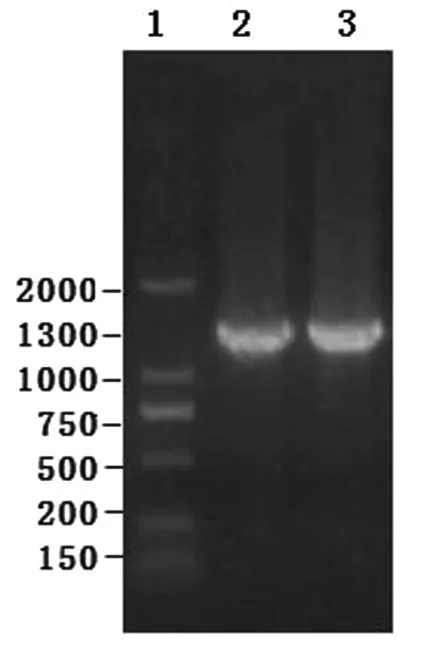
图1 ZP3基因PCR产物电泳检测1.DNA Marker DL2000;2、3.金堂黑山羊和藏山羊ZP3基因扩增结果Fig.1 Electrophoresis results of ZP3 PCR product1.DNA marker DL2000;2,3.ZP3 of Jintang black goat and Tibetan goat
2.2 金堂黑山羊和藏山羊ZP3基因CDS序列分析
用DNAman软件对测序所得ZP3序列拼,得到1269bp同样长度的片段.金堂黑山羊与藏山羊基因编码区的同源性为99.85%,均长1269bp,编码422个氨基酸.金堂黑山羊和藏山羊两品种核苷酸序列有两处不同,第356位C→T,第581位A→G,但是核苷酸的差异没有引起氨基酸的差异.金堂黑山羊ZP3基因CDS区的核苷酸序列与藏山羊及NCBI中牛(NM_ 173974)、牦牛(GQ856646)、猪(NM_213893)、鼠(NM _011776)、鸡(NM_204389)进行对比,相应序列间的一致性分别为99.8%、96%、95%、86%、71.5%、54.6%.
利用MEGA6.0构建系统进化树(图2),藏山羊和金堂黑山羊先聚为一类,牛与牦牛聚为一类再与山羊聚为一类,然后和猪,鼠聚为一类,最后和鸡聚成一大类,这与哺乳动物的进化程度保持一致. 2.3 金堂黑山羊和藏山羊ZP3蛋白的结构特征

图2 基于核苷酸序列的金堂黑山羊和藏山羊ZP3基因的系统进化树Fig.2 Phylogenetic tree of ZP3 gene of Jintang black goat and Tibetan goat
Protparam分析结果表明,金堂黑山羊和藏山羊蛋白的分子式为C2074H3260N588O602S23,相对分子质量为46801.6,理论等电点PI为7.19,半衰期为4.4h,不稳定参数为50.01,属不稳定蛋白,脂肪系数82.84.疏水性分析,显示大多数氨基酸为亲水性氨基酸,判定ZP3蛋白为亲水性蛋白.SignalP3.0信号肽预测,ZP3基因第1-22(Met-Pro)氨基酸序列是信号肽位点,信号肽裂解点位于第22位Pro和第23位Glu之间,属分泌型蛋白.TMHMM 2.0进行跨膜区分析,ZP3蛋白只存在一个跨膜区,位于氨基酸第382-404之间.NetOGlyc4.0分析,ZP3基因存在10个O-糖基化位点.
GOR程序在线预测ZP3蛋白二级结构,结果显示ZP3蛋白包括α螺旋、β折叠、无规卷曲,分别占靶蛋白的2.13%、34.36%和63.51%,由此可见无规卷曲在ZP3蛋白二级结构中有重要作用.PHYRE2.0预测ZP3蛋白的三级结构(图3),残基建模覆盖率63% (266个氨基酸残基),置信度达100%,说明ZP3蛋白与模板蛋白的空间结构比较接近,适合的空间结构比例大. 2.4 组织差异表达分析

图3 ZP3蛋白三级结构预测Fig.3 Prediction for the tertiary structure of ZP3 protein
对ZP3基因在金堂黑山羊和藏山羊4个组织中的mRNA表达量的分析结果表明:ZP3基因mRNA在山羊卵巢、子宫、输卵管、垂体中均有表达,但在卵巢表达量最高,与其他组织差异极显著(P<0.01,图4),而两品种之间差异不显著(P>0.05).

图4 ZP3基因在金堂黑山羊和藏山羊不同组织中的相对表达量(Mean±SE)Fig.4 Expression of ZP3 gene in different tissues of Jintang black goat and Tibetan goat(Mean±SE)
3 讨论
精子与ZP3受体结合后,形成受体配体复合物,从而激发精子发生顶体反应和透明带反应.在精卵识别和粘附过程中,ZP3蛋白的O-连接糖基化位点具有极关键作用[18].O-聚糖俗称粘蛋白,与细胞的识别、粘附及细胞凋亡等生命过程密切相关[19].小鼠定点突变实验表明,ZP3蛋白的Ser332和Ser334位的丝氨酸的O-糖基化位点是结合精子的必须基团[20].而研究者在生物进化过程中发现,Ser332和Ser334附近的氨基酸发生改变,导致种的特异性,从而影响精卵结合[21].本研究对多胎山羊品种(金堂黑山羊)和单胎山羊品种(藏山羊)ZP3基因的克隆、生物信息学分析表明,两品种ZP3基因cDNA序列一致性达到99.8%,与牛和牦牛的序列一致性达到96%[22],说明山羊ZP3基因cDNA序列比较保守.山羊的ZP3基因编码区序列与其他物种同源性很高,在哺乳动物中较保守,分子系统进化树构建结果也符合进化系统.山羊ZP3蛋白分子量为46.80kD,与普通牛(46.55kD)和牦牛(46.62kD)蛋白分子量差异不大[22].目前,有关卵透明带的研究主要集中在受精过程时精子的特异性位点识别及相关分子机制[23].本研究发现两个山羊品种的氨基酸序列都为10个O-糖基化位点,说明O-糖基化结合位点不会导致金堂黑山羊和藏山羊在受精环节的差异.
哺乳动物中,人和鼠的ZP3蛋白主要在卵巢组织中表达,在其他组织只有微量表达[12,24];鱼类中,红鲫和中华鲟等的ZP3蛋白在卵巢中的表达量极显著于心脏、肝脏、脑等[25-26].本实验荧光定量PCR结果显示,金堂黑山羊和藏山羊ZP3基因在子宫、输卵管和垂体均有极低表达,但是在卵巢中的表达量显著高于其它组织(P<0.01),说明ZP3基因在卵巢组织具有表达特异性,体现ZP3基因在透明带中的重要作用.但是两个品种间差异不显著(P>0.05),说明ZP3基因不是影响山羊产羔性状的主要原因.
4 结论
本试验首次克隆了山羊ZP3基因,多胎金堂黑山羊与单胎藏山羊ZP3基因CDS编码区存在2处核苷酸差异,但未导致氨基酸的差异.山羊和其他物种的ZP3基因的同源性高,在生物进化上高度保守.山羊ZP3基因包含一个1269bp的开放阅读框,编码422个氨基酸.ZP3基因在两山羊品种间的表达无显著差异.
[1]SUTOVSKY PG,MANANDHAR,GMECAULEY TC,et al.Proteasomal interference prevents zona pellucida penetration and fertilization in mammals[J].Biol Reprod,2004,71(5):1625-1637.
[2]CANDACL LK,WILLIAM FH,JOL LS,et al.Characterization of zona pellucida glycoprotein 3(ZP3)and ZP2 binding sites on acrosome intact mouse sperm[J].Biol Reprod,2002,66(6):1585-1595.
[3]VANROOSE G,NAUWYNCK H,SOOM AV,et al.Structural aspects of the zona pellucida of in vitro-produced bovine embryos:a scanning electron and confocal laser scanning microscopic study[J].Biol Reprod,2000,62(2):463-469.
[4]SUN Y,NELSON BJ,GREMINGER MA.Investigating protein structure change in the zona pellucida with a microrobotic system[J].Int J Robot Res,2005,24(2):211-218.
[5]DONG KW,CHI TF,JUAN YW,et al.Characterization of the biologic activities of a recombinant human zona pellucida protein 3 expressed in human ovarian teratocarcinoma(PA-1)cells[J].Am J Obstet Gynecol,2001,184(5):835-843.
[6]CABAKKERO-CAMPO P,CHIRINOS M,FAN XJ,et al.Biological effects of recombinant human zona pellucida proteins on sperm function [J].Biol Reprod,2006,74(4):760-768.
[7]BAUSEK N,RUCKENBAUER HH,PFEIFER S,et al.Interaction of sperm with purified native chicken ZP1 and ZPC proteins[J].Biol Reprod,2004,71(2):684-690.
[8]JIN M,FUJIWARA E,KAKIUCHI Y,et al.Most fertilizing mouse spermatozoa begin their acrosome reaction before contact with the zona pellucida during in vitro fertilization[J].Proc Natl Acad Sci USA,2011,108(12):4892-4896.
[9]JOVINE L,DARIE CC,LITSCHER ES,et al.Zona pellucida domain proteins[J].Annu Rev Biochem,2005,74:83-114.
[10]CLIAMBERLIN ML,LEAN J.Human homolog of the mouse sperm receptor[J].Proc Natl Acad Sci USA,1990,87(16):6014-6018.
[11]VAN DAIN M,POLMAN JEM,GINNKEN K,et al.Recombinant human zona pellucida protein produced by Chinese hamster ovary cells induces the human sperm acrosome reaction and promotes sperm egg fusion[J].Biol Reprod,1994,51(4):607-617.
[12]WASSARMAN PM,JOVINE L,LITSCHER ES.Mouse zona pellucida genes and glycoproteins[J].Cytogenet Genome Res,2003,105(2-4):228-234.
[13]POLLEY S,DE S,BATABYAL S,et al.Polymorphism of fecundity genes(BMPR1B,BMP15 and GDF9)in the Indian prolific Black Bengal goat[J].Small Ruminant Res,2009,85(2-3):122-129.
[14]何远清,储明星,王金玉,等.6个山羊品种高繁殖力候选基因BMP15多态性研究[J].安徽农业大学学报,2006,33(1):61-64.
[15]许德贵,张家明,唐诗军,等.金堂黑山羊生产性能研究[J].四川畜牧兽医,2006,33(11):25-26.
[16]王杰,王永,欧阳熙,等.藏山羊研究[J].中国畜牧杂志,1993,29 (1):10-13.
[17]PAFFL MW.A new mathematical model for relative quantification in real-time RT-PCR[J].Nucleic Acids Res,2001,29(9):e45.
[18]LITSCHER ES,WASSARMAN PM.Characterization of a mouse ZP3 derived glycopeptide,gp55,that exhibits sperm receptor and acrosome reaction-inducing activity in vitro[J].Biochemistry,1996,35(13): 3980-3985.
[19]STWORA WOJCZYK MM,KISSINGER JC,SOITALNIK SL,et al.O -glycosylation in Toxoplasm gondii:identification and analysis of a family of UDP-GalNAc:polypeptide N-acetylgalactosaminyltransferases[J].Int J Parasitol,2004,34(3):309-322.
[20]CHEN J,LITSCHER ES,WASSARMAN PM.Inactivation of the mouse sperm receptor,mZP3,by site-directed mutagenesis of individual serine residues located at the combining site for sperm[J].Proc Natl Acad Sci USA,1998,95(11):6193-6197.
[21]CONNER SJ,HUGHES DC.Analysis of fish ZP1/ZPB homologous genes-evidence for both genome duplication and species-specific amplification models of evolution[J].Reproduction,2003,126(3):347-352.
[22]徐华伟,字向东,黄磊,等.牦牛ZP3基因的克隆及蛋白质结构的预测[J].生物技术通报,2010,20:157-161.
[23]GUPTA SK,BHANDDARI B,SHRESTHA A,et al.Mammalian zona pellucida glycoproteins:structure and function during fertilization[J]. Cell Tissue Res,2012,349(3):665-678.
[24]KUBO H,KAWANO T,TSUBUKI S,et al.A major glycoprotein of Xenopus egg vitelline envelope,gp41,is a frog homolog of mammalian ZP3 [J].Dev Growth Differ,1997,39(4):405-417.
[25]SHI J,PENG K,SHENG J,et al.A specific genomic organization and a novel promoter sequence for both ZP2 and ZP3 gene expressions in thePingxiangredtransparentcruciancarp,Carassiusauratus var.pingxiangnensis[J].Comp Biochem Phys D,2013,8(4):275-282.
[26]SUN Y,YU H,ZHANG Q,et al.Molecular characterization and expression pattern of two zona pellucida genes in half-smooth tongue sole (Cynoglossus semilaevis)[J].Comp Biochem Phys B,2010,155(3): 316-321.
(责任编辑:李建忠,付强,张阳,罗敏;英文编辑:周序林,郑玉才)
cDNACloning,sequence analysis and tissue expression of goat ZP3 gene
LIU Shuang,LU Jian-yuan,MA Li,XIA Wei,HU Liang,LUO Bin,YANG Ke-wei,ZI Xiang-dong
(The Key Laboratory of Animal Science of State Ethnic Affairs Commission,Southwest University for Nationalities,Chengdu 610041,P.R.C.)
This study was carried out for ZP3 gene cloning and sequence analysis on non-prolific Tibetan goats and prolific Jintang black goats,and its mRNA expression levels in ovary at pro-estrus were determined by real-time PCR.The result showed that the coding region sequence(CDS)of goat ZP3 gene was 1269bp long,encoding 422 amino acids.There were 2 basic differences between the two breeds without amino acid change.The mRNA expression level was higher in ovary than in other tissues (P<0.01),but there was no differential expression between the two breeds(P>0.05).The result shows that ZP3 gene is highly expressed in ovary,but it may not be the major gene to determine the kidding rate in goat.
goat;ZP3;cloning;sequence analysis;qPCR
S814;S827
A
2095-4271(2015)04-0412-05
10.11920/xnmdzk.2015.04.004
2015-04-01
字向东(1963-),男,白族,云南云龙人,教授,研究方向:动物遗传育种与繁殖,E-mail:zixd@sina.com
四川省应用基础项目(2013JY0043);西南民族大学研究生创新型科研项目(CX2014SZ69)
猜你喜欢
云南畜牧兽医(2022年4期)2022-08-19
民间文学(2022年1期)2022-03-31
四川劳动保障(2021年9期)2022-01-18
云南畜牧兽医(2021年6期)2021-12-24
今日农业(2021年15期)2021-11-26
云南畜牧兽医(2021年5期)2021-10-27
民间文学(2018年3期)2018-04-28
作文周刊·小学一年级版(2017年46期)2018-01-16
小天使·一年级语数英综合(2017年4期)2017-04-18
中国畜牧兽医文摘(2015年9期)2015-12-29
