
桔小实蝇体重和糖原的积累及其飞行消耗
2015-11-22 08:01袁瑞玲王晓渭
环境昆虫学报 2015年2期
袁瑞玲,杨 珊,王晓渭,陈 鹏
(1.云南省森林植物培育与开发利用重点实验室,昆明650201;2.云南省林业科学院,昆明650201;3.西南林业大学,昆明650224)
桔小实蝇Bactrocera dorsalis(Hendel)又名东方果实蝇,属双翅目Diptera、实蝇科Tephritidae,是40多科250多种水果和蔬菜的重要害虫,因其生活周期短、寄主广泛、繁殖力强、危害性大,被称为“水果杀手”,一直被世界各国列为重要的检疫性害虫 (Drew and Hancock,1994;Chen and Ye,2008;Suganya et al.,2010;Arévalo - Galarza and Follett,2011)。桔小实蝇1911年在中国台湾最早被记录,经过100年的迁移和扩散,其已在中国长江以南的大部分地区分布危害,尽管强迁飞能力已被报道是造成其分布和发生区不断扩张,以及根除后又再次发生的重要原因之一 (陈鹏等,2007;李伟丰等,2007;Shi et al.,2010),但目前国内外涉及桔小实蝇迁飞行为发生机理方面的相关研究甚少。明确桔小实蝇迁飞过程中的能量利用与物质代谢是阐述桔小实蝇迁飞机制的基础,而与此相关的研究尚未见报道。
昆虫的迁飞伴随着较高的代谢速率和能源物质的消耗,主要的能源物质一般有碳水化合物、脂类化合物、氨基酸等其中一种或两种以上(Rankin,1992)。甜菜夜蛾 Spodoptera exigua(Hübner)属于糖类和脂肪都能利用的混合型,小桕天蛾Philosamia cynthia主要利用脂肪,而红头丽蝇Calliphora erythrocephala及大多数双翅目昆虫主要利用糖类为飞行提供能源 (Beenakkers,1969;李克斌等,2005)。桔小实蝇为双翅目昆虫,基于此,本文拟通过测定桔小实蝇不同日龄干重和糖原的累积及最强飞行日龄不同吊飞时间处理的雌、雄成虫糖原的含量和体重消耗,为揭示桔小实蝇飞行与能源消耗的相关关系提供基础数据,为桔小实蝇的检疫、监测预警和防治提供科学依据。
1 材料与方法
1.1 供试虫源
试验所用虫源均为云南省林业科学院森林保护研究所饲养的桔小实蝇种群,经室内连续继代饲养约10代的成虫。饲养条件为:温度25℃ -28℃,湿度为60% -80%,光周期为 L16∶D8。同一时期羽化的成虫聚养在同一养虫笼 (75.0 cm×60.0 cm×45.0 cm)内,每24 h补充一次饲料和饮用水。
1.2 试验处理及虫体取样
分别取1、4、7、10、15、20、25日龄的桔小实蝇雌、雄成虫各12头,用乙醚快速麻醉后,于烘箱中80℃烘烤12 h至恒重,称单头干重,然后放入装有变色硅胶的1.5 mL离心管中,于-20℃的冰箱中保存备用。
取最强飞行日龄15日龄 (袁瑞玲等,2014)的桔小实蝇,用乙醚轻微麻醉,在飞行磨 (即昆虫飞行数据微机采集系统)吊臂的一端直接粘取少量市售502胶水,小心的粘于成虫的前胸背板上,确认桔小实蝇头胸部、腹部和翅基部未受到影响,将带有桔小实蝇的吊臂放回飞行磨上待测,吊臂的另一端与吊虫端需保持水平。温度25℃-28℃,湿度为60% -80%,光照为自然光照,晚间补给光照,吊飞时间分别为1、2、5、10和24 h,整个飞行过程中不补充水分和营养。每处理雌、雄成虫各12头。计算机系统记录飞行时间、距离等参数。设同一日龄的不飞行个体为对照。吊飞结束后,于烘箱中80℃烘烤12 h,称干重,然后放入装有变色硅胶的1.5 mL离心管中,于-20℃的冰箱中保存备用。
1.3 飞行前后体重的测定
15日龄成虫吊飞测试前,分别测量雌、雄成虫单头虫重,吊飞结束后,分别对测试个体进行称重,二者体重相减,即可得到测试个体的体重消耗量。测试个体的体重消耗量与吊飞测试前该测试个体的单头虫重之比×100,即可得到测试个体的体重消耗百分比。
体重消耗百分比 (%)= 〔(吊飞前体重-吊飞后体重)/吊飞前体重〕 ×100
1.4 虫体糖原含量的测定
将烘干后去掉头、足、翅的单头虫体放入1.5 mL离心管中,捣碎,加入30%的KOH 400μL,振荡器上混匀,沸水浴30-40 min后取出,立即加入无水乙醇900μL混匀,离心 (12000 r/min,15 min)后,弃上清液,往沉淀物中加入1 mL 0.5 MHcl。沸水浴2 h后过滤,用10 mL离心管定容至3 mL,取1 mL,用改进的蒽酮法测糖原含量。将所得数值乘0.927(李克斌和罗礼智,1998),即为糖原值 (单位mg/g)。
改进的蒽酮法:对照取1 mL蒸馏水,待测样品各取1 mL,分别加到玻璃试管中,每支试管用移液管分别加入5 mL 0.2%蒽酮溶液;按下秒表计时,摇匀,反应70 S,用冰水迅速冷却后,室温平衡,在UV1100II单光束紫外可见分光光度计(UV1100II Series UV-VIS Spectrophotometer)上,于620 nm测定吸光度。
1.5 数据分析处理
所获数据用SPSS17.0统计软件进行方差分析,对各处理间方差进行齐性Levene检验,各组方差齐时 (P>0.05)或组间方差不能达到齐性,但对数据进行转换后,组间方差齐时,经正态检验后,用Duncans新复极差法进行多重比较;组间方差经数据转换后仍不齐时 (P<0.05),采用Games-Howell法进行多重比较。
其中,不同日龄桔小实蝇的雌雄虫干重方差齐 (P>0.05);桔小实蝇雌雄虫不同日龄糖原含量数据经对数转换;吊飞测试中雌虫体重消耗数据经对数转换,雄虫体重消耗数据经反正弦平方根转换;其他试验数据方差不齐 (P<0.05)。
2 结果与分析
2.1 虫体干重与虫龄的关系
不同日龄桔小实蝇的成虫干重有一定程度的差异,并随着日龄的增加其虫体干重呈现先增加后下降的现象 (图1)。雌、雄虫相比较,1日龄雌、雄虫干重没有差异,且为最低值,平均单头重量为2.47 mg,极显著低于其他日龄 (P<0.01);其他日龄的雌虫干重均高于雄虫。在雄虫中,7日龄的干重最高,达4.33 mg/头,显著高于除4、20、25日龄以外的其他日龄 (P<0.05)。就雌虫而言,20日龄的干重最高,达5.89 mg/头,显著高于其他日龄 (P<0.05),是1日龄雌虫的2.4倍,其中4、7、10、15和25日龄之间差异不明显 (P >0.05)。

图1 不同日龄桔小实蝇干重的变化Fig.1 Changes of dry body weight of Bactrocera dorsalis(Hendel)at different days after emergence
2.2 不同日龄虫体糖原含量的动态
不同日龄桔小实蝇体内糖原的含量变化见图2。1日龄虫糖原含量最低,随日龄的增加,雄虫糖原的含量逐渐增加,至10日龄达到最高值42.82 mg/g,其后含量逐渐降低,7、10、15、20日龄糖原含量没有显著差异,但极显著高于1、4和25日龄 (P<0.01)。就雌虫而言,糖原含量先随日龄的增加而增加,在7日龄时达到较高值,其后糖原含量在4日龄水平上下波动,15日龄含量最高,达29.77 mg/g,显著高于除7日龄外的其他日龄(P<0.05),是1日龄虫含量的4.6倍。雌雄虫相比较,除25日龄外,各个日龄单位重量的雄虫糖原含量略高于雌虫。

图2 不同日龄桔小实蝇糖原含量的变化Fig.2 Changes of glycogen at different days after emergence in Bactrocera dorsalis(Hendel)
2.3 飞行对桔小实蝇体重的消耗动态
吊飞处理的测定结果表明,桔小实蝇飞行大量消耗体重,吊飞2 h后的雌雄虫体重均显著低于对照 (P<0.05)。由图3可以看出,桔小实蝇飞行过程中体重消耗百分比Y(%)随吊飞时间x(h)的延长而增加,两者呈直线相关,雌虫:Y♀=1.2899x+5.5696(r=0.9961);雄虫:Y♂=1.8550x+3.2210(r=0.9962)。但实际飞行时间与吊飞时间往往不等,因为在飞行的过程中桔小实蝇会自行停歇。吊测的桔小实蝇实际飞行时间、飞行距离及体重消耗情况见表1。
分析实际飞行时间、距离和体重消耗三者间的关系,得到以下直线模型。体重消耗Y(%)依飞行时间t(h)的变化方程为:

体重消耗Y(%)与飞行距离D(km)的关系式为:

飞行距离D(km)依飞行时间t(h)的变化方程为:

由表1还可以看出,相同的吊飞时间内,雄虫实际飞行的时间和距离较雌虫长 (远),从而表现为雄虫体重消耗高于雌虫 (图3),而事实是,单位实际飞行时间 (h)和单位飞行距离 (km)的雌虫体重消耗均高于雄虫,其原因是雌虫个体较雄虫大体重较重。

表1 不同吊测时间处理的桔小实蝇成虫实际飞行时间、距离及其体重消耗Table 1 The actual flight duration,distance and weight consumptions of adults with different tethered time
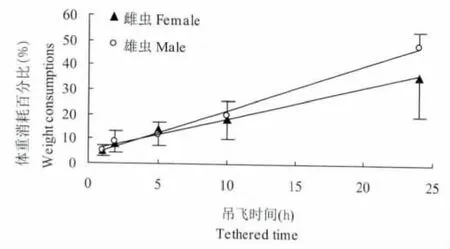
图3 桔小实蝇飞行过程中体重消耗与处理时间的关系Fig.3 The relation between tathered time and weight consumption during Bactrocera dorsalis(Hendel)migration
2.4 桔小实蝇飞行过程中糖原的消耗
桔小实蝇吊飞过程中糖原含量的变化见图4。结果表明,糖原含量和飞行时间呈显著的负相关。吊飞前糖原的含量雌虫为29.77 mg/g,雄虫为35.1 mg/g,吊飞24 h后分别降低至3.67 mg/g和3.95 mg/g。方差分析结果表明,吊飞2 h,糖原含量与对照间即达到显著差异 (P<0.05),吊飞5 h达到极显著差异 (P<0.01),随着飞行时间的增加,糖原含量显著下降。表明糖原是桔小实蝇飞行的能源物质之一。
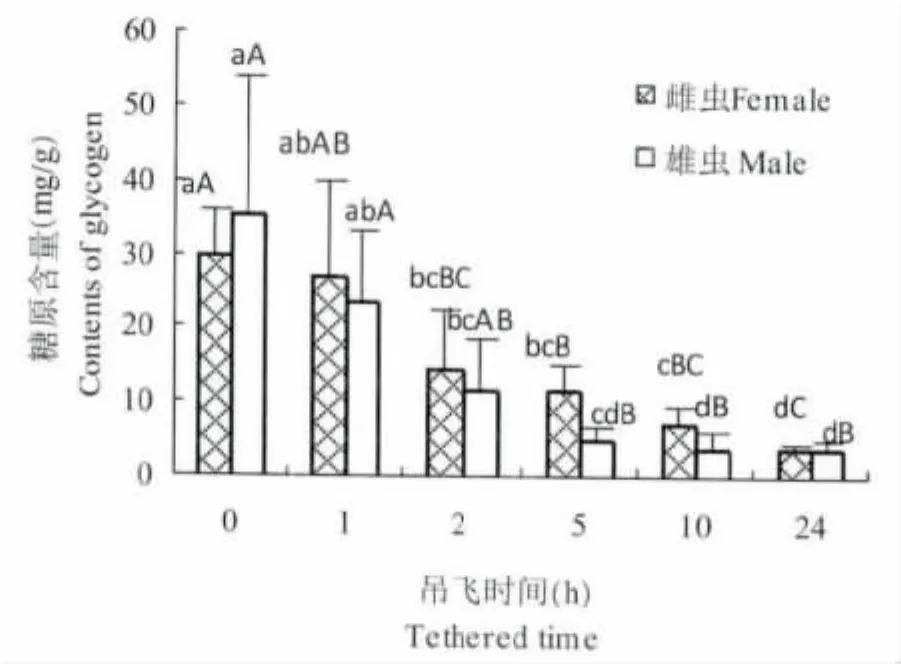
图4 桔小实蝇成虫吊飞处理的虫体内糖原含量 (mg/g)Fig.4 The glycogen contents of adults with different tethered time
飞行过程中糖原消耗量G(mg/g)依飞行时间t(h)的变化模型为Logistic模型:

同理亦可建立糖原消耗依飞行距离的变化模型。糖原消耗与实际飞行时间和飞行距离呈正相关,与用于飞行的体重消耗关系一致。比较糖原消耗和体重消耗 (图5、6),雌虫 (图5),随着体重消耗的增加,糖原消耗占虫体重消耗率也在增加,虽然吊飞10、24 h时糖原消耗占虫体重消耗的百分比有所降低,但从体重消耗看,吊飞24 h比5 h高出2.7倍,而糖原消耗占虫体重消耗率,吊飞5 h比24 h,仅高1.5倍,表明糖原消耗随吊飞时间的延长持续增加,证明雌虫飞行消耗糖原。雄虫的糖原消耗占虫体重消耗率的变化同雌虫相似,说明糖原是桔小实蝇飞行的能源物质之一。

图5 桔小实蝇雌虫不同吊飞时间内的体重消耗与糖原消耗比较Fig.5 The comparision between weight consumptions and glycogen consumptions of female Bactrocera dorsalis(Hendel)with different tethered time
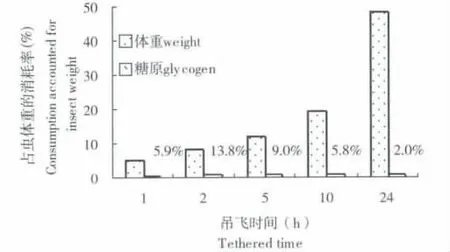
图6 桔小实蝇雄虫不同吊飞时间内的体重消耗与糖原消耗比较Fig.6 The comparision between weight consumptions and glycogen consumptions of male Bactrocera dorsalis(Hendel)with different tethered time
分析糖原的消耗速率发现,随飞行活动的增加,虽然糖原的消耗与实际飞行时间和飞行距离呈正相关,但速率呈现先增加后逐渐减缓的趋势,单位实际飞行时间糖原消耗,雌虫为16.53-47.05-24.45 mg/(h·g),雄虫为39.43-67.63-9.48 mg/(h·g);单位飞行距离糖原消耗,雌虫为10.25-34.09-20.47 mg/(km·g),雄虫为18.70-42.95-7.36 mg/(km·g)。由此看出,飞行的初期,雄虫所消耗的糖原速率高于雌虫,随着飞行时间和距离的增加雌虫消耗糖原的速率高于雄虫。
3 结论与讨论
国内外研究表明,桔小实蝇具有远距离迁飞能力。Steiner(1957)在美国夏威夷,通过标记重捕方法研究发现,桔小实蝇迁飞距离达37 km。Iwahashi(1972)在日本小笠原群岛研究发现,桔小实蝇可飞越50 km的海面。朱耀沂和邱辉宗(1989)基于标记释放试验发现,桔小实蝇可从台湾本岛随东北风飞到27 km以外的小琉球岛。Froerer et al.(2010)基于标记重捕法研究发现,在美国夏威夷岛上的高山区,桔小实蝇迁飞距离达2-11.39 km。在中国西南纵向岭谷区桔小实蝇迁移扩散距离可达97 km(陈鹏等,2007)。梁帆等(2001)应用昆虫飞行磨初步测试发现广州地区当地的桔小实蝇一次飞行最远距离可达46.16 km,证明桔小实蝇可远距离迁飞。本试验吊飞24 h的测试中,雌虫平均飞行距离为1.27 km,变幅为0.33-3.35 km,雄虫平均飞行距离为4.23 km,变幅为0.001-10.16 km,最远飞行距离与Froerer et al.(2010)研究结果较为一致,在夏威夷岛桔小实蝇迁飞距离在2-11.39 km之间,再次试证了桔小实蝇具有较强的自主飞行能力。本研究与上述其他研究中桔小实蝇的飞行能力存在差异的原因,很可能是由室内试虫与野外桔小实蝇自身的差异引起。室内饲养的桔小实蝇种群,继代饲养约10代左右,其生理、生物学等特性与野生种群存在差异;吊飞是在静风、被动悬吊的室内条件,且供试昆虫未进行补充营养,使室内吊飞测试的桔小实蝇飞行距离低于野外种群。
能源贮备是昆虫飞行的物质基础,本研究测定了不同日龄桔小实蝇干重和糖原的累积动态,其总体表现为随日龄的增加,干重和糖原先增加后下降。对桔小实蝇飞行能力的测试 (袁瑞玲等,2014)和卵巢解剖 (陈敏等,2014)发现,羽化初期的成虫,翅较软,飞行肌发育不完全,远距离飞行能力较弱。随着日龄的增加,虫体内能源物质不断积累,飞行能力也逐渐增强。4日龄后雌虫卵巢开始发育;一般到5日龄时,成虫开始出现交配现象;7日龄时出现交配高峰,并有少量个体开始产卵;10日龄时,桔小实蝇成虫已进入产卵期,表现出第一个飞行能力的高峰;15日龄飞行能力最强,随后桔小实蝇会继续补充营养,积累能源物质继续交配、产卵;19-20日龄时达到产卵高峰。桔小实蝇雌虫干重在20日龄,糖原含量在15日龄达最高,说明桔小实蝇干重和糖原的贮备与生殖及飞行能力间存在明显的相关性。其体内糖原的累积并不随性器官的发育成熟而降低,而是随日龄的增加在强飞行能力期间 (10-25日龄),同时也是交配产卵期7-20日龄内达到最高,表明桔小实蝇不存在多数迁飞性昆虫所特有的“卵子发生与飞行共轭”现象。
研究中发现,不同日龄桔小实蝇体内糖原的累积量和吊飞过程中糖原的消耗量几乎都存在雄虫高于雌虫的现象,这与迁徙蚱蜢Melanoplus sanguinipes雌雄个体飞行相同时间,雄性个体消耗的能源物质远高于雌性个体 (Rankin,1992);粘虫飞行初期雄蛾动用海藻糖的比例要比雌蛾大得多,而在适宜的温、湿度条件下,飞行同样的时间雄蛾消耗的能源物质也比雌蛾多 (王宗舜和欧阳迎春,1995)的研究结果一致。这种性别差异导致的能源物质储存、利用率的不同,很可能是因为雄虫为了尽快的寻找到合适的雌虫繁衍下一代,更早的运用了能源物质,而雌虫,交配后还需寻找后代更好更理想的生存场所,其所释放的能源物质相比雄虫会缓慢一些。也有报道认为可能是由于不同性别昆虫酶活性的差异所致 (李克斌和罗礼智,1999),其根本原因、生理机制有待研究。
糖类和脂肪被认为是昆虫飞行的两类主要能源物质。糖类因其在血淋巴中有较高的溶解度能够被迅速动用,在昆虫的起飞阶段起重要作用。在某些昆虫中糖类是唯一的可以利用的能源物质,其中包括大多数的双翅目昆虫,膜翅目昆虫及某些鳞翅目昆虫 (Steele,1981;Wheeler,1989)。而其它的一些迁飞昆虫,在飞行的起始阶段利用糖类,而持续飞行过程则转向利用脂类 (Beenakkers et al.,1984)。飞蝗在持续飞翔期间维持使用糖类作为飞翔肌的能源,是飞翔肌提供消耗能量的1/4,其余的3/4消耗能量由脂类提供 (van der Horst et al.,1978)。棉铃虫在整个飞行期间糖原、脂肪提供的能量占总能量消耗的14.12%和85.87%(吴孔明和郭予元,1998)。甜菜夜蛾在初始阶段主要利用糖类作为能源物质,而后期飞行又可通过脂类代谢提供能量 (韩兰芝等,2005)。糖原是糖类贮存的主要形式之一。本研究结果表明,桔小实蝇在整个吊飞期间,糖原一直处于动用状态,表明糖原是桔小实蝇飞行的能源物质之一。而不同吊飞时间的桔小实蝇糖原消耗占体重消耗率在14%以下,这就说明,糖原不可能是桔小实蝇长时间 (远距离)飞行的唯一能源物质,其飞行的能源物质应该还有其他种类。飞行过程中脂肪是否作为能源物质提供能量?其供能占能量消耗的比例如何?还需要进一步研究。
根据试验结果拟合的多个数学模型直观的体现了桔小实蝇成虫体重变化、糖原消耗与飞行时间和距离之间的相互关系,为阐明桔小实蝇远距离飞行的能量利用与物质代谢的关系提供了科学依据,为桔小实蝇的综合防控提供了基础信息。
References)
Arévalo - Galarza L, Follett PA. Response of Ceratitis capitata,Bactroceradorsalis,and Bactrocera cucurbitae(Diptera:Tephritidae)to metabolic stress disinfection and disinfestation treatment[J].Journal of Economic Entomology,2011,104(1):75 -80.
Beenakkers AMT.Carbohydrate and fat as a fuel for insect flight:a comparative study[J].Journal of Insect Physiology,1969,15:353-361.
Beenakkers AMT,van der Horst DJ,van Marrewijk WJA.Insect flight muscle metabolism[J].Insect Biochem.,1984,14:243-260.
Chu YI,Chiu HT.The re-establishment of Dacus dorsalis Hendel(Diptera:Trypetidae)after the eradication on Lambay Island [J].Chin.J.Entomol.,1989,9:217-230.[朱耀沂,邱辉宗.小琉球东方果实蝇灭绝处理后再发生为害原因之探讨[J].中华昆虫,1989,9:217-230]
Chen M,Chen P, Ye H,et al. Morphological structures and developmental characteristics of the ovaries of Bactrocera dorsalis[J].Journal of Environmental Entomology,2014,36(1):83 -88.[陈敏,陈鹏,叶辉,等.桔小实蝇卵巢形态结构及发育特征研究 [J].环境昆虫学报,2014,36(1):83-88]
Chen P,Ye H.Population dynamics of Bactrocera dorsalis(Diptera:Tephritidae)along with analysis on the factors influencing the population in Baoshanba,Yunnan,China [J].Entomological Science,2007,10(2):141-147.
Chen P,Ye H,Mu QA.Migration and dispersal of the oriental fruit fly,Bactrocera dorsalis in regions of nu jiang river based on fluorescence mark[J].Acta Ecologica Sinica,2007,27(6):2468 -2476.[陈鹏,叶辉,母其爱.基于荧光标记的怒江流域桔小实蝇(Bactrocera dorsalis)的迁移扩散 [J].生态学报,2007,27(6):2468 -2476]
Chen P,Ye H.Relationship among five populations of Bactrocera dorsalis based on mitochondrial DNA sequences in western Yunnan,China[J].Journal of Applied Entomology,2008,132(7):530 -537.
Drew RAI,Hancock DL.The Bactrocera dorsalis complex of fruit flies(Diptera:Tephritidae:Dacinae)in Asia [J]. Bulletin of Entomological Research,1994(Suppl.2):1-68.
Han LZ,Zhai BP,Zhang XX,et al.Activity of enzymes related to energy metabolism in the flight muscle of beet armyworm[J].Acta Ecologica Sinica,2005,25(5):1101-1106.[韩兰芝,翟保平,张孝羲,等.甜菜夜蛾飞行肌中与能量代谢有关的酶活性[J].生态学报,2005,25(5):1101-1106]
Iwahashi O.Movement of the oriental fruit fly adults among islets of the Ogasawara Islands[J].Environmental Entomology,1972,1:176-179.
Froerer KM,Peck SL,McQuate GT,et al.Long-distance movement of Bactrocera dorsalis(Diptera:Tephritidae)in Puna,Hawaii:How far can they go?[J].American Entomologist,2010,56(2):88-94.
Li KB,Luo LZ.Effects of larval rearing density on energetic reserves in the adults of oriental armyworm Mythimna Separata(Walker)[J].Acta Entomol.Sin.,1998,41(3):250-257.[李克斌,罗礼智.粘虫幼虫密度对成虫能源物质含量的影响[J].昆虫学报,1998,41(3):250-257]
Li KB,Luo LZ.Activities of enzymes in the flight muscle of pupal and adult oriental armyworm,Mythimna separata(Walker)[J].Acta Entomol.Sin.,1999,42(1):37-43.[李克斌,罗礼智.粘虫飞行肌中与能量代谢有关的酶活性研究 [J].昆虫学报,1999,42(1):37-43]
Liang F,Wu JJ,Liang GQ.The first reportof the teston the flight ability oforiental fruit fly[J].Acta Agricultureof University Jiangxi,2001,23(2):259-260.[梁帆,吴佳教,梁广勤.桔小实蝇飞行能力测定试验初报[J].江西农业大学学报,2001,23(2):259-260]
Li KB,Gao XW,Cao YZ,et al.Dynamics of energy reserves and utilization after tethered-flight in the beet armyworm,Spodoptera exigua(Hübner)(Lepidoptera:Noctuidae) [J]. Acta Phytophylacica Sinica,2005,32(1):13-17.[李克斌,高希武,曹雅忠,等.甜菜夜蛾能源物质积累及其飞行能耗与动态[J].植物保护学报,2005,32(1):13-17]
Li WF,Yang L,Tang K,et al.Microsatellite polymorphism of Bactrocera dorsalis(Hendel)populations in China[J] .Acta Entomol.Sin.,2007,50(12):1255-1262.[李伟丰,杨朗,唐侃,等.中国桔小实蝇种群的微卫星多态性分析[J].昆虫学报,2007,50(12):1255-1262]
Rankin MA.The cost of migration in insect[J].Annu.Rev.Entomol.,1992,37:533-559.
Steiner LF.Field evaluation of oriental fruit fly insecticides in Hawaii[J].Journal of Economic Entomology,1957,50:16-24.
Steele JE. The Role of Carbohydrate Metabolism in Physiological Function in Energy Metabolism in Inseets[M].New York:Liss Press,l981,101-134.
Shi W,KerdelhuéC,Ye H.Population genetic structure of the oriental fruit fly,Bactrocera dorsalis(Hendel)(Diptera:Tephritidae)from Yunnan province(China)and nearby sites across the border[J].Genetica,2010,138(3):377-385.
Suganya R,Chen SL,Lu KH.Target of rapamycin in the oriental fruit fly Bactrocera dorsalis(Hendel):Its cloning and effect on yolk protein expression [J]. Archives of Insect Biochemistry and Physiology,2010,75:45-56.
van der Horst DF,van Doorn JM,Beenakkers AMT.Dynamics in the haemolymph trehalose pool during flight of the locust Locusta migratoria[J].Insect Biochem.,1978,8:413-416.
Wheeler CH.Mobilization and Transport of Fuels to the Flight Museles.In Insect Flight[M].Boca Raton.FL.:CRC Press,1989,273-303.
Wang ZS,Ouyang YC.Carbohydrates mobilization and utilization during initial flight period in the moths Mythimna separata(Walker)[J].Acta Entomol.Sin.,1995,38(2):146-152.[王宗舜,欧阳迎春.东方粘虫飞行初期糖类的动用和消耗[J].昆虫学报,1995,38(2):146-152]
Wu KM,Guo YY.The relationship between utilization of energy substances and sustained flight in cotton bollworm,Helicoverpa armigera[J].Acta Entomol.Sin.,1998,41(1):15-19.[吴孔明,郭予元.棉铃虫飞翔的能源物质及消耗[J].昆虫学报,1998,41(1):15-19]
Yuan RL,Yang S,Wang XX,et al.Test on flight ability of Bactrocera dorsalis(Hendel)[J].Journal of West China Forestry Science,2014,43(6):66-71.[袁瑞玲,杨珊,王晓谓,等.桔小实蝇飞行能力测试[J].西部林业科学,2014,43(6):66-71]
猜你喜欢
热带生物学报(2022年6期)2022-11-29
甘肃农业科技(2021年6期)2021-07-25
环境昆虫学报(2020年5期)2020-11-11
科学导报(2019年35期)2019-09-03
西北农林科技大学学报(自然科学版)(2018年5期)2018-05-24
新农村(2018年32期)2018-04-02
湖南农业(2016年3期)2016-06-05
湖南农业(2016年3期)2016-06-05
湖南农业(2015年5期)2015-02-26
江苏农业科学(2014年3期)2014-07-16
