
TMV侵染后烟草中信号分子水杨酸和过氧化氢的变化
2015-11-24 07:13:59顾厚武何宽信刘再强徐庆凯郭家明陈学平
中国烟草学报 2015年5期
顾厚武,何宽信,刘再强,徐庆凯,郭家明,陈学平
1 江西省烟草公司,江西南昌洪城路298号330025;2 中国科学技术大学烟草与健康研究中心,安徽合肥徽州大道1129号 230052
TMV侵染后烟草中信号分子水杨酸和过氧化氢的变化
顾厚武1,何宽信1,刘再强1,徐庆凯1,郭家明2,陈学平2
1 江西省烟草公司,江西南昌洪城路298号330025;2 中国科学技术大学烟草与健康研究中心,安徽合肥徽州大道1129号 230052
以两个抗病品种、一个感病品种为材料,利用抗体和定位染色对TMV侵染后烟草中的信号分子水杨酸和过氧化氢的时空变化进行检测。结果表明:接种TMV能够诱导水杨酸(SA)含量的上升,到接种后12h达到最大,且接种叶比同侧上部叶上升幅度要大,品种间存在差异;抗病品种接种TMV能够诱导过氧化氢的产生,随着时间延长过氧化氢从局部向外扩散,而感病品种和磷酸盐缓冲液(PBS)对照不能激发过氧化氢的产生。
TMV;信号转导;烟草
植物具有一整套细胞学机制去保护自己抵御病原菌的侵袭。在许多情况下寄主识别病原菌激活所谓的超敏反应(hypersensitive response, HR),这个超敏反应是具有一系列特征的抗性反应,如定位细胞死亡、产生超氧离子和过氧化氢、葡聚糖沉积、细胞壁增强、次生代谢产物和对微生物具有抗性活性的蛋白质的合成[1-3]。HR在表型上存在着差异,从单细胞HR到扩散型坏死区域,其主要作用是有效地限制病原菌在宿主体内的繁殖、定居及扩散。与这种局部反应相关的是,经过几天到一周时间被侵染的植物产生新的抗性,从而对病原菌的再次侵染甚至其它病菌的感染均具有很强的抗性。这种抗性水平可以扩散到整个植株,通常称为系统获得性抗性(systemic acquired resistance,SAR)[4-6]。在实现SAR过程中,水杨酸(SA)、茉莉酸(JA)、乙烯、过氧化氢(H2O2)等信号分子的转导起着至关重要的作用。不同的作物涉及到不同的信号分子,烟草对TMV系统获得性抗病性中的信号分子是水杨酸和过氧化氢。虽然已经有一些研究报道了这些信号分子的作用[7-10],但其时空变化以及品种间的差异性未见报道。本研究采用特异抗体和定位化学方法对病程过程中信号分子水杨酸和过氧化氢进行测定,并分析品种间的差异性,以期在系统获得性抗病性信号转导机理方面作些探索。
1 材料与方法
1.1 参试品种
参试品种包括具有系统获得性抗病特性的品种Xanthi-nc、HZNH和感病品种K358。
1.2 样品处理
上述品种常规方法育苗,烟苗8叶左右移栽到盆钵中,待烟苗12叶左右时,分别在不同烟株的第5叶上,采用涂抹方法沿叶片主脉两侧接种TMV、进行SA涂抹和PBS涂抹。每品种各处理12株(其中6株用于测定SA、6株用于H2O2的的定位观察),3次重复。
水杨酸测定样品:处理后2h,4h,6h,8h,12h,24h取各处理叶和同侧上部叶0.5g左右剪碎放置在小瓶中,加5mL甲醇并封口,置于零下20℃中保存待测。
过氧化氢定位观察样品:TMV接种2h,6h,12h,24h、SA涂抹6h,12h,24h后取各处理叶和同侧上部叶,立即进行定位观察的相关处理。
1.3 水杨酸测定和过氧化氢定位观察方法
水杨酸测定:利用南京农业大学植物生理组研制的抗体对待测样品进行SA检测;
过氧化氢定位观察:将样品置于千分之一的DAB溶液中染色4h,而后用70%乙醇脱色数小时。
2 结果与分析
对不同品种实施接种TMV、SA涂抹和PBS涂抹后,在不同时间段对处理叶和同侧上部叶进行SA和过氧化氢定位观察,结果如下:
2.1 不同处理下信号分子SA的动态变化
由图1可见,接种TMV后烟草抗病品种HZNH的接种叶和上部叶的SA含量均呈上升趋势,在观测时间段内,到12h时达到最大,到24小时时有所下降。其中接种叶比上部叶上升的幅度要大一些,而相应没有接种的对照叶SA含量在几次取样中没有明显的变化,该差异暗示SA是由TMV接种诱导的。
涂抹SA后,其同侧上部叶SA含量均较对照要高(图2),其中涂抹6小时后比对照的SA含量差异更加明显。说明涂抹SA同样可以诱导同侧上部叶内源SA含量的增加。而对照的同侧上部叶SA含量基本无变化。

图1 TMV接种对HZNH品种烟草下部接种叶、上部叶片中的SA含量的影响Fig.1 Effect of TMV injection on SA content in the upper and the lower leaves of HZNH tobacco plant

图2 SA处理HZNH后同侧上部叶SA含量Fig.2 SA content in ipsilateral upper leaves of HZNH after SA treatment
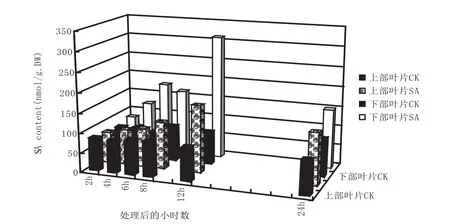
图3 TMV接种对Xanthi-nc品种烟草下部接种叶、上部叶片中的SA含量的影响Fig.3 Effect of TMV injection on SA content of the upper and the lower leaves of Xanthi-nc tobacco

图4 SA处理后Xanthi-nc同侧上部叶SA含量的变化Fig.4 SA content changes in ipsilateral upper leaves of Xanthi-nc after SA treatment
与烟草品种HZNH相似,烟草抗病品种Xanthi-nc在接种TMV后接种叶和同侧的上部叶SA含量呈现上升趋势,在观测时间段内,最大值均出现在接种后的12h(图3),但后者的上升斜率大于前者,说明烟草品种Xanthi-nc在TMV诱导后体内SA激活表达比HZNH更为敏感。涂抹SA处理后,其同侧上部叶较相应的对照叶SA的含量有一定上升,在12小时时更加明显(图4)。

图5 TMV接种对K358品种烟草下部接种叶、上部叶片中的SA含量的影响Fig.5 Effect of TMV injection on SA content in the upper and the lower leaves of K358
对于感病品种K358而言,进行接种TMV处理后接种叶和同侧上部叶SA的含量也表现出明显的上升趋势(图5),接种叶的SA含量上升幅度比抗病品种Xanthi-nc小、与HZNH相当,同侧上部叶SA的含量上升幅度远小于抗病品种Xanthi-nc、稍小于HZNH。虽然感病品种K358在接种TMV后也激发了信号分子SA的表达,但并没有表现出抗病特性,这可能与不同遗传型对SA敏感程度及其它生化途径的差异有关系。
同样,感病品种K358在涂抹SA后其同侧上部叶SA含量也比对照处理代谢高一些,但在不同时间段处理和对照间的差异没有明显的变化(图6)。这与抗病品种HZNH、Xanthi-nc相比存在差异。

图6 SA处理后烟草品种K358接种叶和上部叶SA含量的变化Fig.6 SA content changes in the upper and the lower leaves of K358 afer SA treatment

图7 不同处理下不同烟草品种过氧化氢的动态变化Fig.7 Dynamic changes of H2O2 among different varieties under different treatments
2.2 不同处理下过氧化氢的动态变化
在对3个烟草品种进行接种TMV、涂抹SA和涂抹PBS处理后进行染色,观察过氧化氢的动态变化,其结果见图7。图中出现褐色的是过氧化氢染色部分,颜色越深过氧化氢越多。在3个品种中,感病品种K358接种TMV和涂抹SA处理后均没有观察到明显的过氧化氢(图7-A,B)。而抗病品种Xanthinc和HZNH在进行相关的处理2h后均可以观察到明显的过氧化氢,并在随后的几个时段观察发现过氧化氢迅速增加并向四周扩散(图7-C,D,E,F)。在PBS处理后没有观察到明显的过氧化氢的产生(图7-G)。说明感病品种不具有被诱导激活的防卫反应机制,PBS也不是诱导物质。
3 讨论
3.1 SA和H2O2在系统获得性抗病性中作用的分析
水杨酸作为植物抗病反应的重要信号分子,涉及并参与植物的超敏反应(HP)和SAR反应,在植物的SAR信号转导和抗病反应中起着关键作用[11-13]。许多研究表明植物在HP的最初阶段,病原菌或激发子与位于原生质膜的受体结合后,G蛋白可能通过活化蛋白激酶增加与诸如O2.-、H2O2等活性氧密切相关的原生质膜上的NAD(P)H氧化酶、依赖于NADP的过氧化物酶和超氧化物合酶等活性,从而最终导致氧化迸发(oxidative burst)[13]。而O2.-、H2O2等活性氧不仅可以直接杀伤病原菌,参与膜脂过氧化,介入HP,而H2O2还可以促进细胞壁木质化和HP细胞死亡。已经发现病原菌侵染后SA水平急剧增加,而且SA可以同时通过增加SOD等H2O2产生酶类的活性和抑制CAT/APX等降解酶类的活性,最终积累H2O2来提高植物的抗病性[12]。我们另外一个实验中,与本实验相同的两个具有SAR的抗病品种也获得了枯斑症状[5],并且有效地阻止了花叶病的发生。
SA和SA结合过氧化氢酶相互作用导致过氧化氢酶活性的抑制和SA的产生。从机理上讲,这两个偶联事件可能涉及到SA信号转导途径,进而导致SAR的建立和HP的发展。本研究直接观察了烟草在接种TMV和涂抹SA后信号分子过氧化氢的动态变化,为相关的理论解释提供了最直观的证据。同时,我们发现的一个黄花烟品种HZNH也具备这个特征。在另外一个实验中,我们克隆出HZNH的相关抗性基因,发现这两个抗性品种虽然不是一个种,但这个与SAR相关的基因却具有很高的同源性[11]。
3.2 不同烟草品种对SA含量上升的敏感差异性分析
不同作物在产生SAR时的信号分子存在一些差异,有的作物信号分子是SA,而有的是JA[2,6]。第二信使有的是过氧化氢,而有的是乙烯。说明不同遗传型作物本身在形成SAR特性时就已经存在一些差别。对于具有SAR特性的烟草品种来说,SA含量的中度或稍微升高就会诱导抗性产生,但品种间存在一些差异。其原因可能主要有以下两个方面:系统获得性抗病性的诱导产生过程中涉及到一系列生化代谢反应,SA仅仅是激发子诱导激活这些反应,如果被处理植株本身在其它代谢环节中有缺陷,即使SA含量提高也无法激活SAR反应;不同遗传型对SA的敏感程度不同或者有的就钝感,当SA含量没有达到一定的阈值时根本就无法激活相应的反应。具有SAR特性的抗病品种能够形成明显的枯斑,而感病品种则不能[5],本实验中感病品种K358 SA含量上升没有激发SAR可能就是由于缺乏某种机制。其具体原因还需要进一步设计实验加以验证。
4 结论
本研究以国际上公认的TMV抗性品种Xanthinc和作者发现的新抗性品种HZNH为材料,以接种TMV和涂抹SA为诱导因素,在不同时间、部位对信号分子过氧化氢信号和系统物质SA进行了检测,证明这两个抗性材料均属于系统获得抗病性、其主要途径是SA系统、信号转导分子就是过氧化氢,并且新抗性品种HZNH的抗性基因与Xanthi-nc具有很高的同源性、均属于N基因家族成员[11](另外文章中)。而感病品种K358不具有此类特征。这些对于进一步研究TMV抗性机理及改良烟草品种抗病性具有重要的参考意义。
[1]王钧, 金巧玲.植物系统获得性抗性的信号转导 [J].生命科学, 1998, 10(01): 7-13.
[2]李德红, 潘瑞炽.水杨酸在植物体内的作用 [J].植物生理学通讯, 1995, 31(02): 144-149.
[3]蔡新忠, 郑重.水杨酸在植物抗病反应中的作用 [J].植物生理学通讯, 1998, 34(04): 297-304.
[4]余迪求, 岑川, 李宝健, 等.植物系统获得的抗病性和信号传导 [J].植物学报, 1999, 41(02): 115-124.
[5]陈学平, 夏凯, 孔繁明, 等.烟草品种对TMV抗性差异的比较研究 [J].中国烟草科学, 2001, 22(01): 1-3.
[6]Low P, Merida J.The oxidative burst in plant defense:function and signal transduction [J].Physiol Plantarum,1996, 96(3): 533-542.
[7]Chen Z, Silva H, Klessig D F.Active oxygen species in the induction of plant systemic acquired resistance by salicylic acid [J].Science, 1993, 262(5141): 1883-1886.
[8]Hammond-Kosack K, Jones J D.Responses to plant pathogens [J].Biochemistry and molecular biology of plants, 2000, 1102-1156.
[9]Chamnongpol S, Willekens H, Moeder W, et al.Defense activation and enhanced pathogen tolerance induced by H2O2in transgenic tobacco [J].Proceedings of the National Academy of Sciences, 1998, 95(10): 5818-5823.
[10]Dinesh-Kumar S, Tham W-H, Baker B J.Structure–function analysis of the tobacco mosaic virus resistance gene N [J].Proceedings of the National Academy of Sciences, 2000,97(26): 14789-14794.
[11]Zhang GY, Chen M, Guo JM, Xu TW, Li LC, Xu ZS, Ma YZ, Chen XP.Isolation and Characteristics of the CN Gene,a Tobacco Mosaic Virus Resistance N Gene Homolog, from Tobacco.Biochem Genet.2009.47(3-4):301-314.
[12]Chen Z, Malamy J, Henning J, et al.Induction,modification, and transduction of the salicylic acid signal in plant defense responses [J].Proceedings of the National Academy of Sciences, 1995, 92(10): 4134-4137.
[13]Durner J, Shah J, Klessig D F.Salicylic acid and disease resistance in plants [J].Trends Plant Sci, 1997, 2(7): 266-274.
Study on variation of signal molecule SA and H2O2in TMV inoculated tobacco
GU Houwu1, HE Kuanxin1, LIU Zaiqiang1, XU Qingkai1, GUO Jiaming2, CHEN Xueping2
1 Jiangxi Provincial Tobacco Corporation, Nanchang 330002, China;2 University of Science and Technology of China, Hefei 230052, China
Two disease-resistant and one susceptible tobacco varieties were used to determine variation of signal molecule SA and H2O2in tobacco inoculated with TMV by the method of antibody and location staining.Results showed that inoculation with TMV could induce the rise of SA which reached maximum in 12h.SA content in the inoculated leaves rose higher than that in the same side upper leaves, and such difference was found among different varieties.TMV inoculation could also induce the rise of H2O2and facilitate H2O2spread from inside to outside while neither susceptible variety nor control group treated with PBS could.
TMV; signal transduction; tobacco
顾厚武,何宽信,刘再强,等.TMV侵染后烟草中信号分子水杨酸和过氧化氢的变化[J].中国烟草学报,2015,21(5)
江西省烟草公司“提高烟草抗逆性新型调节物质研制及应用”(赣烟2011年98号)和池州市烟草公司“生物绿肥在生产中应用研究”(池烟2011.11.18)
顾厚武,硕士,农艺师,研究方向:烟叶栽培,Email:359216119@qq.com
陈学平,博士,教授,研究方向:烟草生物技术、烟草栽培生理生化,Email:chenxp08@ustc.edu.cn
2014-07-13
:GU Houwu, HE Kuanxin, LIU Zaiqiang, et al.Study on variation of signal molecule SA and H2O2in TMV inoculated tobacco[J].Acta Tabacaria Sinica, 2015,21(5)
猜你喜欢
浙江林业科技(2021年3期)2021-07-13 09:03:56
新农村(2018年24期)2018-09-14 06:55:38
长江大学学报(自科版)(2018年14期)2018-07-17 08:37:02
中国中药杂志(2017年11期)2017-06-22 11:37:34
中国中药杂志(2017年1期)2017-03-06 21:19:22
湖北农业科学(2015年23期)2016-01-08 20:37:10
中国药物应用与监测(2015年5期)2015-12-11 03:15:53
化工进展(2015年6期)2015-11-13 00:27:14
结核与肺部疾病杂志(2015年3期)2015-07-18 11:08:56
应用化工(2014年11期)2014-08-16 15:59:13
