
光照和辐射条件与林业生产的关系
2015-12-02 04:17骆宗诗赵顺才刘小云
四川林业科技 2015年3期
骆宗诗,赵顺才,刘小云
(1.四川省林业科学研究院,四川成都 610081;2.四川省富顺县林业局,四川富顺 643000)
1 引言
太阳辐射是地球表面热量的来源,是热量平衡和水分循环的重要决定因素,也是林木生命活动过程中的主要影响因子和能量来源[1,2,3,4]。林业生产的实质是利用植物的光合作用,而光合作用的本质是植物把光能转化成化学能。因此,可以说“现代林业生产实质上是经营光合作用的企业”[1]。林业产量不管是经济产量还是生物产量的高低取决于植物对光能的转化和利用,经济产量的上限,最终还是受光能利用效率的限制。太阳辐射对林木的影响依其强度、性质(或波长)和持续时间,以及光暗交替周期长短而定。林业生产和经营可以通过各种育林技术措施最大限度地利用太阳辐射的光量和能量,提高林业生产力[1,2,11~13,17,18]。
2 光照和辐射条件与林业生产的关系
光照和辐射对林业生产的影响主要表现在光谱(光质)、光时和光强3个方面。
2.1 光谱(光质)与林业生产
太阳辐射主要由紫外线、可见光和红外线3部分组成。由于大气对太阳辐射的吸收和散射具有选择性,所以当太阳辐射通过大气时,不仅辐射强度减弱了,而且光谱组成也发生了变化[2,15,22]。到达地面的太阳辐射随太阳高度角的增大,紫外线和可见光波所占的比例增加,红外线所占的比例相应减少;太阳高度角变小,则长波光的比例增加。光谱成分(光质)在空间上的变化是低纬度处短波多,随纬度增高长波光增多;随海拔的升高,短波光的量也逐渐增加。在季节变化上,夏季短波光较多,冬季长波光增多;一天之内,中午短波光较多,旱晚则长波光增多。不同的光谱成分对植物有不同的作用,植物叶片对太阳辐射的吸收、反射和透射的程度,也直接与辐射的波长有关。
2.1.1 紫外线对植物的生态作用
到达地面的紫外线波长在290 nm~400 nm间,能量仅5%。紫外线的长波部分对植物有刺激作用,可促进种子发芽,这已用于苗木种子催芽。Arehur等[2]通过实验认为紫外线对果实着色效果最好,可见光只有微弱的作用,红外线则几乎无效[1,2]。山地因紫外线多,温差大,不仅有利于果实着色、成熟,而且一般蛋白质含量高。此外,紫外线的短波部分对植物有强烈的抑制生长作用,甚至破坏原生质。紫外线能引起向光性的敏感和促进花青素的形成。因此,高山植物一般都具有茎干短矮、叶面缩小、毛茸发达、叶绿素增加、茎叶富含花青素,花色鲜艳等特征。
2.1.2 红外线对植物的生态作用
波长大于760 nm的近红外线,叶吸收很少,大部分被反射和透射,而对远红外线吸收较多。叶对红外线的反射,阔叶树比针叶树更明显,这是利用红外感光片进行航空摄影和遥感技术以区别针阔叶林的原理[3]。波长更长的红外线,可用热遥感器探知,从而快速准确地发现和预报森林火灾。感知更长波段的遥感器,可探病虫害,因为感染病虫害的树木要比健康者高几度。红外线的生态作用是促进植物茎的延长生长,有利于种子和孢子萌发,提高植物体的温度。
2.1.3 可见光区对植物的生态作用
可见光波段的波长在380 nm~760 nm之间,又可分为红光(760 nm~626 nm),橙光(626 nm~595 nm),黄光(595 nm~575 nm),绿光(575 nm~490 nm),蓝光(490 nm~435 nm),紫光(435 nm~380 nm)七色光。太阳辐射的连续光谱中,植物光合作用利用和色素吸收,具有生理活性的波段称生理有效辐射或光合有效辐射,约在380 nm~740 nm间,这与可见光波段基本相符,对植物有重要意义,因此生态学上往往着重研究可见光与植物之间的关系[1~4]。生理辐射约占太阳总辐射中的 40%~50%。生理有效辐射中,红、橙光是被叶绿素吸收最多的部分,具有最大的光合活性,红光还能促进叶绿素的形成。蓝、紫光也能被叶绿素、类胡萝卜素所吸收。光合作用很少利用绿光,这是因为叶透射和反射的结果,不同波长的光对光合产物成分也有影响。实验表明,红光有利于碳水化合物的合成,蓝光有利于蛋白质的合成。在诱导植物形态建成、向光性和色素形成等方面,不同波长的光合作用有异。蓝紫光与青光对植物伸长生长及幼芽形成有很大作用,能抑制植物的伸长而使其矮态,青蓝紫光还能引起植物向光性的敏感,并能促进花青素等植物色素的形成。红光影响植物开花、茎的伸长和种子的萌发。红外线和红光是地表热量的基本来源,它们对于植物的影响主要是间接地以热效应反应出来。
2.2 光时与林业生产
光照时间的长短作为生态因子,对林业生产的影响是十分重要的。光照持续时间的变化,不仅影响植物的地理分布,而且对于植物生长的营养期和繁殖期均有影响。对于林木,日照长短可影响其生长、花芽分化、开花结实、地上部分发育、休眠和抗寒性等。
2.2.1 光照长度对开花的生态作用
根据植物开花所需的日照长短,可区分为长日照植物(通常需要14 h以上的光照才能开花)、短日照植物(一般需要14 h以上的黑暗才能开花)和中日照植物(花芽形成需要中等日照时间)。植物开花与光敏色素有关[2]。在光周期诱导下,通过光敏色素系统的转化,导致叶内形成某种开花激素。光敏色素在植物体内有两种状态,一种是吸收红光的P660,一种是吸收远红光的P730,它们之间的关系为:
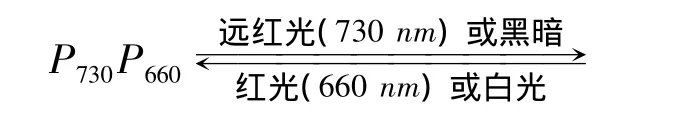
研究表明,植物开花需要一定的P730与P660比值。短日照植物要求较低的P730/P660比值,当日长时,P730增多,就使短日照植物开花受到抑制;当秋季到来夜渐长时,P730减少,就开始花的形成。由光敏色素参加的开花反应,使植物在一年中的适当季节开花。在漫长的秋夜中,如果用红光照射植物,光敏色素P660将很快地转变为P730,短日照植物的开花就受到阻碍。利用这个原理,用闪光打破长夜,就可以阻止短日照植物的开花,使菊花和一品红等的开花延迟几个月。
木本植物开花结实不仅受光周期控制,而且还有其它影响因素,如营养积累和光照强度等。目前,对树木光周期反应的研究还不够深入,大部分实验是用实生苗或插条完成,光周期如何控制成年树木繁殖还了解不多。许多树种的开花似乎属于日中性型(完成开花和其它生命史阶段与日照长短无关),树木在黑暗或连续光照下照样打破休眠,表明休眠的结束不是光调节现象,温度常是光周期反应的重要补充因子[3,16,14,19~21]。
2.2.2 日照长度对叶片生长的生态作用
叶片生长除受水肥条件外,还与光照强度、温度高低十分密切。据 Lenn(1969)报道[1,18],华盛顿脐橙在昼/夜温度为24℃~19℃的条件下,以光照时间长度为8 h的叶面积为100%,则12 h的为107.8%,16 h的为170.2%;在30℃~25℃的高温条件下,叶面积也随光照时间的增长而增大,相应为100%,120.8%,127.4%。在低温条件下叶片数量随日照时间的增长而增多,但高温条件下则不明显。
2.2.3 日照长度对高粗生长的生态作用
许多研究者指出,短日照对树木新梢和幼苗生长产生抑制作用,而长日照则产生促进作用。这可能因为在长光照条件下,成熟的叶片里产生一种刺激形成层活动的植物生长激素类的物质,一些落叶树种在长日照下可以延长落叶期。短日照因抑制枝条生长和促进落叶,形成层活性也随之而受影响,所以短日照抑制树木茎粗生长[1]。
2.2.4 日照长度与抗性和休眠
生长在温带和寒带植物,在冬季表现出一种抗逆性的适应性休眠。一般情况下短日照条件刺激幼叶产生一种刺激物质,传递到芽中抑制芽的生长,可诱发休眠。如,桃树顶芽在长日照(16 h)及17℃~23℃条件下不进行休眠,如果放在13 h日照条件下一个时期即可进入休眠,叶片很快衰老脱落[2,6]。因长光照影响而引起生长延长到生长季晚期的木本植物,常因抗寒锻炼不足,比生长在正常下的易于受冻。一些热带经济林树种,在短日照下可连续生长,不一定诱发休眠。越冬的抗寒性决定于越冬期的糖类物质的积累和木栓化的程度[1,6]。
2.2.5 日照长度的生态类型对树木引种的意义
光周期现象已在各种类型植物和不同纬度地区发现,但以气候易变且季节明显的极地、高纬度和高山地区最明显。光周期是控制野生植物地理分布、也是影响树种栽培范围的重要因素。大多数短日照植物原产热带,其原产地生长季日照时间短;长日照植物原产温带和寒带,夏季生长发育旺盛,一天受光时间长。经济价值高的针叶树种多是长日照植物。北树南移,树木会出现两种情况,一是枝条提前封顶,缩短生长期、生长缓慢、抗逆性差,容易被淘汰;另一种情况是出现二次生长,延长生长期。把短日照植物向北移,其高生长量经常比原产地大,因为日照时数较长和诱导休眠所需的短日照直到晚夏或秋季才出现[3,6]。因休眠的延后,可能在初霜前尚未进及休眠,故常受霜害。由于这些影响,会使树种的自然分布区或人工栽培区受到限制。
因为光周期对树木生活和适应性的影响,树种分布区内,光周期反应有相当大的变化,故有光周期生态型的分化。如树种分布区的南部和北部、高海拔和低海拔、谷地和坡地。这些不同环境中的树木,光周期适应可能不同。已知杨、云杉、松、落叶松和桦属,产自不同纬度和海拔的种源培育的实生苗,在顶芽形成的光周期反应上表现出明显的生态群变异,即产自北方和高海拔的种源培育的实生苗比产自南方和低海拔的,在较长的日照下形成顶芽和停止高生长[2,3,13]。
因此,在树木引种时,除了要考虑温度因素外,还要考虑光周期现象。但并非所有树种对光周期都有强烈反应。有些树种对日照强度敏感,如,油橄榄原产区年日照总时数在2 500 h以上,在雅典达2 756 h,太阳总辐射量每年为556×109J/cm2。这是我国引种油橄榄的限制因子之一[13]。
2.3 光照强度与林业生产
太阳辐射投射到森林群落中,由于林冠的存在,其强度和性质均发生改变。林内太阳辐射分布、林内光照强度的垂直分布与森林经营有着密切的关系。地表光照强度的时空变化规律是随纬度的增加而减弱,随海拔的升高而增强,南坡的辐射量大于北坡且坡度越大差别越大,一年中以夏季最强冬季最弱,一天中以中午最强早晚最弱。因此,不同的生态地理种有其特定的光照强度[3,24~26]。这在南北、高低海拔的引种中必须加以注意。
2.3.1 林冠的反射率、吸收率和透射率
太阳辐射投射到林冠被林冠层反射、吸收,只有部分透过林冠到达林内。这种能力受林冠叶面积大小、叶片厚度、绿色程度和表面性状不同而不同;不同林种则受树种、种植方式和密度及经营方式的影响而不同。反射率一般在20%左右,吸收率一般在70%左右,透射率一般在10%左右。例如,北方针阔叶混交林,上层树冠吸收的光占79%,反射的光占10%,射入群落下层的光为11%。松林林冠反射的光占12%,吸收的光占58%,射入松林下层的光占30%[2]。光强影响林下地被物的种类、数量和生长。据研究,如果林内透光率在0.6%以下,则草本植物不能生长,当达到2%~3%,才能较好地生长发育;如果达到4%~6%,则林下杂木也能生长[1]。这对于开展多种多层经营,建立经济林人工生态群落具有实践意义。
2.3.2 林内的光斑和阴影
到达林内的太阳辐射可分为二部分,一部分是通过林冠枝叶的间隙射到林内的直射光,一部分为透过林冠枝叶的散射光和叶片的透射光。投射到林内的直射光表现为大小不同的光斑,光斑多为园形,有时呈片状。光斑随林分的郁闭度、层次增加而减少。由于林冠空隙的小孔衍射和放大作用,园形光斑的太阳辐射强度比空旷地的弱,其减弱程度与树冠中透光空隙至林地光斑的距离的平方成反比。据观测,直径为5 cm的光斑,其强度减弱50%左右。片状光斑的太阳辐射强度则与空旷地差别不大。
透过林冠枝叶的散射光和叶片的透射光组成林内的阴影,其辐射强度一般仅为空旷地的50%~70%,并以绿光为多。在空旷地的散射辐射中含生理辐射的50%左右,而林内,由于林冠的多次反射和吸收,往往只含生理辐射的30%左右,在密林中可减少到10%左右。
在光斑和阴影的明暗交替的断续光下,有些植物如葡萄(A,C,Leopld,1975)的光合强度比连续光下的光合强度高,但明暗交替的周期太长则下降[1]。
2.3.3 林内光照的垂直分布
林内光强在垂直方向的变化差异很大,据Heinick(1963)研究[1],把树冠每向下91 cm分为一层,则其相对光照强度是:顶层为全光照的93%、2层为70%、3层为42%、4层为25%、5层为21%,光照强度随着叶幕的加深而急剧降低。如用数学模型表达则是:

式中,dQ为太阳辐射透过林冠时的减弱强度,K为林冠对太阳辐射的衰减系数,或者称林冠的消光系数,其值是一个林木群体特征,决定于树种、种植方式和密度、林龄、郁闭度、林冠枝叶稀密度等,可以实测。对于一个具体的林分,K为常数。Q是投射到林冠表面上的太阳辐射光照强度;dM为林冠层厚度。其积分式为:

式中,Q'为透过林冠到达林内的太阳辐射光照;M是林冠厚度。据Heinicke研究,叶面积系数与叶幕光能分布密度相关。如果把M改用叶面积系数L来表示,则Q'=Q-KL,此式就是Monsi公式。如果把林冠分为若干层次,即可求出林冠各层的光照强度[1]。
3 林业生产中提高光能利用率的途径
所谓光能利用率,是光合作用贮存的化学能与接受的有效光量之比[1,2,5],直接影响生物生产量,且受太阳辐射、温度、水分、养分等自然条件限制。光能利用率表示的方法很多,有的用植被的总生产量除以同一时间、同一面积植被吸收的光合有效辐射;有的用净生产量求算;有的用太阳辐射吸收率来表示等等。不同植物群落以及同一植物群落在不同的气候条件下具不同的光能利用率。以净初级生产量计算的光能利用率,热带雨林最高,为1.5%,北方针叶林0.75%,冻原为0.25%,半荒漠最低,为0.04%[3]。根据全球范围内的计算,植物光能利用率欧洲为 1.26%,南美洲为 1.13%,北美洲为0.94%,亚洲为0.88%,非洲为0.59%,澳大利亚为0.44%,前苏联为 1.09%[4]。
不同林分类型、树种特性、林分起源的光能利用率差异较大。张祝平用红外线CO2气体分析法和量子传感器分层法在鼎湖山测定了阔叶林和针阔混交林的光合有效辐射,结果表明,阔叶林总生产力的光能利用率为14.28%,混交林为12.01%[9]。阳性树种比阴性树种的光能利用率高,如马尾松的光能利用率为 2.03%2.17%[8],红松为 1.55%[9]。天然林比人工林的光能利用率高,如任豆林的天然林和人工林分别为5.43%和1.39%[10]。
由此可见,植物的光能利用率一般是很低的,但潜力也很大。因此,在林业生产中,如何设法提高林木的光能利用率问题,越来越显出其重要性。如,培育或筛选高光合效能和低光呼吸的优良树种,或保持森林的适宜密度和叶面积指数,以充分利用光能。更新造林初期,幼林郁闭前,进行林下间作或充分利用喜光先锋树种的快速更新;营造各种类型的混交林和异龄林以形成多层次的林冠结构和垂直郁闭;根据树种特性、年龄阶段、环境条件及生长规律,及时通过间伐及人工修枝措施,调整林分密度及郁闭度。
随着林业科学的发展,今后在选育高光能利用率的林木品种,模拟天然林合理的林分结构及制订整套科学的经营措施和管理方案,将成为有效提高森林光能利率的方向。
[1]王利溥.经济林气象[M].昆明:云南科技出版社,1995.
[2]曲仲湘,吴玉树.编著.植物生态学(第二版)[M].北京:高等教育出版式社,1983.
[3]李景文.森林生态学[M].北京:中国林业出版社,1994.
[4]穆彪.农业气象学[M].贵阳:贵州科技出版社,1997.
[5]穆彪.试论贵州省太阳辐射条件与林木生长[J].贵州农学院报,1991,10(1):46~52.
[6]赵宝忱,刘自强,吴淑杰.不同森林群落结构与光能利用率的关系[J].植物研究,1995,15(.2):256~262.
[7]张祝平.鼎湖山森林群落的光能利用效率[J].植物生态学与地植物学学报,1990,14(2):139~149.
[8]宛志沪,刘先银.大蜀山马尾松人工林晴大太阳辐射光谱及光能利用率的研究[J].中国农业气象,1994,15(1):25~29.
[9]张彦东,卢伯松,丁冰.红松人工林能量环境与光能利用率的研究[J].东北林业大学学报,1993,21(1):35~42.
[10]张祝平,何道泉,敖惠修.任豆林的生物量和光能利用率[J].植物生态学报,1996,20(6):502~509.
[11]孙时轩.造林学(第二版)[M].北京:中国林业出版社,1992.
[12]叶镜中,孙多.森林经营学[M].北京:中国林业出版社,1995.
[13]沈熙环.林木育种学[M].北京:中国林业出版社,1990.
[14]李俊清,传照,姚成滨.林冠下红松幼树叶绿素含量与生长关系的研究[J].东北林业大学学报,1990,18(2):21~26.
[15]刘志则,马钦彦,潘向丽,等.华北落叶松人工林放叶过程中的辐射特征[J].生态学报,1997,17(5):519~524.
[16]戴继光.阳光照射对樟子松雨季造林成活率的影响[J].西北林学院学报,1998,13(4):54~57.
[17]田国良.呼伦贝尔草原的太阳分光辐射能和光合潜力[J].地理学报,1980,35(1):76~82.
[18]苏睿丽,李 伟.沉水植物光合作用的特点与研究进展[J].植物学通报,2005,22(增刊):128~138.
[19]王文林,王国祥,李强,等.水体浊度对菹草(Potamogeton cripus)幼苗生长发育的影响[J].生态学报,2006,26(11):3586~3593.
[20]Ochieng C A,Short F T,Walker D I.Photosynthetic and morphological responses of eelgrass(Zostera marina L.)to a gradient of light conditions[J].Journal of Experimental Marine Biology and Ecology,2010,382:117~124.
[21]Backman T W,Barilotti D C.Irradiance reduction:effects on metabolism,growth and survival[J].Aquatic Botany,1995,51:67-86.
[22]Mark J B,Scott W N.Direct measurements of light attenuation by epiphytes on eelgrass Zostera marina[J].Marine Ecology Progress Series,2002,238:73~79.
[23]Korschgen C E,Green W L,Kenow K P.Effects of irradiance on growth and winter bud production by Vallisneria americana and consequences to its abundance and distribution [J].Aquatic Bontany,1997,58:1~9.
[24]Joanne C B,Scott W N.Responses of eelgrass Zostera marina seedlings to reduced light[J].Marine Ecology Progress Series,2001,223:133~141.
[26]Ruiz J M,Romero J.Effects of disturbances caused by coastal constructions on spatial structure,growth dynamics and photosynthesis of the seagrass Posidonia oceania [J].Marine Pollution Bulletin,2003,46:1523~1533.
猜你喜欢
农业科技通讯(2023年1期)2023-02-12
环保科技(2020年2期)2020-05-21
浙江农林大学学报(2019年6期)2019-11-13
装备环境工程(2019年9期)2019-10-24
浙江农业学报(2019年7期)2019-08-14
中国新技术新产品(2019年21期)2019-01-20
电子技术与软件工程(2017年23期)2018-01-17
少儿科学周刊·少年版(2017年7期)2017-09-29
少儿科学周刊·儿童版(2017年7期)2017-09-29
科学家(2016年12期)2017-09-29
