
镉胁迫下泡菜乳酸乳球菌的形态变化
2015-12-07 02:58连元元谷新晰黄昆仑田洪涛
食品科学 2015年1期
连元元,王 颖,李 晨,谷新晰,盛 耀,黄昆仑,*,田洪涛,*
(1.河北农业大学食品科技学院,河北 保定 071001;2.中国农业大学食品科学与营养工程学院,北京 1000 83)
镉胁迫下泡菜乳酸乳球菌的形态变化
连元元1,王 颖2,李 晨1,谷新晰1,盛 耀2,黄昆仑2,*,田洪涛1,*
(1.河北农业大学食品科技学院,河北 保定 071001;2.中国农业大学食品科学与营养工程学院,北京 1000 83)
通过扫描电镜和透射电镜分别观察不同质量浓 度水平的Cd2+对泡菜乳酸乳球菌(Lactococcus lactis subsp. lactis)细胞的影响,扫描电镜结果显示:Cd2+质量浓度在0、10 mg/L时,泡菜乳酸乳球菌呈椭圆形、表面光滑、菌体生长繁殖旺盛,随着Cd2+质量浓度的增加菌体细胞表面产生白色颗粒状物质、菌体细胞存活数量大幅下降(OD600nm值由1.336下降到0.515)。当添加200 mg/L Cd2+时,几乎没有见到明显的菌体、显示有少量棱形晶状物。透射电镜结果显示:当Cd2+质量浓度为0~50 mg/L时泡菜乳酸乳球菌结构完整、细胞内容物分布均、菌体生长较为正常,当菌体暴露于100、200 mg/L Cd2+时菌体细胞出现异常现象,如细胞破裂、内容物从薄膜穿孔中释放、质壁分离等。两类电镜结果均表明:在低质量浓度Cd2+(≤50 mg/L)胁迫下,对泡菜乳酸乳球菌的生长几乎不产生影响,添加Cd2+质量浓度上升到100、200 mg/L时泡菜乳酸乳球菌正常生长受到抑制。
乳酸乳球菌;镉胁迫;电镜;亚细胞结构
镉是一种过渡金属,是对生物有机体最具威胁的重金属元素之一。目前我国粮食作物镉污染情况大致如下:1980年中国农业环境报告,我国农田土壤镉污染面积为9 333 hm2,随后有人提出镉污染面积为13 333 hm2(根据GB 15618—1995《土壤环境质量标准》土壤中镉的限量标准是0.006 mol/kg);最近据不完全统计,我国受镉污染的农田面积达到2.8×105hm2,每年生产的镉含量超标的农产品达1.46×1010kg(我国GB 2762—2012《食品中污染物限量》规定大米中镉限量标准是0.2 mg/kg),使得镉污染严重威胁着农业生态环境、人类健康安全。生物吸附法是最近发展较快较先进的去除重金属方法,生物吸附法与传统方法相比具有如下优点:原料来源丰富,品种多;设备简单,运行费用低;吸附量大,处理效率高; pH值和温度条件范围宽。
目前镉暴露产生了严重的健康危害,而且镉在膳食来源中的暴露不断增加,但没有可行的方法去除食品和饲料中的镉。对于食品重金属的去除,除了酿酒酵母属于食品微生物外,其他的食品微生物,例如食品益生菌研究甚少。而最近食品中公认的益生菌乳酸菌越来越受到关注,关于利用其去除食物中污染的重金属离子在国内处于初步研究,但也显示出了可行性和潜力[1]。
实验选取镉作为重金属靶元素。实验中的菌株是自行由泡菜中提取并保存的一株泡菜乳酸乳球菌。经实验发现,其最高耐镉质量浓度可达200 mg/L,由于其是食品中的益生菌(根据进化树比较发现其同源性与Lactococcus lactis subsp. cremoris strain IMAU40012同源性上最近,而后者为中国益生菌网中可查到的一株益生菌),因此可用于去除食品中的镉。有研究表明镉对食用菌生长的影响不仅体现在生长速率上,还体现在不同程度地影响细胞膜结构和细胞器[2]。扫描电镜和透射电镜可以用来清晰地观察微生物细胞表面和内部的结构以及细胞器。本实验利用了这两种电镜方法探索讨论了镉添加后泡菜乳酸乳球菌菌体表面和亚细胞结构的变化,可以直观地观察到重金属在其细胞表面和亚细胞器上的分布和毒性作用,从而推测重金属的吸收途径和可能参与的机体代谢,以便后续总结出其与镉之间的相互作用机制,揭示其耐镉分子机制,最后应用于食品中镉污染的治理。同时为衡量此株泡菜乳酸乳球菌是否适应重金属环境以及重金属耐受性大小提供依据。
1 材料与方法
1.1 菌株
来源:分离自含Cd2+的泡菜,保存在中国农业大学东校区金工楼。
1.2 试剂
2.5 %戊二醛(医用)、25%戊二醛(医用)溶液武汉市帝科化工有限公司;锇酸 北京精华耀邦医药科技有限公司;磷酸缓冲液(0.1 mol/L,pH 7.2) 北京奥博来科技有限责任公司;0.2 mol/L磷酸氢二钠贮备液、乙醇 北京化工厂;0.2 mol/L磷酸二氢钠贮备液北京鹏彩精细化工有限公司;环氧丙烷 东营汇中化工有限公司;SPI812环氧树脂 北京中兴百瑞技术有限公司;以上试剂均为分析纯。
1.3 仪器与设备
TM3000扫描电镜、JEM-1230透射电镜、JSM-6301F扫描电镜、JEOL-2010F高分辨率透射电镜 日本 Hitachi公司。
1.4 方法
将单菌落挑到新鲜的液体MRS培养基中进行活化,以1%接种量转接到新鲜的MRS培养基中,在37 ℃条件下预培养6 h,添加Cd2+至终质量浓度10、50、100、200 mg/L,20 h后收集菌液,去离子水清洗3 遍,制备电镜样品。
1.4.1 扫描电镜样品前处理
取各实验组菌体(未添加Cd2+和Cd2+添加培养菌体),蒸馏水洗涤5 次,以祛除菌体表面浮色,5%戊二醛固定,1%锇酸再固定,乙醇梯度(30%、50%、70%、80%、90%、100%)脱水,乙酸异戊酯处理后,临界点干燥喷金,实验采用TM3000型扫描电镜观察菌体表面特征变化。
1.4.2 透射电镜样品前处理
取各实验组菌体(未添加Cd2+和Cd2+添加培养菌体),清洗后立即放入福尔马林的溶液中浸泡保存,环氧树脂包埋,超薄切片后,采用JEM-1230型透射电镜观察富集前后菌体内部结构变化。
2 结果与分析
2.1 扫描电镜结果
当培养基中Cd2+达到50 mg/L时菌体依旧大量存活,菌体表面附着有白色细小盐状晶体。图1a显示为分辨率1 μm时的菌体形态,有白色颗粒状晶体以团状存在,包裹在菌体细胞外。当培养基中添加的Cd2+达到100 mg/L时,菌体量较少,出现白色颗粒状盐沉淀。图1b(分辨率为1 μm)中更明显的显示出白色凝集状盐结晶,可以观察到背景中正分裂的菌体和一较长菌体,猜测可能与形成的细菌生物膜来黏附重金属以适应环境压力有关[3]。Aymerich[4]、Behr[5]和Kubota[6]等也发现乳酸菌在逆境环境中可以形成生物质。图1c显示为Cd2+200 mg/L处理时通过扫描电镜观察到的泡菜乳酸乳球菌形态,几乎没有见到明显的菌体,显示有少量棱形晶状物,与形成结晶生物化石可能存在关联。重金属在微生物表面通过与螯合物形成沉淀或在细胞壁上与特殊结构如S-蛋白形成核中心,可促进生物矿石晶体生长。而且这种沉淀和生物矿石有时有重叠的现象,尚不清楚是哪个作用单独或主要起到了固定重金属的作用[7]。
2.2 透射电镜结果
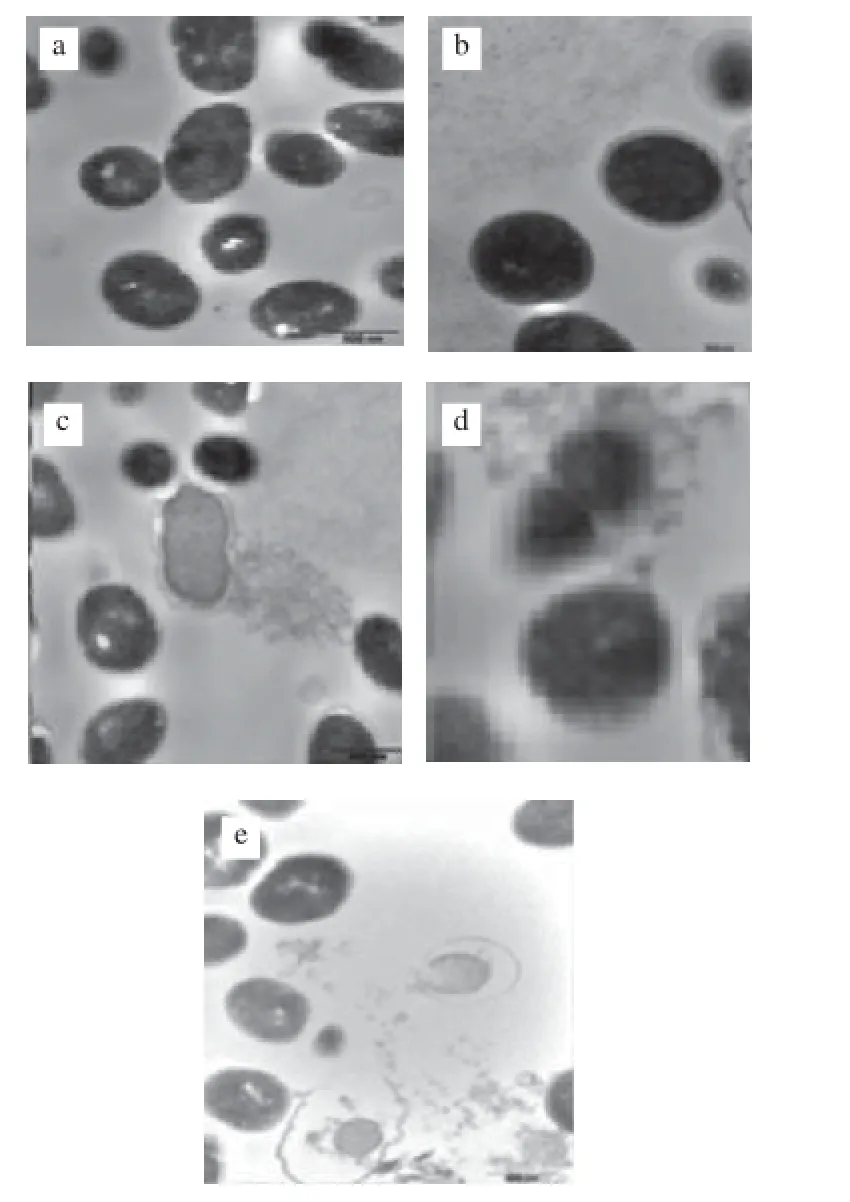
图2 添加Cd2+泡菜乳酸乳球菌透射电镜图Fig.2 Scanning electron micrographs of Lactococcus lactis subsp. lactis exposed to different concentrations of Cd2+
图2a所示为未添加Cd2+,处于分裂期有完整细胞的泡菜乳酸乳球菌(透明部分为切片不当所致,与菌体形态无关)。图2 b、2c所示为Cd2+在质量浓度10、50 mg/L时透射电镜图,菌体受Cd2+影响不大,多数为正常生长和处于分裂期的菌体细胞,细胞完整饱满,内容物分布均匀致密,细胞壁薄。Cd2+质量浓度50 mg/L时有少量质壁分离现象,质壁分离处有气泡或囊泡,可能和菌体自身隔离Cd2+采取的防御措施有关,是细胞内阻止胞质积累的重金属对重要细胞器毒害的有效策略[8-10],这在真核微生物如藻类、真菌、酵母中经常出现。真菌积累的大部分Co2+、Mn2+通常情况下存在于小气泡中,并以离子的形式存在或结合于多聚磷酸盐上或通过蛋白质发挥贮存、调节金属离子浓度的作用,例如聚球菌属(Synechococcus sp.)产生的金属硫蛋白[11]。当Cd2+、Zn2+浓度较高时,聚球菌属(Synechococcus sp.)的抗Cd、Zn基因smtA、smtB诱导表达,并且受smtB产物的抑制,smtA金属硫蛋白半胱氨酸残基可与过量的金属阳离子产生沉淀[12-13]。
图2d、2e是Cd2+质量浓度添加至100、200 mg/L时乳酸菌微观透射显微镜图,显示有些菌还是正常现象。有些菌已经开始有异常或凋敝的现象。以上图片可以呈现出乳酸菌体异常凋敝的动态过程。未添加Cd2+和低质量浓度Cd2+组别菌体横切面为规则圆形,细胞壁规则致密,细胞膜完整,菌体饱满,表面无黑色颗粒物,当Cd2+质量浓度高至100 mg/L时,菌体细胞壁质地疏松,厚度增大,出现气泡、囊泡,表明菌体对Cd2+具有较高抗性;Cd2+质量浓度继续升高至200 mg/L时,出现质壁分离,菌体细胞膜有明显穿孔,有内容物释放现象,猜测为胞内染色质和蛋白结构物。非必需金属元素可取代结合位点的必需元素,与配基反应,产生毒害作用,导致核酸、蛋白质构象改变,干扰氧化磷酸化及渗透压的平衡,从而导致菌体代谢失常和凋敝[14-15]。樊霆[16]在研究真菌CTB430对Cu和Zn抗性时比较菌体富集Cu和Zn前后菌体内部结构变化时也有类似损伤菌体真丝及细胞壁膜现象出现。
Cd2+质量浓度较低(50 mg/L以下)时菌体正常生长,质量浓度逐渐升高时有黑色电子密度颗粒、液泡等一些异常现象出现,体现了菌体自身防御机制在抗Cd2+毒性方面的作用,Cd2+质量浓度较高(200 mg/L)时出现质壁分离,细胞膜出现穿孔,内容物外泄等结果推测泡菜乳酸乳球菌可能具有通过气泡或囊泡隔离Cd2+的作用。金属离子进入细胞后,通过区域化作用(c ompartmentalization)分布在细胞内的不同部位,可将有毒金属离子封闭(如进入液泡或线粒体)或与热稳定蛋白结合,转变成为低毒的形式,是更有效的解毒方式[17]。
3 结 论
扫描电子显微镜照片显示,较低Cd2+质量浓度(10、50 mg/L)处理时,泡菜乳酸菌细胞长短径及细胞分裂,表面完整性与对照组比对都没有出现明显改变,这说明较低Cd2+质量浓度对于菌体影响不大;加大Cd2+质量浓度(100、200 mg/L)时,存活的菌体细胞量很少,有大量白色颗粒晶体盐以及菱形状岩石出现,这可能是镉的各种沉淀形式(后续实验通过扫描电镜与X射线谱图联用,发现Cd2+质量浓度100、200 mg/L时晶状体的主要元素是分别是C、O、Na、Cd和C、O、Cd)。
透射电子显微镜扫描结果显示,未添加Cd2+菌和较低Cd2+质量浓度(10、50 mg/L)时泡菜乳酸菌细胞完整度较好,内容物分布较充实;当Cd2+质量浓度升至100、200 mg/L时菌体细胞破裂穿孔较普遍,内容物大量释放在胞外环境中,内部有黑色电子密度颗粒堆积,质壁分离,细胞周质出现含有黑色电子密度颗粒的气泡或囊泡,推测是Cd的络合物。
低质量浓度Cd2+(10、50 mg/L)条件下,泡菜乳酸菌能较正常生长代谢,高质量浓度Cd2+(100、200 mg/L)时,泡菜乳酸乳球菌受Cd2+影响较大出现不正常的衰败,死亡。在泡菜乳酸菌细胞表面有镉沉淀,内部有镉的络合物,这与菌体自身的耐镉机制有关。具体的耐镉机制后续会进一步研究,可通过双向电泳寻找差异蛋白,之后进行质谱鉴定,根据鉴定结果进行实时荧光定量聚合酶链式反应实验,确定关键耐镉基因,即最终确定耐镉的分子机制。
[1] 江南大学. 一种具有排镉功能的植物乳杆菌及其用途: 中国, CN102827796 B[P/OL]. 2012-12-19[2013-10-30]. http://www.google. co.uk/patents/CN102827796B?cl=zh&hl=zh-CN.
[2] 黄敏敏, 江枝和, 翁伯琦, 等. 镉对姬松茸菌丝体细胞超微结构的影响[J]. 热带作物学报, 2011, 32(6): 1082-1085.
[3] DONLAN R M. Biofilms: microbial life on surfaces[J]. Emerging Infectious Diseases, 2002, 8(9): 88l-890.
[4] AYMERICH T, MARTÍN B, GARRIGA M, et al. Microbial quality and direct PCR identifi cation of lactic acid bacteria and nonpathogenic staphylococci fromartisanal low-acid sausages[J]. Applied and Environmental Microbiology, 2003, 69: 4583-4594.
[5] BEHR J, GÄNZLE G M G, VOGEL R F. Characterization of a highly hop-resistant Lactobacillus brevis strain lacking hop transport[J]. Applied and Environmental Microbiology, 2006, 72: 648-649.
[6] KUBOTA H, SENDA S, NOMURA N, et al. Biofilm formationby lactic acid bacteria and resistance to environmental stress[J]. Journal of Bioscience and Bioengineering, 2008, 106: 381-386.
[7] GLASAUER S, LANGLEY S, BEVERIDGE T J. Sorption of Fe(hydr)oxides to the surface of Shewanella putrefaciens: cell-bound fi ne-grained minerals are not a lways formed de novo[J]. Applied and Environmental Microbiology, 2001, 67: 5544-5550.
[8] COBBETT C, GOLDSBROUGH P. Phytochelatins and metallothionei roles in heavy meta l detoxifi cation and homeostasis[J]. Annual Review of Plant Biology, 2002, 53: 159-182.
[9] CLEMENS S. Evolution and function of phytochelatin synthases[J]. Journal of Plant Physiology, 2006, 163(3): 319-332.
[10] CAI Yong, SU Jinhui, LENA Q Ma, Low molecular weight thiols in arsenic hyperaccumulator Pteris vittata upon exposure to arsenic and other trace elements[J]. Environmental Pollution, 2004, 129: 69-78.
[11] MARTIN S, RENE K, VACEK J, et al. Electrochemical study of heavymetals and metallothionein in yeast Yarrowia lipolytica[J]. Bioelectrochemistry, 2003, 60: 29-36.
[12] 徐柳, 宋琴, 茆灿泉. 金属结合蛋白(肽)与环境重金属生物修复[J].生物工程杂志, 2004, 24(4): 39-43.
[13] COOK W J, KAR S R, TAYLOR K B, et al. Crystal structure of the cyanobacterial metallothionein repressor SmtB: a model for metalloregulatory proteins[J]. Journal of Plant Physiology, 1998, 275: 337-346.
[14] 滕应. 重金属污染下红壤微生物生态特征及生物学指标研究[D]. 杭州: 浙江大学, 2003.
[15] BRUINS M R, KAPIL S, OEHME F W. Microbial resistance to metals in the environment[J]. Ecotoxicology and Environmental Safety, 2000, 45(3): 198-207.
[16] 樊霆. 真菌对重金属的抗性机制及富集特性研究[D]. 长沙: 湖南大学, 2009.
[17] 王保军, 杨惠芳. 微生物与重金属的相互作用[J]. 重庆环境科学, 1996, 18(1): 35-39.
Morphological Change of Lactococcus lactis subsp. lactis under Cadmium Stress
LIAN Yuanyuan1, WANG Ying2, LI Chen1, GU Xinxi1, SHENG Yao2, HUANG Kunlun2,*, TIAN Hongtao1,*
(1. College of Food Science and Technology, Hebei Agricultural University, Baoding 071001, China; 2. College of Food Science and Nutritional Engineering, China Agricultural University, Beijing 100083, China)
Using scanning electron microscopy (SEM) and transmission electron microscopy (TEM), the effects of different levels of cadmium (Cd2+) on Lactococcus lactis subsp. lactis cells were examined. The SEM results showed that when the mass concentration of Cd2+was 0 and 10 mg/L, the bacterium displayed an oblong shape with smooth surface and grew vigorously. With increasing concentration of Cd2+, white granules appeared on the surface and the number of surviving cells began to fall (i.e., OD was reduced from 1.336 to 0.515). Under the stress of Cd2+at 200 mg/L, almost no visible bacteria were seen and a small amount of prismatic crystals app eared. The TEM results revealed that when the mass concentration of Cd2+was 0–50 mg/L, the cell structure of this strain was intact and the cellular contents were evenly distributed, and the strain grew normally. When exposed to 100 and 200 mg/L Cd2+, it began to change abnormally such as ruptured cells and release of cellular contents f rom holes in the cellular membrane. In addition, plasmolysis was observed. Together, these results indicated that Lactococcus lactis subsp. lactis was hardly affected by Cd2+at low concentrations (≤ 50 mg/L) but inhibited at high concentrations (100 and 200 mg/L).
Lactococcus lactis subsp. lactis; cadmium stress; electron microscope; subcellular structure
Q934.3
A
1002-6630(2015)01-0124-04
10.7506/spkx1002-6630-201501024
2014-01-26
国家自然科学基金主任基金项目(31140065);河北省自然科学基金项目(C2012204099)
连元元(1986—),女,硕士研究生,研究方向为益生菌资源的开发与应用。E-mail:544408643@qq.com
*通信作者:黄昆仑(1968—),男,教授,博士,研究方向为转基因产品食用安全评价与检测、食品安全风险评估技术与检测技术、食品生物技术。E-mail:hkl009@163.com
田洪涛(1963—),男,教授,博士,研究方向为益生菌资源的开发与应用,工业微生物技术。E-mail:tht631022@163.com
猜你喜欢
实验室研究与探索(2021年11期)2021-01-28
山东化工(2020年15期)2020-02-16
数学大王·中高年级(2019年12期)2019-12-24
实验技术与管理(2019年6期)2019-07-12
智富时代(2018年7期)2018-09-03
智富时代(2018年7期)2018-09-03
教育教学论坛(2017年40期)2017-10-26
恋爱婚姻家庭(2016年18期)2016-07-23
岩矿测试(2015年3期)2015-12-21
岩矿测试(2015年3期)2015-12-21
