
豫西北奥陶系马家沟组遗迹化石及其沉积环境①
2015-12-08 06:15牛永斌单婷婷董小波高文秀
沉积学报 2015年2期
牛永斌 单婷婷 董小波 周 硕 高文秀
(1.河南理工大学资源环境学院 河南焦作 454000;2.河南省生物遗迹与成矿过程重点实验室 河南焦作 454000;3.河南理工大学现代教育中心 河南焦作 454000)
遗迹化石是在某种沉积条件下形成的生物沉积构造,它指示某种特定的沉积条件。在实体化石贫乏或完全缺失的情况下,是进行沉积古环境分析非常实用的相标志[1-5]。特别是对于华北盆地早古生代的海相碳酸盐岩沉积区,完整的生物实体化石相对缺乏,而遗迹化石非常丰富,利用遗迹化石进行沉积古环境分析具有得天独厚的条件[6-9]。
研究区主要包括豫西北济源—焦作—安阳林州一线,位于 35°12'53″~35°25'5″N,113°6'27″~113°22'38″E(图1);构造上隶属于华北盆地太行山分区,区内奥陶系碳酸盐岩非常发育。前人对于区内奥陶系的沉积微环境,现有的研究成果非常薄弱;且多从华北或河南省等较大范围内对奥陶系沉积古环境进行了初步研究。如叶连俊等[10]、王英华等[11]、冯增昭等[12]利用岩石组合序列对华北奥陶系沉积建造、古环境和古地理进行了研究;刘印环等[13]综合岩性组合和古生信息对河南省奥陶系岩相古地理进行了研究;胡斌等[14-15]利用岩性信息、沉积构造、古生物实体化石和遗迹化石对焦作青龙峡景区奥陶系马家沟组的沉积环境和沉积演化进行了初步研究。笔者在国家自然科学基金项目的支持下,利用2011~2013近三年的时间对豫西北奥陶系马家沟11处剖面中的8处剖面进行了详细的测定与描述,目前共鉴定和描述遗迹化石10属17种,这对补充华北奥陶系遗迹化石研究和沉积古环境分析具有重要的借鉴意义。
1 地层概况
研究区奥陶系地层出露良好,厚达800 m以上[16],上覆石炭系本溪组地层,呈平行不整合接触;下伏寒武系三山子组地层,亦呈平行不整合接触;自下而上依据岩性差异可以划分为七个岩性段[17]。笔者带领项目组人员对博爱后寨(一至七段)、西小底(第七段)、影视城(第五段)、东交口(二至六段)、穆家寨(二段至五段)、葡萄峪(一至三段)、台道(四至六段)、王窑(六至七段)、青龙峡(一至七段)等八处剖面进行详细的观测(图2),各段岩石学特征如下:
马一段:厚度较稳定,岩性为含砾泥质白云岩、页状泥质白云岩、姜黄色疙瘩状泥质灰岩夹中晶白云质灰岩等,下部见鸟眼构造。以灰色、深灰色中厚层状泥晶灰岩作为与马二段的分界,未见实体化石和遗迹化石。
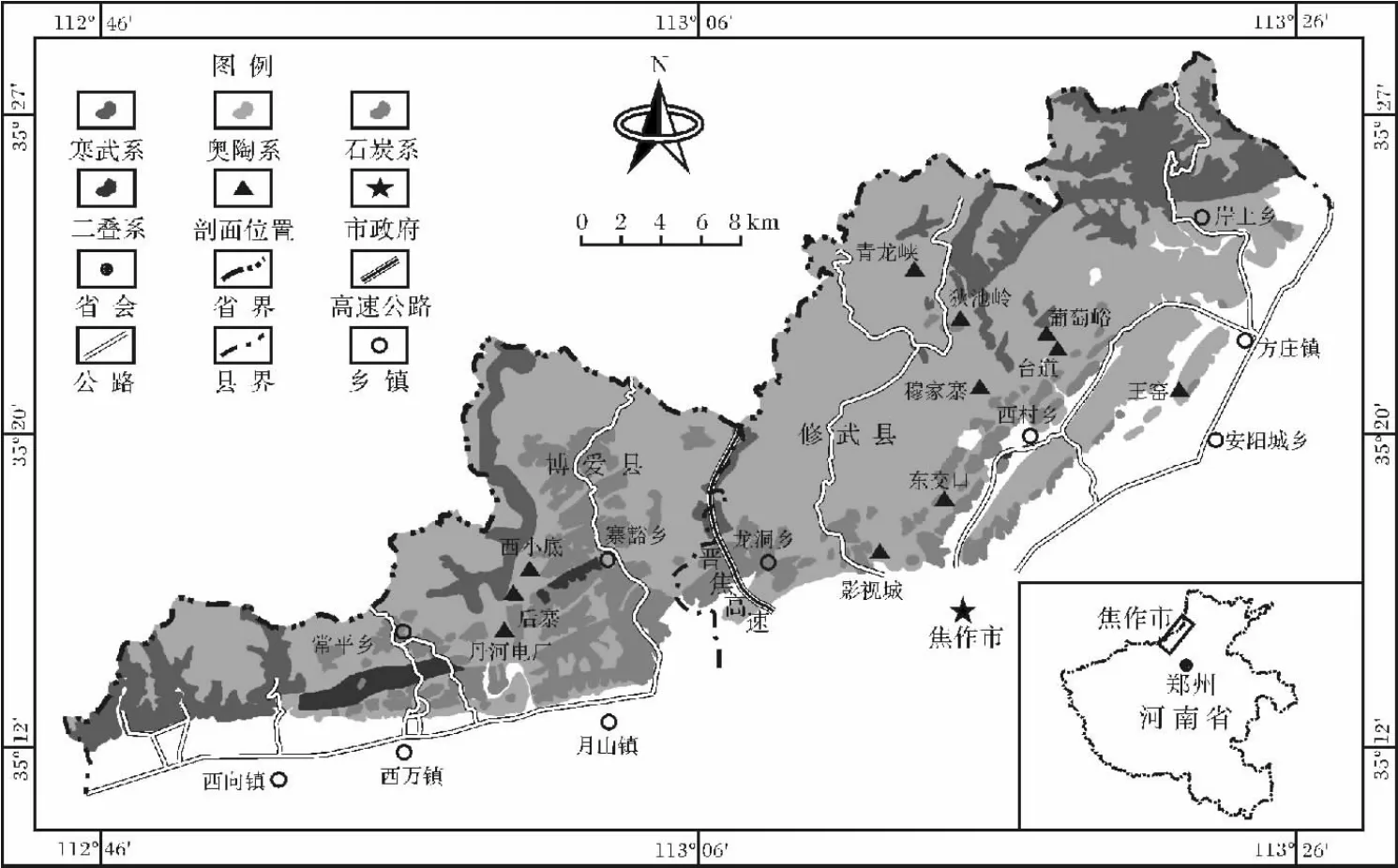
图1 研究区地理位置图Fig.1 Location of the study area
马二段:区内分布稳定,厚度较大。下部为深灰色角砾状灰岩,中间为深灰色中厚层状泥灰岩夹中薄层状灰质白云岩,上部为深灰色厚层状灰岩,白云质灰岩;水平纹理发育,中、下部含少量生物化石碎屑,上部含少量甲壳动物碎屑。葡萄峪剖面、青铜峡剖面上部见 Psilonichnus、Thalassinoides、Helminthopsis、Planolites和Palaeophycus遗迹化石。
马三段:下部为浅灰、灰红、黄褐色角砾状白云岩,以此为标志与马二段分界,发育石盐假晶和鸟眼构造;上部为中厚层状白云质灰岩,层厚向上变薄,白云质含量逐渐增加;实体化石较少,遗迹化石Thalassinoides、Helminthopsis极为发育,见 Planolites、Palaeophycus、Chondrites和 Gordia遗迹化石。
马四段:下部为灰黄色薄层状泥灰岩,泥质白云岩向上过渡到中厚层状灰岩,与马三段界限明显;上部为中厚层状白云质灰岩。马四段见大量实体化石,化石种类主要有腕足类、腹足类、有孔虫类、棘皮类、介形虫类、牙形石,局部见甲壳类和三叶虫类化石碎屑[14-15]。下部遗迹化石 Thalassinoides、Helminthopsis较为发育,见 Planolites、Palaeophycus,上部见 Rhizocorallium、Phycodes(青龙峡剖面)遗迹化石。
马五段:本段岩性主要为深灰色—灰黑色厚层—巨厚层石灰岩,水平层理较为发育;生物碎屑含量相对于马四段有所减少,但是有孔虫、头足类、腹足类仍有发现。遗迹化石分布特征是下部发育Thalassinoides、Helminthopsis和 Chondrites遗迹化石,中部发育Thalassinoides、Helminthopsis遗迹化石,上部发育Chondrites和Scolicia遗迹化石。
马六段:下部为灰色中层灰质白云岩、斑状白云质灰岩,发育Thalassinoides、Helminthopsis遗迹化石,见鸟眼构造与石膏假象;上部为中层白云质灰岩夹角砾状灰岩,水平层理发育,顶部见Helminthopsis、Chondrites、Planolites和 Gordia遗迹化石。
马七段:本段底部为黑色藻叠层灰岩,向上逐渐过多为亮晶砂屑灰岩和白云质灰岩[14-15],未见遗迹化石,水平层理、波状层理发育;上部为灰黑色厚层状砂屑灰岩,灰黑色生物碎屑灰岩,见海百合实体化石,遗迹化石Thalassinoides发育。
2 遗迹化石组成和分类
豫西北奥陶系马家沟组碳酸盐岩中遗迹化石经过鉴定和系统分析,目前共识别出10属17种,综合已鉴定遗迹化石造迹生物的行为习性和所能反应的生态习性特征,可划分为进食迹、觅食迹和居住迹三个生态类型,各生态类型的围岩特征与分布层段如表1。

图2 豫西北奥陶系马家沟综合柱状图Fig.2 Synthetic column of Ordovician Majiagou Formation in the western Henan province

表1 豫西北奥陶系马家沟组遗迹化石统计表Table 1 The trace fossils statistics of Ordovician Majiagou Formation in the western Henan province
3 遗迹化石鉴定与描述
居住迹(Domichnia)
螃蟹迹属Ichnogenus Psilonichnus Curran and Pemberton,1984
分叉形螃蟹迹Psilonichnus upsilon
(图版Ⅰ-a)
描述:分形螃蟹迹居住潜穴多呈近直立柱状,多为Y形、J形分叉,有时亦为U形分叉,潜穴内无衬壁[18],潜穴直径介于 3~8 mm,长约 10~30 mm。潜穴充填物多为细粒灰泥质或白云石颗粒。
讨论:这类遗迹化石直立—近直立的外部形态反映的多为生物的居住潜穴(即居住迹)。前人研究认为这类遗迹化石的造迹生物主要为甲壳类或其他生态习性类似的生物,在碎屑岩沉积环境中常见于前滨最上部、海滩滨后等沉积环境,在碳酸盐岩沉积环境主要出现在潮上带下部和潮间带上部(高潮坪)的沉积环境,反应中低能的滨浅海环境[19]。
产地层位:焦作葡萄峪,奥陶系马家沟组二段上部。
海生迹属 Ichnogenus Thalassinoides Ehrenberg,1944
模式种Thalassinoides callianassa Ehrenberg,1944
(图版Ⅰ-b)
描述:该遗迹种在岩层内部呈网状在三维空间内延伸,具有分枝这一特点明显。其潜穴管延展连续性较好,常见到有潜穴管呈垂直岩层延展而和岩层表面连通。其潜穴管常见Y形分枝和近垂向分支,相互之间经常穿插连接,构成一套在水平方向上延伸较广的网状分布潜穴系统。遗迹化石为内生迹,潜穴管光滑且无衬壁,分支处会略显膨大变粗。研究区此种遗迹化石潜穴直径在0.5~1.5 cm之间,分叉角度较大,围岩多为泥晶生屑灰岩而充填物多为沙晶白云岩,充填物抗风化能力强于围岩,经常可见因风化侵蚀围岩而使潜穴相对凸显于岩层表面。
讨论:因为Thalassinoides在岩层中呈三维空间展布,在岩石表面显露的化石形态多样,但分枝现象很明显,且横向连续性较好。研究区内该化石分布较为广泛,是最为常见的遗迹化石之一。Thalassinoides是一种居住和觅食潜穴,常产出于滨海潮间带附近,指示海岸附近所在[20]。前人研究成果认为Thalassinoides在白垩纪以后的造迹生物主要是甲壳动物的十足目虾类动物[14-15],但在奥陶纪其造迹生物还不确定,因为目前还没有十足目虾玉虾类动物在当时就已存在的证据,研究区Thalassinoides不仅发育在潮间带,也发育在潮下带环境。
产地层位:焦作后寨、东郊口、葡萄峪、王窑、青龙峡,马家沟组二段上部、五段、六段、七段中上部。
水平海生迹 Thalassinoides horizontalis Myrow,1995
(图版Ⅰ-c)
描述:该遗迹种多呈沿着岩层面分布的具有分支的网状结构,其分枝状呈Y字型。潜穴管为亚圆柱形和圆柱形,直径在0.5~1.2 cm之间,其分枝处膨大现象不明显,分枝间距较为均一,所形成的网状空格大小也比较均一。化石潜穴管外壁光滑,未见到内部构饰。其充填物为灰质白云岩,围岩以泥晶灰岩为主,暴露面经风化后使得化石相对浮现于岩层表面。研究区此遗迹化石潜穴中有可见到较大的白云石晶形和少量甲壳动物实体化石。
讨论:研究区Thalassinoides horizontalis与Thalassinoides callianassa相比,其潜穴直径变化不大,构成网状结构的网格大小较均一,化石形态特征比较明显。该遗迹化石主要沿水平方向分布和连通,垂直方向的潜穴管发育不明显。研究区Thalassinoides潜穴中发现的甲壳动物化石,这为甲壳动物参与该类遗迹化石的建造提供了证据。此外,有学者认为分枝处有膨大现象的Thalassinoides是由虾类或者体型类似的生物所造,因为此类造迹生物在潜穴内需要转动身体以调转行进方向,在分叉口就留下了转动身体时磨损潜穴壁的膨大特点。而另一些能够不掉头而后退运动的造迹生物很可能就不会在分叉处留下膨大的潜穴特点[20-22]。
产地层位:焦作葡萄峪、东郊口、王窑、青龙峡,马家沟组三段上部,四、五、六下部,七段。
根珊瑚迹属 Ichnogenus Rhizocorallium Zenker,1836
詹尼斯根珊瑚迹 Rhizocorallium jenense Zenker,1936
(图版Ⅰ-d)
描述:该遗迹种U型潜穴保存完好清晰可辨,整体扁平,管壁明显且内部具有清晰的回填纹构造,两翼管凸于岩石表面。整体与层面近平行保存,化石长约12 cm,两端圆弧状,大体上看向着内部回填纹所指示方向管径变粗。潜穴外径在1~1.5 cm之间变化,内径在0.5~1.2 cm之间,管壁厚度均匀,约0.3 cm左右。围岩为灰黑色泥晶灰岩,充填物呈深灰色,比围岩耐风化。
讨论:该遗迹潜穴具有回填纹构造可能反映Rhizocorallium是一种进食遗迹。Seilacher(1967)认为Rhizocorallium的蹼状构造的方向受到沉积速率或侵蚀速度的控制[20,23],其潜穴管径和潜穴整体大小成正比。Basan和Scott(1979)对Rhizocorallium的构造特征作过补充说明,认为其两翼近平行,之中的蹼状纹层呈同心状前进式,内部充填物和母岩的成分相同,在一些样本的翼管上经常可以见到动物的抓痕[23]。多数Rhizocorallium为内生迹,全部保存在岩石内,围岩被风化侵蚀之后才出现浮起于岩石表面的现象。
产地层位:焦作青龙峡,奥陶系马家沟组马四段中上部。
进食迹(Fodinichnia)
丛藻迹属 Ichnogenus Chondrites Sternberg,1833
B型丛藻迹 Chondrites type-1 Osgood,1970
(图版Ⅰ-e)
描述:B型丛藻迹外部形态为细小的树枝状潜穴系统,其每束分枝沿一条细长的主枝两侧分布为羽状,每个分枝长度较短且不集中,单个遗迹潜穴管互不交叉,在多个遗迹时有互相叠加的情况,整体为较为分散的潜穴群。潜穴直径多在0.1~0.3 cm,每组潜穴管直径均一,分枝之间角度基本相同,为30°左右。潜穴内的充填物与围岩成分和颜色基本相同。
讨论:Chondrites属化石是一种分布范围广、延续时间长的遗迹化石,从前寒武纪到现代,可出现在多种环境中(晋惠娟,1999)。该遗迹化石的造迹生物不明,目前还没有确切发现,普遍认为主要是生活在沉积物界面和水面下方比较深处的食沉积物动物[24-25],是一种高度分支的潜穴系统[26-30]。参考 Osgood(1970)所描述的 Chondrites type-1标本[20],形态上与研究区部分 Chondrites非常接近,因而定为Chondrites type-1。主要在研究区的葡萄峪和青龙峡地区发育,个体在研究区的Chondrites属中是最小的。
产地层位:焦作葡萄峪、青龙峡,奥陶系马家沟组五段底部。
C型丛藻迹 Chondrites type-2 Osgood,1970
(图版Ⅰ-f)
描述:该遗迹种潜穴分枝系统呈树枝状,出现多次分枝的情况,聚集较密呈菊花状;其潜穴分枝结构较为简单,多为双分枝或者三分枝形态,每个分枝夹角为约为30°左右,分枝潜穴的直径在0.2~0.5 cm间,单个分支管长1 cm左右,每个分枝形态类似并且粗细均匀。围岩与充填物相似。
讨论:Chondrites type-2的典型特征是多次分枝和常多方向放射状形如菊花。Chondrites是一种穿相性遗迹化石[29-31],且造迹生物多种,可与多种其他遗迹化石共生,而其充填物性质也随不同环境、不同围岩而有所不同,抗风化强度不尽相同。Bromley和Ekdale(1984)认为Chondrites是形成在缺氧环境中的,甚至比一般任何的生物更能存在于缺氧环境中,其分布主要受控于氧气含量而不仅仅是水深和沉积环境[31]。
产地层位:焦作青龙峡,奥陶系马家沟六段下部。
缠结丛藻迹 Chondrites cf.intricatus(Brongniart,1823)Sternberg,1833
(图版Ⅰ-g)
描述:该遗迹种潜穴分枝从一点大致向一个方向辐射生长,分枝较为密集,出现两次以上分叉。形态上呈半圆状向外辐射。单个潜穴管直径在0.1~0.2 cm,角度不大在20°左右,一簇潜穴管长2 cm左右,单个分支管长不足1 cm。多数为一束潜穴像一个方向辐射,可偶尔出现潜穴分枝围绕一点向四周辐射的现象。
讨论:根据杨式溥对Chondrites intricatus的描述,这类遗迹化石分枝呈辐射状潜穴系统,枝的直径为1 mm,分枝成锐角,常自一点向外侧成近半圆形,枝密集。整体遗迹宽约3 cm[20]。研究区发现的 Chondrites cf.intricatus遗迹化石从分枝的粗细、分枝方式、潜穴排列特征,同Chondrites intricatus较为相似,但又有所不同,故定为Chondrites cf.intricatus(相似种)。此外,研究区发现的该遗迹种与 Chondrites type-2在同层位发育,多分布于岩层面上。该种遗迹化石与Chondrites type-2相比,外形多呈半圆状,每组潜穴管方向分散度较小,且潜穴管直径较为细小,不见360°放射状生长。
产地层位:焦作青龙峡、葡萄峪,奥陶系马家沟组三段,六段下部。
古藻迹属 Ichnogenus Palaeophycus Hall,1847
管状古藻迹 Palaeophycus tubularis Hall,1847
(图版Ⅱ-a)
描述:管状古藻迹潜穴外形多呈直—微弯曲的柱状,且常保存在层面上。管壁清晰可辨、表面光滑没有发现衬饰,长度不大,呈两头收缩变细的特征,多为1~7 cm。薄片观察发现潜穴内部充填物与围岩成分相同,均为泥晶灰岩;潜穴内部充填物较围岩粒度较大。
讨论:Palaeophycus形态变化大,多数表面光滑,充填物没有明显特点。该种和Chondrites保存在同一层面上,可以判断其造迹生物能适应贫氧环境这一特点与Chondrites的造迹生物类似。依据Palaeophycus tubularis直到微弯曲的圆柱形潜穴形态,保存在层面上的特征,其表现出更多的是进食迹的特征,故依据生态习性分类把它归为进食迹。与Planolites相比,Palaeophycus 的主要特征是具有衬壁[15,20]。与Helminthopsis相比,Palaeophycus虽弯曲但不成蛇曲状。
产出层位:焦作葡萄峪、青龙峡,马家沟三段中上部,四段、六段。
赫伯特古藻迹Palaeophycus heberti Saporta,1872
(图版Ⅱ-b)
描述:该遗迹种明显呈圆柱状潜穴,凸出围岩保存,内部充填物被溶蚀而凹陷或中心残余,呈烟囱状。其潜穴形态均一,直径变化在0.3~1.5 cm之间,长度约3~5 cm。管壁光滑,厚度0.3~0.7 cm,没有纹饰,多为直到微弯曲状。分枝现象不常见,常沿层面分布,呈底迹,少数为表迹或内生迹。
讨论:依据 Palaeophycus heberti圆柱形潜穴形态,多凸出围岩保存在层面上,而不是直立—近直立保存的外貌特征,更多呈现的是进食迹的特征。故文中依据生态习性分类把它归为进食迹,而不是居住迹。较厚的衬壁构造是Palaeophycus heberti的明显特征,以此可将其与 Palaeophycus tubularis相区分[14-15,20]。
产出层位:焦作葡萄峪、东郊口、青龙峡,马家沟组二段上部,三、四段。
漫游迹属 Ichnogenus Planolites Nicholson,1873
山地漫游迹 Planolites montanus Richter,1937
(图版Ⅱ-c)
描述:研究区该遗迹种遗迹化石基本沿岩层面分布,潜穴呈亚圆柱形,表面光滑无纹饰,充填物与围岩区分明显,潜穴颜色较深为灰黑色,无明显衬壁。潜穴大多独立分布,没有分枝现象,没有明显交叠现象。潜穴管短且直,长度多在1~4 cm,直径0.2~0.4 cm。
讨论:在 Planolites属中,研究区的 Planolites montanus个体较小。潜穴管多为直或微弯,常见直径小于5 mm,潜穴表面光滑。该遗迹化石模式种常为相互交叉排列,表面光滑或者有纹理,直径可达15 mm[14-15]。切面圆或椭圆,可能为觅食构造,造迹生物为食沉积物的动物,如多毛虫和其他一些蠕虫[20]。此种遗迹化石全球分布范围较广,能够适应各种水深要求,且持续时间较长,从前寒武纪晚期到更新世均有发现。
产地层位:焦作后寨、葡萄峪、王窑,马家沟组三段、四段顶部、五段下部、六段下部。
环形漫游迹 Planolites annularius Walcott,1890
(图版Ⅱ-d)
描述:研究区该遗迹种化石呈水平或略微倾斜状沿层面分布。潜穴多数为柱状,没有衬壁,长约3~6 cm,直径在0.2~0.5 cm之间,表面有环节规则分布,宽窄均匀。一般每厘米见2~3个环形痕迹。
讨论:该种遗迹化石在研究区发现较少,仅在葡萄峪地区马三段地层中见到。且因为普遍经过风化后,部分化石的潜穴表面环节特征不明显。
产地层位:焦作葡萄峪,马家沟组二段。
拟藻迹属 Ichnogenus Phycodes Richter,1850
掌状拟藻迹 Phycodes palmatus Hall,1853
(图版Ⅱ-e)
描述:该遗迹种因形似手掌而得名,表现为簇的潜穴管,一端汇聚一端分散。每个潜穴管为圆柱状,互相之间不相交,且只发现一次分叉,没有在分枝上二次分叉。潜穴管分枝直径0.3~1 cm,长约11 cm,根部分枝长4~7 cm,分支角度有变化,在15°~30°之间。潜穴管壁光滑,整体呈掌状或花束状。其潜穴充填物为泥晶灰岩,研究区有的层位Phycodes潜穴充填物为浅色,有的为深色。
讨论:Phycodes palmatus遗迹种常通过几根分枝形成“掌形”,且分枝较为粗大。前人研究成果一般认为该遗迹化石为某种蠕虫类动物的进食遗迹,多在浅海环境出现[7,14,20]。
产地层位:焦作葡萄峪、青龙峡,奥陶系马家沟组四段中上部。
觅食迹(Pascichnia)
拟蠕形迹属Ichnogenus Helminthopsis Heer,1877
拟蠕形迹未定种Helminthopsis ichnosp.
(图版Ⅱ-f)
描述:该遗迹种沿层面分布,形态变化大。潜穴管直径1~2 cm,长度5~30 cm,充填物颜色较深。单个潜穴不分枝、不穿插,管壁光滑未见衬壁,其弯曲不规则,弯曲角度变化大,且弯曲弧度不确定,可以有良好弧度也可以呈折线弯曲。
讨论:参考前人关于Helminthopsis的研究资料,潜穴直径多呈4~10 cm,外形多为小型—大型弧弯曲[17]。研究区该遗迹化石个体很大,没有发现已定名属种中有相似的,因此作为未定名种的Helminthopsis。与其相比,Gordia的主要不同处其是没有呈现明显的蛇曲状弯曲,而是形态呈圆环状或不封闭,且个体相对较小。
产出层位:焦作葡萄峪、王窑、青龙峡,奥陶系马家沟三、四、五段。
阿伯尔拟蠕形迹 Helminthopsis abeli Ksiazkiewicz,1977
(图版Ⅱ-g)
描述:阿伯尔拟蠕形迹潜穴多呈凸起状常沿岩层面发育,潜穴管粗细、弯曲不等,整体上呈蛇曲状或具有蛇曲状的趋势,局部较为平直。潜穴直径在0.5~2 cm,长5~15 cm之间。充填物黑色或深灰色,成分与围岩皆为泥晶灰岩,围岩含有生物碎屑。
讨论:前人在建立此遗迹属时认为此潜穴形态没有明显表现出较为规则的蛇曲状,但总体上看具有形成蛇曲状的趋势[20]。Wetzel和 Bromley(1996)通过研究发现该遗迹种常表现为开口不规则的蛇曲形,并认为Helminthopsis可能是一种生存在深度不大的沉积物中的底栖生物的觅食迹,其沉积物中食物比较丰富[20]。
产出层位:焦作青龙峡、葡萄峪,奥陶系马家沟组二段中、三段。
象形拟蠕形迹 Helminthopsis hieroglyphica Heer,1877
(图版Ⅱ-h)
描述:该遗迹种表现出弯曲程度很高的蛇曲形,整体没有分枝,直径均一,为0.8 cm左右。潜穴整体上呈弯曲弧形,局部比较平直,长度10~18 cm。
讨论:该遗迹化石附近有包括腹足类和头足类动物在内的较多实体化石,虽不能证明其为造迹生物,但二者相关性很好,有待进一步研究。Helminthopsis hieroglyphica通常被认为是一种底迹充填潜穴。
产地层位:焦作葡萄峪、青龙峡,马家沟组三、四、五、六段。
线形迹属 Ichnogenus Gordia Emmons,1844
磨砾石线形迹Gordia molassica Heer,1865
(图版Ⅲ-a)
描述:该遗迹种微凸起于层面,呈现出复杂无规则的任意弯曲,局部出现交切现象,并呈多次穿插。潜穴管不分枝,管壁光滑,管径较细,约0.2 cm左右,可呈不规则交叉状态,没有分枝出现,整体宽度一致,长度3~15 cm。充填物与围岩相同,颜色差异因为围岩风化后呈灰黄色。
讨论:Gordia主要特点是互相穿插、交切特征明显,弯曲不规则且频繁[14-15]。研究区该遗迹种特点是潜穴管细长且延伸形态不规则,没有表现出Hel-minthopsis属的蛇曲形或蛇曲趋势,长度不等,较长者常会出现互相穿插现象,较短者穿插不明显,充填物颜色稍深于围岩。
产出层位:焦作葡萄峪、青龙峡,奥陶系马家沟组马家沟组三段,六段下部。
蠕形迹属 Ichnogenus Scolicia de Quatrefages,1849
蠕形迹(未定种)Scolicia ichnosp.
(图版Ⅲ-b)
描述:该遗迹种为弯曲带状遗迹,表面光滑,带宽均匀,约1.6 cm,至末端变细。个体较大,充填物与围岩相同。见不完全保存的衬壁。
讨论:该遗迹属最初由de Quatrefages解释为长的环带动物;而Hantzschel认为是腹足类的潜穴遗迹;Seilacher则认为是由心形海胆形成的潜穴,但是Smith和Crimes发现Scolicia遗迹化石只在白垩纪以后才是心形海胆形成的[14-15]。由此,Scolicia的造迹生物极可能是多种生物[7]。且研究区发现Scolicia和Chondrites常常共生。
产出层位:焦作青龙峡,奥陶系马家沟组六段下部。
4 沉积环境分析
本次发现的遗迹化石多为局限—开阔碳酸盐岩台地中常见的分子,综合研究区遗迹化石的古生态特征和宿主岩层的沉积学特征,将研究区遗迹化石划分为3个遗迹组合。即Psilonichnus-Thalassinoides遗迹组合、Helminthopsis-Gordia遗迹组合和 Chondrites-Scolica遗迹组合。
4.1 Psilonichnus-Thalassinoides遗迹组合及沉积环境
该遗迹组合的岩性组成为中厚层泥质白云岩、灰质白云岩、白云质灰岩或亮晶胶结的泥粒灰岩夹薄层的细晶白云岩,局部地区夹厚5~18 cm的黄褐色角砾状白云岩,砾石大小多为0.3 cm×0.7 cm。该遗迹组合发育层段见鸟眼、干裂构造,局部地区见石盐假晶现象,中厚岩层中水平层理发育。
该遗迹组合主要有居住迹和少量的觅食迹组成。其中居住迹主要由“J”、“Y”潜穴或穿插网状分布的潜穴系统及少量的近水平潜穴组成,遗迹化石分子主要为Psilonichnus、Thalassinoides,遗迹化石的分异度低,丰度中等,局部较高。泥质灰岩或泥晶灰岩中发育少量的进食潜穴Helminthopsis,多与层面平行。个体较大,达17~22 cm,丰度低,分异度高。
该遗迹组合反映了局限碳酸盐岩台地的潮坪环境,局部有潮汐沟道切入,潮道内形成砾石沉积。根据平均高潮线可以划分为潮上坪和潮间坪,潮间坪依据海平面、平均低潮线可以细分为高潮坪、中潮坪和低潮坪。潮上坪一般水动能较弱、环境较稳定,适宜于生物掘穴居住,因此发育大量的Psilonichnus和网络状Thalassinoides潜穴,潮间坪位于平均高潮线和平均低潮线之间,一般水体相对较动荡;但对于局限台地内的潮间坪,水体动荡程度往往又不是太高,氧气充分,有机质丰富,内碎屑灰岩和生物碎屑发育;局限碳酸盐岩台地内的高潮坪类似于潮上坪发育大量Psilonichnus和网络状 Thalassinoides潜穴,中潮坪—低潮坪主要发育相对较粗大的网状Thalassinoides潜穴和Helminthopsis潜穴。
4.2 Helminthopsis-Gordia遗迹组合及沉积环境
该遗迹组合的岩性组成为灰—灰褐色中厚层的白云质灰岩、泥粒—粒泥灰岩、泥晶灰岩夹中层状的灰质白云岩,层厚35~72 cm,生物碎屑相对发育。水平层理发育、局部地区见波痕构造和粒序层理。
该遗迹组合主要有进食迹和觅食迹组成。遗迹化石分子包括:Helminthopsis、Gordia、Planolites、Palaeophycus、Thalassinoides等。遗迹化石多沿层面或斜切层面分布,多呈蛇曲型,常见交叉截切现象。遗迹化石的分异度低—中等,丰度中等,局部较高。野外岩石新鲜断面上遗迹化石 Helminthopsis和 Thalassinoides潜穴充填物相对围岩较浅;而遗迹化石Gordia、planolites、Palaeophycus潜穴充填物和围岩分异不明显。
该遗迹组合反映了低能、静水的沉积环境,局限碳酸盐岩台地的潟湖或开阔碳酸盐岩台地内的滩间海和台内洼地即为这类环境。根据Boucot等[32]显生宙全球古气候重建成果,华北盆地奥陶纪处于干旱气候带上,该区域内局限碳酸盐岩台地中的潟湖的盐度相对较高,只适宜广盐度生物生存,窄盐度生物相对较少,这些生物在生命活动时留下进食迹和觅食迹,如 Helminthopsis、Planolites、Palaeophycus、Thalassinoides等遗迹化石相对实体化石较丰富;对开阔碳酸盐岩台地内的滩间海和台内洼地一般是正常盐度的沉积环境,细粒沉积物中有机质丰富,生物较为丰富,大量的生物在生命活动过程留下丰富多样的遗迹化石,如 Helminthopsis、Gordia、Planolites、Palaeophycus等,因此对比二者而言,开阔碳酸盐岩台地内的滩间海和台内洼地沉积环境中生物实体化石和遗迹化石都较局限碳酸盐岩台地的潟湖丰富。
4.3 Chondrites-Scolicia遗迹组合沉积环境
该遗迹组合的岩性组成主要为灰—深灰中厚层泥晶灰岩,层厚43~87 cm,局部见巨厚层泥晶灰岩,层内纹层不明显,呈块状层理。该遗迹组合主要有进食迹和觅食迹组成。遗迹化石分子包括:Chondrites、Scolicia、Helminthopsis等。其中,进食迹 Chondrites是由未知食沉积动物建造的树枝状高度分支的潜穴系统[33-34]。通常从一个或多个水平或倾斜或垂直管状为其始端向下或向外延展,分支一般2~4级不等,分支角度30°~50°同一个分支潜穴分支角度和分支直径常保持不变,通常从一个或多个水平或倾斜或垂直管状为其始端向下或向外延展[33-34]。而觅食迹Scolicia遗迹化石为沿层面呈水平蛇曲形发育的三叶型遗迹,中叶平,具窄而深的边缘沟[20],该遗迹组合中的Helminthopsis分子形体相对较小,长3~12 cm,宽0.2~0.7 cm。
该遗迹组合反映了低能、静水、贫氧、还原性的沉积环境。在该遗迹组合中Chondrites遗迹化石的指相意义最为明显。在一系列缺氧和富氧事件交替出现的层序地层中,Chondrites总是最后在缺氧和缺生物的纹层沉积之前消失[33-34],而后又会在生物扰动重新有所改善的环境中最先出现,是一个较为典型的机会种。并且研究区Chondrites出现的宿主岩层为深灰—黑色的泥晶灰岩,在溶蚀的Chondrites潜穴内见残余的沥青充填(图版Ⅲ-c,d),这些都可以指示其环境为低能、静水、贫氧、还原性的沉积环境。相对而言Scolicia的指相意义较弱,目前它被认为是海洋深度上的穿相遗迹,虽然在深海沉积环境常常能够见到(特别是深海复理石沉积中),但也有一些见于浅海的Cruziana遗迹相[20]。综合研究区该遗迹组合发育的宿主岩石、生物实体化石等信息,笔者认为该遗迹组合发育于开阔碳酸盐岩台地内台内洼地的氧化还原界面附近。
综合岩石类型、岩石序列组合、沉积构造、生物化石、遗迹化石或遗迹化石组合,参考非常适合华北盆地的塔克(1974)碳酸盐岩沉积模式,建立了研究区奥陶系马家沟组的沉积模式(图3)和垂向沉积环境演化序列(图2)。
5 结论
(1)豫西北奥陶系马家沟组目前共识别出遗迹化石三大生态类型10属17种,即①居住迹(Domichnia):Psilonichnus upsilon、Thalassinoides callianassa、Thalassinoides horizontalis、Rhizocorallium jenense;②进食迹(Foidichnia):Chondrites type-1、Chondrites type-2、Chondrites intricatus、Palaeophycus tubularis、Palaeophycus heberti、Planolites montanus、Planolites annularius、Phycodes palmatus;③ 觅食迹 (Pascichnia):Helminthopsis ichnosp.、Helminthopsis abeli、Helminthopsis hieroglyphica、Gordia molassica、Scolicia ichnosp。
(2)综合研究区鉴定遗迹化石所揭示的古生态和宿主岩层的沉积学特征,将其划分为3种不同沉积条件下的遗迹组合:①Psilonichnus-Thalassinoides遗迹组合代表了局限碳酸盐岩台地的潮坪环境;②Helminthopsis-Gordia遗迹组合反映局限碳酸盐岩台地的潟湖或开阔碳酸盐岩台地内的滩间海和台内洼地等低能、静水的沉积环境环境;③Chondrites-Scolica遗迹组合开阔碳酸盐岩台地内台内洼地的氧化还原界面附近的沉积环境。综合岩石类型、岩石序列组合、沉积构造、生物化石、遗迹化石或遗迹化石组合,建立了研究区奥陶系马家沟组的沉积模式和垂向沉积演化序列。
References)
1 杨式溥.遗迹化石及其对古环境分析的意义[J].沉积学报,1984,2(4):8-18.[Yang Shipu.Trace fossils and their significance for interpretation of the paleoenvironments[J].Acta Sedimentologica Sinica,1984,2(4):8-18.]
2 杨式溥.遗迹化石的古环境和古地理意义[J].古地理学报,1999,1(1):7-19.[Yang Shipu.Palaeoenvironmental and palaeogeographic significance of trace fossils[J].Journal of Palaeogeography,1999,1(1):7-19.]
3 Taylor A,Goldring R,Gowland S.Analysis and application of ichnofabrics[J].Earth-Science Reviews,2003,60(3/4):227-259.
4 Mcllroy D.The Application of Ichnology to Palaeoenvironmental and Stratigraphic Analysis[M].London:Geological Society Special Publications,2004:157-178.
5 Seilacher A.Trace Fossil Analysis[M].Berlin,Heidelberg:Springer-Verlag,2007:75-131.
6 李日辉.内蒙古桌子山地区中奥陶世公乌素组的遗迹化石及遗迹相[J].古生物学报,1993,32(1):89-109.[Li Rihui.Trace fossils and ichnofacies of Middle Ordovician Gongwusu Formation,Zhuozishan,Inner Mongolia[J].Acta Palaeontologica Sinica,1993,32(1):89-109.]
7 李日辉.陕西富平中奥陶统金粟山组的遗迹化石及沉积环境[J].沉积学报,1993,11(2):27-33.[Li Rihui.Trace fossils from Middle Ordovician Jinsushan Formation,Fuping,Shanxi and their depositional environments[J].Acta Sedimentologica Sinica,1993,11(2):27-33.]
8 费安玮.桌子山中奥陶世公乌素组遗迹化石组合与古地理环境[J].现代地质,2000,14(3):366-374.[Fei Anwei.Trace fossil assemblages and palaeoenvironment of Middle Ordovician Gongwusu formation,Zhuozishan,Inner Mongolia[J].Geoscience,2000,14(3):366-374.]
9 方国庆,毛曼君.陕西富平上奥陶统遗迹化石及其环境意义[J].同济大学学报:自然科学版,2007,35(8):1118-1121.[Fang Guoqing,Mao Manjun.Trace fossils and sedimentary environments of upper Ordovician of Fuping in Shaanxi,China[J].Journal of Tongji University:Natural Science,2007,35(8):1118-1121.]
10 叶连俊.华北地台沉积建造[M].北京:科学出版社,1983:64-85.[Ye Lianjun.Sedimentary Associations of the Northern China Platform[M].Beijing:Science Press,1983:64-85.]
11 王英华,张秀莲,杨承运.华北地台早古生代碳酸盐岩岩石学[M].北京:地震出版社,1988:117-133.[Wang Yinghua,Zhang Xiulian,Yang Chengyun.Carbonate Petrology of Early Paleozoic of North China Platform[M].Beijing:Seismological Press,1988:117-133.]
12 冯增昭,王英华,张吉森,等.华北地台早古生代岩相古地理[M].北京:地质出版社,1990:49-74.[Feng Zengzhao,Wang Yinghua,Zhang Jisen,et al.Lithofacies Paleogeography of Early Paleozoic of North China Platform[M].Beijing:Geology Press,1990:49-74.]
13 刘印环,王建平,张海清,等.河南省寒武系和奥陶系[M].北京:地质出版社,1991:87-164.[Liu Yinhuan,Wang Jianping,Zhang Haiqing,et al.Cambrian and Ordovician of Henan Province[M].Beijing:Geological Publishing House,1991:87-164.]
14 胡斌,张文涛,牛永斌.焦作云台山与青龙峡景区中奥陶统马家沟组沉积特征及演化规律[J].河南理工大学学报:自然科学版,2011,30(4):411-417.[Hu Bin,Zhang Wentao,Niu Yongbin.Sedimentary characters and evolution of the Middle Ordovician Majiagou Formation in the Black Dragon Canyon,Yuntai Mountain,Jiaozuo area[J].Journal of Henan Polytechnic University:Natural Science,2011,30(4):411-417.]
15 张文涛.云台山世界地质公园奥陶系马家沟组碳酸盐岩微相及遗迹群落研究[D].焦作:河南理工大学,2011:51-59.[Zhang Wentao.Carbonate rock microfacies and Ichnocoenosis in the Ordovician Majiagou Formation,Yuntai world geopark[D].Jiaozuo:Henan Polytechnic University,2011:51-59.]
16 牛永斌,董小波,朱信生,等.豫西北太行山奥陶系露头区裂缝发育特征及主控因素[J].现代地质,2013,27(4):809-818.[Niu Yongbin,Dong Xiaobo,Zhu Xinsheng,et al.Fracture characteristics and main controlling factors of Ordovician outcrop zones of Taihang Mountains in the north-west of Henan province[J].Geoscience,2013,27(4):809-818.]
17 席文祥,裴放.河南省岩石地层[M].武汉:地质大学出版社,1997:81-115.[Xi Wenxiang,Pei Fang.Stratigraphy(Lithostratic)of Henan Province[M].Wuhan:China University of Geosciences Press,1997:81-115.]
18 张国成,王昆.四川峨眉山龙门洞下三叠统嘉陵江组一段遗迹组构及其沉积学意义[J].古地理学报,2010,12(3):281-290.[Zhang Guocheng,Wang Kun.Ichnofabrics and their sedimentary significance of the member 1 of Lower Triassic Jialingjiang Formation at Longmendong Emei Mountains,Sichuan province[J].Journal of Palaeogeography,2010,12(3):281-290.]
19 胡斌,王冠中,齐永安.痕迹学理论与应用[M].徐州:中国矿业大学出版社,1997:10-41,126-151.[Hu Bin,Wang Guanzhong,Qi Yongan.The Theory and Application of Ichnology[M].Xuzhou:China University of Mining Technology Press,1997:10-41,126-151.]
20 杨式溥,张建平,杨美芳.中国遗迹化石[M].北京:科学出版社,2004:29-263.[Yang Shipu,Zhang Jianping,Yang Meifang.Trace Fossils in China[M].Beijing:Science Press,2004:29-263.]
21 Frey R W,Howard J D.Trace fossils from the Panther Member,Star point Formation(Upper Cretaceous),Coal Creek Canyon,Utah[J].Journal of Paleontology,1985,59(2):370-404.
22 Seilacher A.Fossil behavior[J].Scientific American,1967,217(2):72-80.
23 Basan P B,Scott R W.Morphology of Rhizocorallium and associated traces from the Lower Cretaceous Purgatoire Formation,Colorado[J].Palaeogeography,Palaeoclimatology,Palaeoecology,1979,28:5-23.
24 晋慧娟,李育慈.古代深海遗迹化石群落在沉积学中的应用[J].科学通报,1999,44(2):123-130.[Jin Huijuan,Li Yuci.Application of the ancient deep trace fossil group in deep sea to the sedimentology[J].Chinese Science Bulletin,1999,44(2):123-130.]
25 晋慧娟.古代深海沉积体系遗迹化石研究[J].矿物岩石地球化学通报,1999,18(1):42-45.[Jin Huijuan.Research on trace fossils of ancient deep-water sedimentary system[J].Bulletin of Mineralogy Petrology and Geochemistry,1999,18(1):42-45.]
26 周志澄.生物成因的构造在环境解释中的应用——遗迹学研究的新进展[J].古生物学报,1995,34(2):228-249.[Zhou Zhicheng.Application of biogenic structures to environmental interpretation-advancement in studies of Ichnology[J].Acta Palaeontologica Sinica,1995,34(2):228-249.]
27 龚一鸣.遗迹化石Chondrites的指相意义和阶层分布[J].古生物学报,2004,43(1):94-102.[Gong Yiming.Facies characteristics and tiering distributions of Chondrites[J].Acta Palaeontologica Sinica,2004,43(1):94-102.]
28 赵小明,段其发,牛志军,等.青海南部曲麻莱-治多地区三叠纪遗迹化石的地层学意义[J].地层学杂志,2007,31(4):333-346.[Zhao Xiaoming,Duan Qifa,Niu Zhijun,et al.The stratigraphic significance of the Triassic trace fossils in the Qumarleb-Zhidoiarea,Southern Qinghai[J].Journal of Stratigraphy,2007,31(4):333-346.]
29 赵小明,牛志军,段其发,等.青海曲麻莱-治多地区巴颜喀拉山群(上三叠统)遗迹化石[J].古生物学报,2007,46(1):122-134.[Zhao Xiaoming,Niu Zhijun,Duan Qifa,et al.Trace fossils of the Upper Triassic Bayanharshan Group in the Qumarleb-Zhidoiarea,Southern Qinghai[J].Acta Palaeontologica Sinica,2007,46(1):122-134.]
30 晋慧娟,李育慈,方国庆.古代深海底质氧控的遗迹化石群落[J].沉积学报,2003,21(1):75-80.[Jin Huijuan,Li Yuci,Fang Guoqing.Oxygen-dependent ichnocoenose in paleo-pelagic substrates[J].Acta Sedimentologica Sinica,2003,21(1):75-80.]
31 Bromley R G,Ekdale A A.Trace fossil preservation in flint in the European chalk[J].Journal of Paleontology,1984,58(2):298-311.
32 Boucot A J,陈旭,Scotese C R,等.显生宙全球古气候重建[M].北京:科学出版社,2009:34-39.[Boucot A J,Chen Xu,Scotese C R,et al.Reconstruction of Phanerozoic Global Paleoclimate[M].Beijing:Science Press,2009:34-39.]
33 牛永斌,钟建华,钟福平,等.柴达木盆地南缘石炭系Chondrites遗迹化石的发现及其地质意义[J].古地理学报,2008,10(5):529-535.[Niu Yongbin,Zhong Jianhua,Zhong Fuping,et al.Discovery of trace fossil Chondrites in the Carboniferous in the south margin of Qaidam basin and its geological significance[J].Journal of Palaeogeography,2008,10(5):529-535.]
34 牛永斌,钟建华,胡斌.小尺度地质体三维建模研究——以遗迹化石Chondrites和岩心三维建模为例[J].古地理学报,2008,10(2):207-214.[Niu Yongbin,Zhong Jianhua,Hu Bin.Research of 3D modeling on small-scale geologic body—Taking 3D modeling on trace fossil Chondrites and drilling core as an example[J].Journal of Palaeogeography,2008,10(2):207-214.]
猜你喜欢
华人时刊(2021年17期)2021-12-02
西南石油大学学报(自然科学版)(2021年3期)2021-07-16
——哈尔滨市马家沟小学教育剪影
黑龙江教育(教育与教学)(2021年12期)2021-03-22
矿产勘查(2020年11期)2020-12-25
矿产勘查(2020年7期)2020-12-25
西南石油大学学报(自然科学版)(2019年5期)2019-12-20
西南石油大学学报(自然科学版)(2018年5期)2018-11-06
紫禁城(2017年6期)2017-08-07
散文诗(2017年22期)2017-06-09
现代企业(2015年7期)2015-02-28
