
扁角豆芫菁幼虫取食与生长发育关系研究
2015-11-25 02:58母银林陈祥盛
环境昆虫学报 2015年2期
母银林,陈祥盛
(贵州大学昆虫研究所,贵州昆虫资源开发利用特色重点实验室,贵州山地农业病虫害重点实验室,贵阳 550025)
扁角豆芫菁Epicauta impressicornis Pic 亦称凹角豆芫菁,为鞘翅目Coleoptera 芫菁科Meloidae 昆虫。体内含斑蝥素,药用功能与斑蝥 Lytta vesicatoria 相似,且斑蝥素含量比斑蝥高(李晓飞等,2007),不仅具有抗肿瘤、发泡、利尿作用,还有杀虫、除草、抗病毒、抗菌等作用。此外,其幼虫对中华稻蝗Oxya chinensis Thunb.捕食率较高,对控制蝗虫种群数量与为害方面有着重要作用(李晓飞等,2006)。
以资源昆虫开发为目的芫菁生物学研究主要集中在我国的西南和华东地区,初步揭示了短翅豆芫菁Epicauta aptara Kaszab (胡周强等,2000;封孝兰等,2012)、大斑芫菁Mylabris phalerata Pallas (胡周强等,1994,1998;黄文华,2001)、锯角豆芫菁Epicauta gorhami Marseul (王雪梅,2007)、毛胫豆芫菁Epicauta tibialis Waterhouse(王雪梅,2007)和眼斑芫菁Mylabrls clchorii Linnaeus (汪会荣,2008)等物种的生物学特性。但缺乏扁角豆芫菁幼虫取食与生长发育的关系研究,严重制约着扁角豆芫菁资源开发的进度。开展本项研究,不仅能明确其供食量和生长发育的关系,还将有助于进一步了解芫菁幼虫的觅食行为,对芫菁的人工养殖有重要意义。为此,笔者在室内对扁角豆芫菁幼虫取食作了试验探索,现将结果整理报道如下。
1 材料与方法
1.1 虫源及主要试验仪器
1.1.1 虫源
试虫为自养扁角豆芫菁繁殖所得。幼虫以中华稻蝗卵为食物,部分中华稻蝗采至贵州省贵阳市花溪区附近稻田内,部分通过留种繁殖所得。收集卵块,放在冰箱中保存备用。
1.1.2 主要试验仪器
RAZ-380A 型人工气候箱(宁波江南仪器厂);亚都YC-D204 型超声波加湿器(北京亚都室内环保科技股份有限公司);木质纱网饲养笼体积(40 cm×40 cm×70 cm);200 mL 富强一次性塑料杯 (贵州富强包装有限公司);电子天平(YP5002 型,上海佑科仪器仪表有限公司);体积为1000 mL的蜀牛烧杯(GG-17 型,上海华增玻璃仪器有限公司);美的BCD-195CMJ 型电冰箱(合肥美的荣事达电冰箱有限公司)。
1.2 方法
1.2.1 蝗卵埋藏深度对1 龄幼虫取食影响
在室内人工气箱内,光照条件L∶D=14 ∶10,温度为恒温32℃±0.2℃,湿度为60%条件下,在200 mL的一次性塑料杯,装半杯含水量为30%的土壤。接入1 龄幼虫和蝗卵,之后用含水脱脂棉覆盖,杯口用保鲜膜封口,再用镊子在保鲜膜上扎孔用以通气。试验设置4个处理,在杯中培养土中挖一小洞,放入蝗虫卵块,然后覆上松土,设置深度 (卵块底部离土表的距离)0 cm、0.5 cm、1 cm、2 cm。待接入1 龄幼虫后,后面处理同上。每个处理5 杯为一个重复,每杯1 头幼虫,相应放入1 块蝗卵,3 次重复。观察1 龄幼虫死亡情况,发现有死亡幼虫,重新接入。观察取食情况,每3 h 检视一次,记录1 龄幼虫从接入到找到蝗卵,开始取食之前所耗用的时间,即寻食期。
1.2.2 幼虫取食量与其生长发育的关系
为探明扁角豆芫菁幼虫取食阶段(1-7 龄)的取食情况,设置3个处理,分别为半蝗虫卵块(约15-20 粒蝗卵),1个卵块 (40-50 粒)、2 卵块(相连)、2 卵块(相离)。卵块放好后,接入1 头活泼的1 龄幼虫,以后方法同上。每处理5头虫,3 次重复。观察1 龄幼虫死亡情况,发现有死亡幼虫,重新接入。观察幼虫发育情况,记录幼虫寻食期、取食期、蛹化虫数、羽化虫数,幼虫羽化后称量其成虫体重。
2 结果与分析
2.1 蝗卵埋藏深度对1 龄幼虫取食影响
设置的4个处理中,寻食期长短不一,蝗卵埋藏深度越深1 龄幼虫寻食期越长(表1)。试验结果表明,蝗卵埋藏深度为0 cm 和0.5 cm的处理,其幼虫均能很快获取食物(图1),平均耗时在18.5 h 左右,取食率达93.33%。蝗卵埋藏深度在1 cm 和2 cm的处理,幼虫寻食期较长,为31.73 h 和41.26 h,且取食率降低,分别为73.33%和53.33%。这说明扁角豆芫菁幼虫嗅觉能力弱,主要以埋藏于土中0.5 cm 深或者与土表面的蝗虫卵块为食。这与野外稻蝗产卵习性:多产于土表、草丛、稻桩、草兜中相吻合。分别对取食率和寻食期作方差分析,再经多重比较,结果表明蝗卵埋藏深度为0 cm 和0.5 cm 分别与2 cm之间,1 龄幼虫取食率和差异显著;与1 cm 之间,1 龄幼虫取食率差异不显著;蝗卵埋藏深度1 cm与2 cm 之间,1 龄幼虫取食率差异显著。在寻食期方面,蝗卵埋藏深度为0 cm 和0.5 cm 分别与1 cm和2 cm 之间差异显著;蝗卵埋藏深度为1 cm和2 cm 之间也表现为差异显著。
2.2 幼虫取食量与其生长发育的关系
饲养中观察到,扁角豆芫菁1 龄幼虫都能在短时间内找到并取食蝗卵,与供卵量多少无关,同时发现幼虫有残杀现象。1/2 块蝗卵的处理中,当蝗卵取食完后,幼虫出卵鞘,不再取食第二块卵(不论卵块分开与否),查幼虫的发育进度,此刻为4 龄幼虫,随后直接入土化为假蛹,继续发育,最后羽化为成虫,但是体形瘦小。取食1 卵块的处理,幼虫发育正常,到5 龄末期才入土化为假蛹,成虫体形肥大,体重和体长约为前者的2 倍,不过有的卵块较大,直到幼虫入土前也不能吃完。2 块蝗卵的处理中,幼虫吃完第1 块卵后入土,不再取食第2 块卵(两卵块连在一起的),最后正常羽化,成虫体形肥大。在试验过程中,观察到幼虫有残杀习性,有的发生在1 龄幼虫之间;有的发生在3-4 龄幼虫之间。

图1 蝗卵埋藏深度对1 龄幼虫取食影响Fig.1 Impacting on first instar`s feeding by locust eggs buried depth

表1 1 龄幼虫取食与蝗卵埋藏深度的关系Table 1 The relationship between food buried depth and feeding larvae number
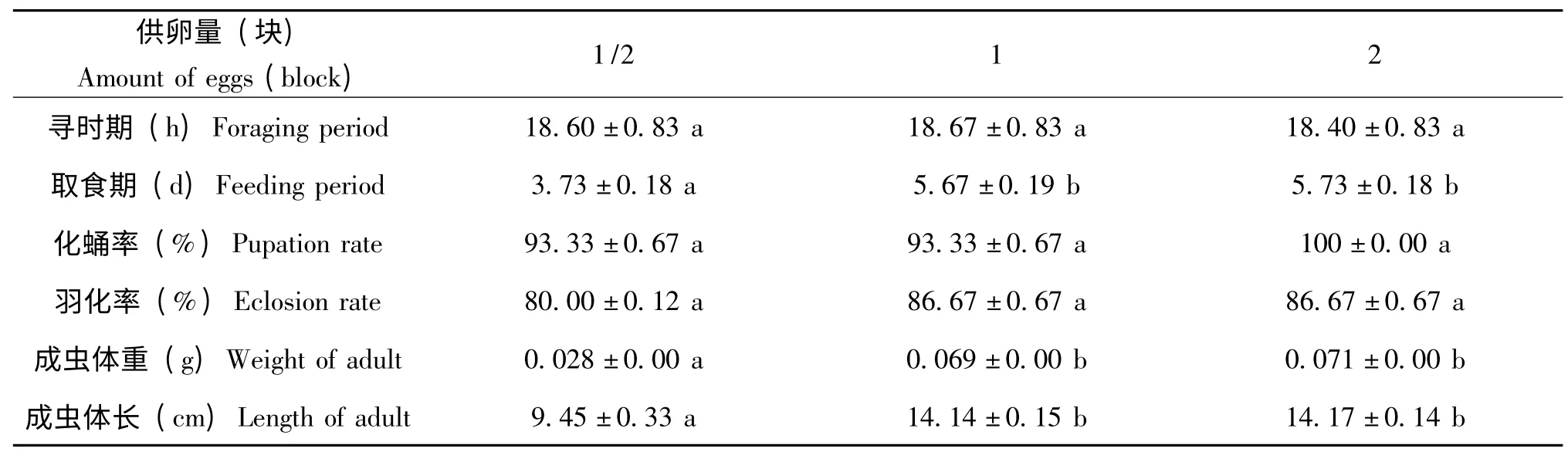
表2 幼虫取食量与其生长发育的关系Table 2 The relationship between larvae taking food intake and its growth
从表2 结果表明,在寻食期,幼虫耗时约18.5 h,对3个处理间进行方差分析,差异不显著。在取食期阶段,取食1/2 块蝗卵的幼虫,耗时约比取食1 块和2 块蝗卵的少一半。经方差分析,取食1/2 块蝗卵的处理与取食1 块和2 块蝗卵的处理间差异显著;取食1 块和2 块蝗卵的处理间差异不显著。在适宜的条件下,3个处理中,幼虫都表现除了较高的化蛹率和羽化率,都达到80%以上。经过方差分析,各个处理间差异不显著。蛹羽化为成虫后,取食1/2 块蝗卵的幼虫体形瘦小,经过称量约0.03 g。取食1 块和2 块蝗卵的幼虫,羽化后成虫体形肥大,称得体重约0.07 g,是取食1/2 块蝗卵的成虫体重的2 倍还多。经过方差分析和多重比较,取食1/2 块蝗卵的处理与另两个处理间差异显著,取食1 块蝗卵和2 块蝗卵的处理间差异不显著。
3 结论与讨论
扁角豆芫菁在蝗卵埋藏的深度(从卵底部与土壤接触算起)为0 cm、0.5 cm 时寻食期最短,可以推知寻食期在0-0.5 cm 内最短。不同蝗卵量的处理,对寻食期进行方差分析,并进行多重比较,彼此间差异不显著。表明食期寻食期长短与供卵量无关。胡周强等(2012)报道芫菁1 龄幼虫的取食与蝗卵埋藏深度密切关系,蝗卵埋藏的深浅决定了1 龄幼虫寻食期的长短,甚至不能找到食物,在蝗卵埋藏的深度(从卵顶部与土壤接触算起)为0 cm、0.5 cm 是寻食期最短。本研究结果与文献报道一致,同样说明蝗卵埋藏的深度对芫菁1 龄幼虫寻食期有影响,在蝗卵埋藏的深度(从卵底部与土壤接触算起)为0 cm、0.5 cm时寻食期最短。比较两个研究结果,说明芫菁不同种间,寻食期最短时的蝗卵埋藏深度有所不同。研究中观察到,当埋藏深度为0 cm 时,全部幼虫取食从卵的底部开始采食;当埋藏深度为0.5 cm时,幼虫采食点多样化,除了露出部分,其它地方都有幼虫采食点。因此,为了方便幼虫采食,建议在扁角豆芫菁饲养中,采用蝗卵埋0.5 cm 深的方式。
本研究表明食量与芫菁生长发育有着密切关系。扁角豆芫菁幼虫饲养中供卵量为1 块时最合适,在此条件下幼虫能正常发育为体形肥大的成虫。在投放饲料前,先称量了卵块重量,卵块鲜重为0.6 g/块能完全被吃完,较大的 (约0.9 g/块)有剩余而造成浪费;而取食1/2 块蝗卵的幼虫提前入土化为假蛹,说明食物不足是导致幼虫提前入土化蛹的原因;表明扁角豆芫菁幼虫对蝗卵的最适需求量约0.6 g 重,建议在以后的扁角豆芫菁饲养工作中,选用重量约为0.6 g 蝗卵进行饲养。这给幼虫人工饲料的研制提供了理论指导。
大斑芫菁和短翅豆芫菁幼虫取食量不同,其寻食期、生长期、入土深度,虫体大小均有一定差异(张含藻等,1994;胡周强等,2012)。说明食量的不同影响两种芫菁的寻食期、生长期、入土深度,虫体大小,两种芫菁寻食期随着蝗卵量的增加而缩短。本研究结果表明食量不同,幼虫在生长期虫体大小上也有差异,但寻食期、入土深度彼此间无差异。扁角豆芫菁寻食期长短不受蝗卵量的影响,而是受蝗卵埋藏深度影响。寻食期的长短影响了芫菁1 龄幼虫历期的长短,进而影响了其整个生活史。因此,研究如何缩短1 龄幼虫寻食期的长短对芫菁的多代饲养有重要意义。
由于本实验中采用的是一次性塑料杯,装土不深,观察到几乎所有的幼虫都是在杯底化蛹的。这可能是导致其入土深度彼此间无差异性的原因。若采用其它装置,装更深的土壤能否影响幼虫的入土深度,还有待进一步研究。芫菁幼虫中,1 龄幼虫在取食阶段残杀最多,3 龄和4 龄残杀相对较少。本研究中,也发现在扁角豆芫菁龄幼虫间有残杀现象,幼虫在同一卵块不同部位取食时,不会发生残杀,当两头幼虫取食相遇后,就会为争夺食物而相互残杀,有时能存活一头幼虫,有时两只幼虫相互残杀致死。残杀会导致幼虫死亡,增加了幼虫的死亡率,从而影响饲养产量,成为限制规模化养殖的一个重要因素。因此,幼虫残杀问题是下一步试验的重点。
References)
Feng XL,Hu ZQ,Zhen DJ.Study on predation habits of Epicauta aptera Kaszab larvae [J].Special Wild Economic Animal and Plant Research,2012,1:19-22.[封孝兰,胡周强,曾德军.短翅豆芫菁幼虫采食习性的研究[J].特产研究,2012,1:19-22]
Li XF,Chen XS,Guo XM.Biological characteristics of Epicauta impressicornis and its control effect to Oxya chinensis [J].Hubei Agricultural Sciences,2006,45 (4):458-459.[李晓飞,陈祥盛,国兴明.凹角豆芫菁的特征特性及对稻蝗的捕食作用[J].湖北农业科学,2006,45 (4):458-459]
Li XF,Chen XS,Wang XM.Investigation on the resource of containing cantharidin insects in Guizhou [J].Hubei Agricultural Sciences,2007,46 (2):300-301.[李晓飞,陈祥盛,王雪梅.贵州含斑蝥素昆虫资源调查及斑蝥素含量的测定[J].湖北农业科学,2007,46 (2):300-301]
Hu ZQ,Zhang HZ.The relationship between blister beetle larvae and food intake [J].Journal of Chinese Medicinal Materials,1994,17 (5):9-11.[胡周强,张含藻.斑蝥幼虫与食量的关系[J].中药材,1994,17 (5):9-11]
Hu ZQ,Xiao JY,Wei HP.Study on the biology of Epicauta aptera Kaszab[J].Entomological Knowledge,2000,37 (5):287-289.[胡周强,肖杰易,韦会平.短翅豆芫菁生物学特性研究[J].昆虫知识,2000,37 (5):287-289]
Huang WH.The Studies on Artificial Rearing and Breeding Ecology of Meloids [D].Fujian:Fujian Normal University,2001.[黄文华.芫菁的人工饲养和生殖生态研究[D].福州:福建师范大学,2001]
Wang HR.Study on the Level of Cantharidin in Mylabris cichorii Linnaeus and artificial rearing [D].Chongqing:Chongqing University,2008.[汪会荣.眼斑芫菁的人工养殖及其斑蝥素含量的研究[D].重庆:重庆大学,2008]
Wang XM.Studies on Biology and Artificial Feeding of Epicauta gorhami Marseul and Epicauta tibialis Waterhouse [D].Guizhou:Guizhou University,2007.[王雪梅.锯角豆芫菁与毛胫豆芫菁的生物学及人工饲养研究[D].贵州:贵州大学,2007]
Zhang HZ,Hu ZQ.Study on the biology of Mylabris phalerala Pallas[J].Journal of Chinese Medicinal Materials,1998,11 (3):15-19.[张含藻,胡周强.大斑芫菁生物学特性研究[J].中药材,1988,11 (3):15-19]
猜你喜欢
环境昆虫学报(2020年6期)2021-01-18
中学生数理化·七年级数学人教版(2020年11期)2020-12-14
植物保护(2019年5期)2019-11-27
艺术品鉴证.中国艺术金融(2018年8期)2019-01-14
艺术品鉴证.中国艺术金融(2018年10期)2019-01-08
艺术品鉴证.中国艺术金融(2018年12期)2018-08-26
金色少年(奇趣科普)(2017年4期)2017-06-05
农村农业农民·B版(2016年7期)2016-10-21
天然产物研究与开发(2014年8期)2014-04-27
中国蔬菜(2012年24期)2012-02-24
