
淮北地区5种不同植物群落土壤呼吸研究
2015-11-07 01:31:14鲁绍伟李少宁
水土保持研究 2015年4期
刘 斌, 鲁绍伟, 石 媛, 李少宁, 陈 波
(1.河北农业大学 林学院, 河北 保定 071000; 2.北京市农林科学院 林业果树研究所, 北京 100093)
淮北地区5种不同植物群落土壤呼吸研究
刘 斌1,2, 鲁绍伟2, 石 媛1, 李少宁2, 陈 波2
(1.河北农业大学 林学院, 河北 保定 071000; 2.北京市农林科学院 林业果树研究所, 北京 100093)
选取河南西平杨树人工林生态系统中5种不同植物群落土地为研究对象,利用Licor-8100土壤呼吸测定系统对其土壤呼吸进行了连续观测,分析、比较5种不同植物群落土壤呼吸的差异及其影响因素。研究结果表明:(1) 5种植物群落下土壤呼吸有所差异,但其日变化及年内变化趋势大致相同,均与土壤温度变化趋势一致,呈单峰曲线,其土壤呼吸大小顺序为:8 m行宽林间小藜—牛筋草地>2 m行宽林间苍耳地>8 m行宽林间水蓼—灰灰菜地>8 m行宽林间苍耳地>2 m行宽林间裸地;(2) 采用土壤温度和湿度单因素模型(Rs=aebTs或Rs=aW+b)进行分析:土壤温度和土壤含水量分别解释了不同植物群落土壤呼吸季节变化的41%~79%和2.3%~21%;而采用双因素模型分析(Rs=aebTsWc),土壤温湿度共同解释了土壤呼吸速率季节变化的49.1%~86.7%,表明不同植物群落的土壤呼吸均受土壤温度和土壤含水量的共同影响;(3) 运用Q10=e10b模型分析,Q10值大小顺序为:8 m行宽林间小藜—牛筋草地(2.47)>2 m行宽林间苍耳地(2.3) >2 m行宽林间裸地(2.7) >8 m行宽林间水蓼—灰灰菜地(2.59)>8 m行宽林间苍耳地(2.22),且5种不同植物群落下土壤呼吸Q10值均表现为春季>冬季>秋季>夏季,即土壤呼吸温度敏感性随着温度的升高呈现出降低的趋势。
土壤呼吸; 不同植物群落;Q10值; 土壤温度
土壤呼吸是全球碳循环中一个重要的过程[1],全球每年因土壤呼吸向大气中释放的碳的估算值约为75~120 Pg,仅次于全球陆地总初级生产力(GPP)的估算值[2]。土壤呼吸指土壤产生和向大气释放CO2的过程,包括土壤微生物呼吸、植物根系呼吸、土壤动物呼吸和含碳物质的化学氧化作用等几个生物学和非生物学部分[3-4]。土壤呼吸过程是陆地生态系统的碳循环中土壤碳的主要输出途径,是大气CO2重要排放源,其排放量是陆地生态系统向大气排放碳的最大通量之一[5]。土壤呼吸是一个受生物和非生物因素控制的复杂过程,具有很大的空间和时间变异性。不同的生态系统因为土壤温度、土壤含水量、植物群落及土壤质地的不同,所受到的影响也不尽相同,造成其土壤呼吸有所差异[6-9]。土壤温度和土壤含水量是土壤呼吸的重要影响因素,评估土壤呼吸的影响因素需要量化土壤温度及土壤含水量对土壤呼吸的影响,并了解植物群落对土壤呼吸的影响[10-11]。土壤呼吸与土壤温度之间往往呈指数相关,土壤呼吸对温度的敏感程度通常用Q10值来反映[12-13],目前对土壤呼吸Q10值的研究已经较为普遍。
不同的植物群落不仅改变了地表植被,也改变了土壤透气性,从而使土壤有机质含量、微生物组成和活性、根系生物量等发生改变,相应的土壤呼吸也就不相同。研究不同植物群落土壤呼吸速率及时空变化特征,阐明其影响因子和调控机制是目前生态学研究的重点内容之一。目前对于生态系统碳循环的研究已经十分全面,且对于不同林分和生物群落的对比研究也逐渐增多,但就同一生态系统内不同植物群落的研究相对较少。本文以河南西平县杨树人工林生态系统为研究对象,对不同植物群落的土壤呼吸进行连续观测,探讨土壤呼吸的日变化、年变化特征,并分析对比不同植物群落之间土壤呼吸的差异及其影响因素为科学合理的评估我国温度森林地区土壤碳库动态提供有力的支撑。
1 研究区概况
淮北平原农林复合生态系统定位研究站位于河南省西平县东,地处淮河流域上游,地理坐标为东经113°36′—114°32′,北纬32°58′—33°24′,是北亚热带和暖温带的过渡区,属湿润大陆性季风气候。试验地位于研究站110 hm2的杨树人工林内,林内树种以欧美杨为主,树龄为7 a,其栽植年份一致,且处于相同的立地条件下,植株生长均匀,林分种类单一,1.5 m株距,2 m行距,每两行间隔8 m,林下植被以苍耳(XanthiumsibiricumPatrin)为主,还有黄鹌菜(Youngiajaponica)、灰灰菜(ChenopodiumalbumLinn.)、小藜(ChenopodiumserotinumL)、水蓼(Polygonumhydropiper)、牛筋草(Eleusineindica(L.) Gaernt)、毛马唐(Digitariaciliaris(Retz.) Koel)、鳢肠(EcliptaprostrataL)等,种类不多,且生长量小。整体上植被种类稀少,生态系统结构简单,抗逆性弱。林下植物的根系主要集中在0—30 cm,60 cm以下无根系分布。
2 试验设计和研究方法
2.1试验方法
在试验地内选取总样地面积为7 200 m2,规格为120 m×60 m,根据人工林的种植配置方式,在典型植被条件下各布设一个测量点,共布设5个测量点。该林地有裸地,有低矮草地,有长势非常好的高约1 m的草本植物,由于杨树的配置方式为1.5 m株距,2 m行距,每两行间隔8 m,有宽林间地和窄林间地,各点的林下植被种类分别为:1号2 m行宽林间裸地、2号8 m行宽林间水蓼、灰灰菜地、3号8 m行宽林间苍耳地、4号2 m行宽林间苍耳地和5号8 m行宽林间小藜、牛筋草地。1号裸地地表有少量的枯落物存在;3号地和4号地的苍耳地上部分较高且非常茂盛,再没有其它植物。
在选择好各测量点后,将环刀水平放置,然后用恒力将环刀垂直缓慢打入,环刀上面用软木或朽木作为缓冲垫,以免环刀受损。打入土部分在4—6 cm为宜。在安放好之后约12 h后方可进行测定,测定时采用美国Licor-8100土壤碳通量测定仪进行定时连续测定。试验在2009年1月—12月进行,每月选取4 d(每月的1号、10号、21号、30号)时间,采用移动式气室全天24 h每隔3 h测定一次,每次测定时间为120 s,采用3次重复测定,取其平均值,并同步测定5 cm土壤温度和10 cm处土壤含水量。
2.2数据分析
目前,国内研究土壤温度和湿度对土壤呼吸的影响时往往采用单因素模型单独分析温度或湿度对土壤呼吸的影响,采用双因素模型分析二者对土壤呼吸共同影响的研究较少。为了分析温度和水分对土壤呼吸的共同影响和单独影响,本研究采取如下模型进行比较[14]:
Rs=aebTsWc;Rs=aebTs;Rs=aW+b。
式中:Rs——土壤呼吸[g/(m2·h)];Ts——5 cm处土壤温度(℃);W——10 cm处土壤含水率(m3/m3);a,b,c——待定参数。
所有数据处理及图表制作均通过SPSS 17.0和Excel 2007软件完成。
3 结果分析
3.1不同植物群落的土壤呼吸日变化
图1显示了5种不同植物群落土壤呼吸速率日变化,由图1不难看出,5种不同植物群落的土壤呼吸速率整体变化趋势是一致的,其最大值出现在11:00—14:00,最小值出现在5:00—8:00左右。其中8 m行宽林间小藜—牛筋草地与2 m行宽林间苍耳地变化趋势一致,最大值出现在11:00,其值分别为1.44 ,1.48 g/(m2·h),最小值出现在5:00,其值分别为1.23,1.34 g/(m2·h),总体来说是夜间小于白天。2 m行宽林间裸地、8 m行宽林间水蓼—灰灰菜地、8 m行宽林间苍耳地三者变化趋势较为接近,此三者的土壤呼吸值均明显低于8 m行宽林间小藜—牛筋草地和2 m行宽林间苍耳地,三者均呈现出先增加而后降低的变化趋势,土壤呼吸最大值分别为1.04,1.21,1.06 g/(m2·h),均出现在14:00;最小值分别为0.84 ,0.93 ,0.91 g/(m2·h),其中2 m行宽林间裸地与8 m行宽林间水蓼—灰灰菜地土壤呼吸最小值出现在5:00,而8 m行宽林间苍耳地略有差异,其最小值出现在8:00。
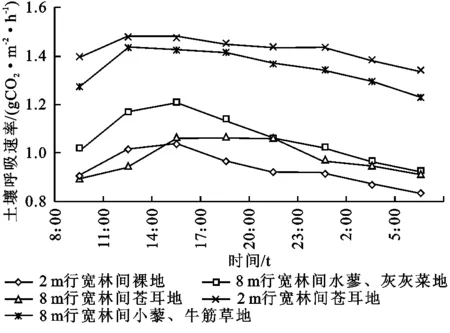
图1 不同植物群落土壤呼吸日变化
3.2不同植物群落土壤温湿度及其土壤呼吸季节变化
图2为2 m宽林间裸地、8 m行宽林间水蓼—灰灰菜地、8 m行宽林间小藜—牛筋草地、2 m行宽林间苍耳地和8 m行宽林间苍耳地5种不同植物群落下5 cm处的土壤温度(Ts)随季节变化的趋势,5种不同植物群落土壤温度的变化均呈现出先增大而后减小的趋势。其最大值均出现在7月,值分别为28.1,28.11,28.64,28.61,28.33℃;最小值均出现在2月份,其值分别为2.8,2.83,3.01,2.94,2.8℃。土壤温度的变化趋势与其对应的土壤呼吸变化趋势是一致的。图3显示了10 cm处土壤含水量在一年尺度上的变化,5种不同植物群落的土壤含水量均呈现出无规律性波动,除了6月份土壤含水量较高外,其余月份土壤含水量相差不大;土壤含水量最小值出现在3月份,其值分别为0.19,0.6,0.58,0.58,0.59 m3/m3;最大值出现在6月份,其值分别为1.89,1.88,1.85,1.82,1.88 m3/m3。6月份降雨量偏大,可能是导致土壤含水量明显大于其余月份的主要原因。土壤含水量的变化与土壤呼吸关系不是十分明显,可能是由于本研究中土壤含水量年季变化不大导致其对土壤呼吸影响不显著,或者是其影响被土壤温度及其他因素的影响所覆盖。
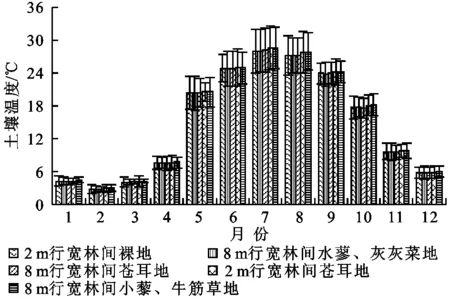
图2 不同植物群落5 cm土壤温度季节变化
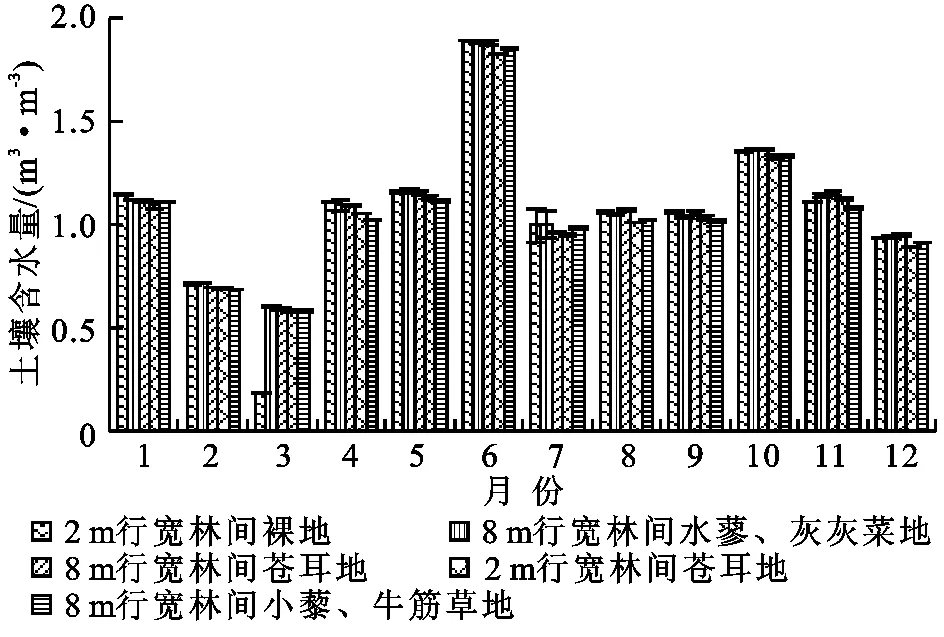
图3 不同植物群落土壤含水量季节变化
图4表明,不同植物群落的土壤呼吸速率随季节变化有所不同。8 m行宽林间小藜—牛筋草地与2 m行宽林间苍耳地变化规律基本一致,均呈明显的单峰曲线;其中8 m行宽林间小藜—牛筋草地的土壤呼吸最大值[2.96 g/(m2·h)]出现在7月,最小值[0.44 g/(m2·h)]出现在1月;2 m行宽林间苍耳地的土壤呼吸最大值[2.72 g/(m2·h)]出现在9月,最小值[g/(m2·h)]同样出现在1月。2 m行宽林间裸地、8 m行宽林间水蓼—灰灰菜地与8 m行宽林间苍耳地三者变化趋势一致,三者均呈现出先增加而后递减的趋势,但是其峰值均不明显,土壤呼吸变化幅度不大,三者土壤呼吸速率变化幅度(最大值与最小值之差除以平均值)大小顺序为2 m行宽林间裸地(139%)>8 m行宽林间水蓼—灰灰菜地(120%)>8 m行宽林间苍耳地(103%)。
不同植物群落下土壤呼吸在非生长季差距不大,而在生长季内8 m行宽林间小藜—牛筋草地、2 m行宽林间苍耳地与2 m行宽林间裸地、8 m行宽林间水蓼—灰灰菜地、8 m行宽林间苍耳地有显著差异。其中8 m行宽林间小藜—牛筋草地和2 m行宽林间苍耳地随季节变化土壤呼吸变动较大,而2 m行宽林间裸地、8 m行宽林间水蓼—灰灰菜地、8 m行宽林间苍耳地三者全年土壤呼吸变化相对较为平缓。土壤呼吸最大值均出现在7月份左右,而其最小值出现在1月份或者12月份。
3.3温湿度与土壤呼吸的关系
采用土壤温度和湿度单因素模型(Rs=aebTs或Rs=aW+b)进行分析:土壤温度和土壤含水量分别解释了不同植物群落土壤呼吸的变化的41%~79%和2.3%~21%;而采用双因素模型分析(Rs=aebTsWc),土壤温湿度共同解释了土壤呼吸速率季节变化的49.1%~86.7%,表明不同植物群落的土壤呼吸受土壤温度和土壤含水量的共同影响,均对土壤温度的变化更为敏感(表1)。
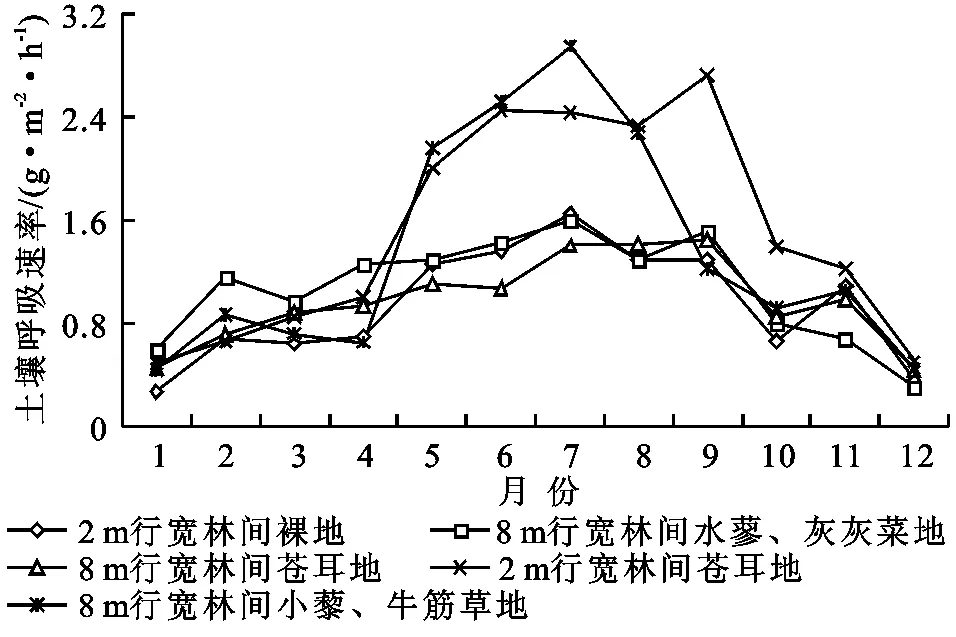
图4 不同植物群落土壤呼吸季节变化
表1中的Rs=aebTsWc模型参数b(温度敏感性因子)大小关系为:8 m行宽林间小藜—牛筋草地>2 m行宽林间苍耳地>2 m行宽林间裸地>8 m行宽林间苍耳地>8 m行宽林间水蓼—灰灰菜地,这表明8 m行宽林间小藜—牛筋草地的土壤呼吸对土壤温度最为敏感,其次为2 m行宽林间苍耳地;而模型参数c的大小关系为:8 m行宽林间小藜—牛筋草地>2 m行宽林间苍耳地>2 m行宽林间裸地>8 m行宽林间水蓼—灰灰菜地>8 m行宽林间苍耳地,表明8 m行宽林间小藜—牛筋草地土壤呼吸对土壤含水量最为敏感,其中2 m行宽林间裸地、8 m行宽林间水蓼—灰灰菜地、8 m行宽林间苍耳地的模型参数c为负值,表明这三种植物群落的土壤呼吸与土壤含水量呈负相关,即随着土壤含水量的增加,土壤呼吸呈降低的趋势。这些差异的出现是不同植物群落下土壤结构、土壤微生物均有所不同所导致的[15-16]。
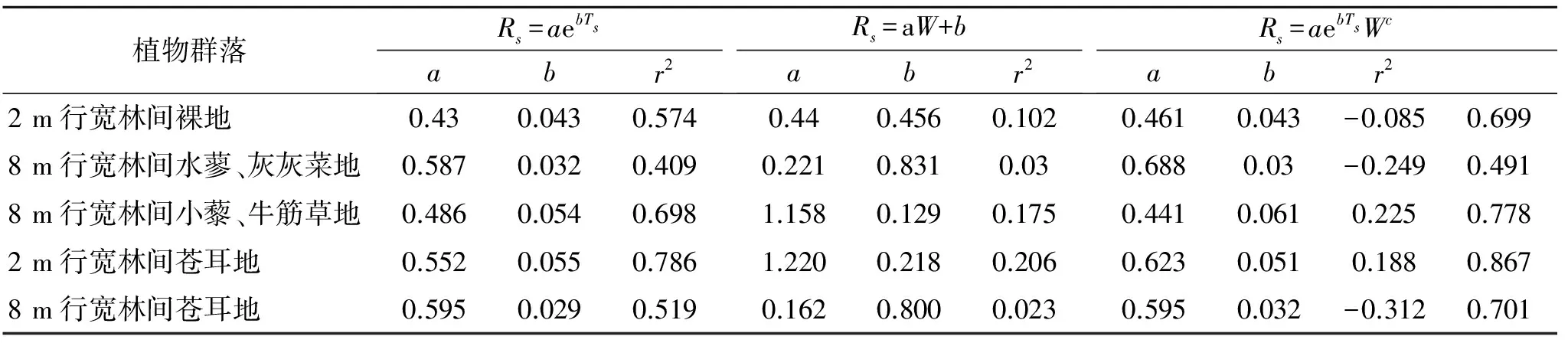
表1 土壤呼吸速率(Rs)与土壤温度(Ts)和土壤含水量(W)不同关系模型参数
3.4土壤呼吸Q10值
为探讨土壤呼吸温度敏感性大小(Q10)的影响因素,采用指数模型Rs=aebTs拟合土壤呼吸速率(Rs)与土壤温度(Ts)之间的关系。由于各地区环境因素的差异,造成对土壤呼吸速率的影响不同,计算Q10值的数学模型也不尽相同,本文采用目前研究较为常见的Q10=e10b模型(式中,b为土壤呼吸与温度指数模型Rs=aebTs中的温度反应系数)计算土壤呼吸Q10值,结果如表2所示。5种不同植物群落的土壤呼吸Q10平均值分别为2.47±0.89,2.3±0.99,2.7±1.35,2.59±1.24,2.22±0.94。8 m行宽林间小藜—牛筋草地Q10值最大,说明其土壤呼吸对温度最为敏感。5种不同植物群落的Q10有明显的季节变化,均呈现出春季>冬季>秋季>夏季的变化趋势,说明在温度偏低时土壤呼吸对温度较为敏感,而在温度较高的季节土壤呼吸对温度的敏感性呈下降趋势。
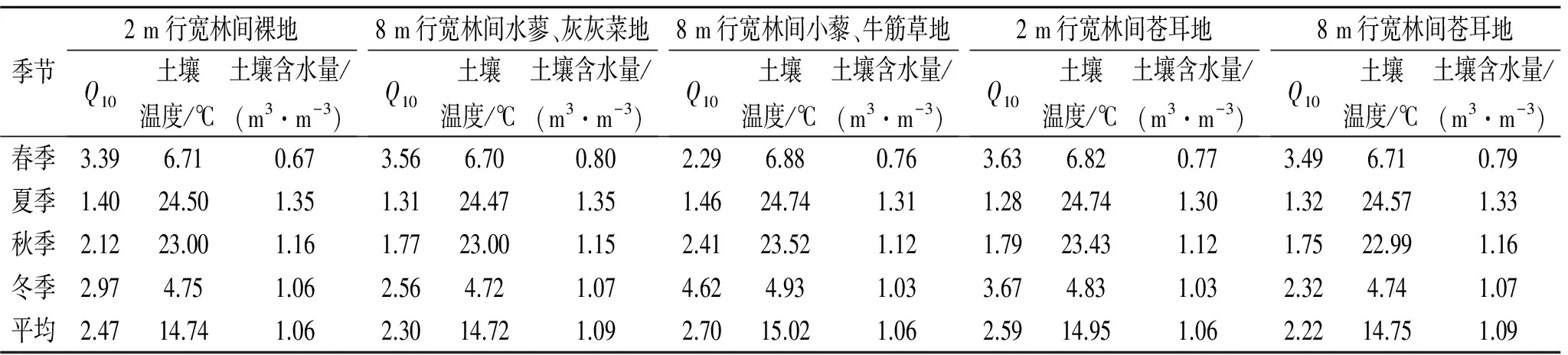
表2 不同植物群落土壤呼吸Q10值
4 讨 论
4.1不同植物群落下的土壤呼吸
5种不同植物群落的土壤温度在一天之中的变化趋势相同,其土壤呼吸与土壤温度变化趋势也大致相同,均呈现先增大而后减小的变化趋势。其最大值均出现在11:00—14:00左右,最小值出现在5:00—8:00左右。有研究表明,土壤呼吸具有明显的昼夜变化格局,黄承才等[17]对中亚热带青冈常绿阔叶林、毛竹和茶园的土壤呼吸日动态研究表明,土壤呼吸峰值出现在12:00—14:00,清晨和傍晚较低,白天逐渐升高;乌俊杰等[15]对西双版纳山地古树茶园、台地茶园和次生林的旱季土壤呼吸的研究表明,3种植物群落下的土壤呼吸有所差异,土壤呼吸最高值大多出现在12:00—16:00,这些研究结果与本研究所得结论基本一致。
研究结果表明,不同植物群落的土壤呼吸具有明显的季节变化特征,土壤呼吸在温度较高的夏季明显高于较为寒冷的冬季。董彬等[18]通过对鲁东南杨树人工林土壤呼吸的研究得出,杨树人工林土壤呼吸具有明显的季节变化趋势,夏季土壤呼吸较高,在7月份达到峰值,冬季土壤呼吸较低,在1月份达到低谷;YOU等[19]对中国东北地区落叶松人工林土壤二氧化碳释放量时间格局研究表明,土壤呼吸最大值出现在7月末或者8月份中旬,最小值出现在1月中旬或者2月初;这与本研究所得结论是一致的。
植物群落不同,所引起的温湿度、有机质含量和pH值等生态环境因子不尽相同,因而土壤呼吸有所差异[20]。8 m行宽林间小藜—牛筋草地植株个体较小,地表植物群落稀疏,有利于空气中二氧化碳扩散流通,且土壤内根系密集,这些均导致其土壤呼吸通量较大;2 m行宽林间苍耳地与8 m行宽林间苍耳地相比,由于2 m行宽林间苍耳地距离杨树林较近,受杨树林根系呼吸的影响更大,导致其土壤呼吸值大于8 m行宽林间苍耳地,说明杨树根系对土壤呼吸有增强作用;2 m行宽林间裸地由于地表无植物群落,土壤中没有活的植物根系,其土壤呼吸主要由土壤中微生物活性等决定,而土壤微生物活性主要与土壤性质、土壤温度、土壤含水量等相关,其变化趋势与土壤温度变化一致,所以土壤温度是决定土壤呼吸的主要因素之一。2 m行宽林间裸地由于靠近杨树人工林,地表枯落物明显多于远离杨树人工林的8 m行宽林间水蓼—灰灰菜地与8 m行宽林间苍耳地,且其土壤呼吸受杨树林根系影响,所以其土壤呼吸与8 m行宽林间水蓼—灰灰菜地和8 m行宽林间苍耳地相差不大,可见枯落物对土壤呼吸也具有加强作用。
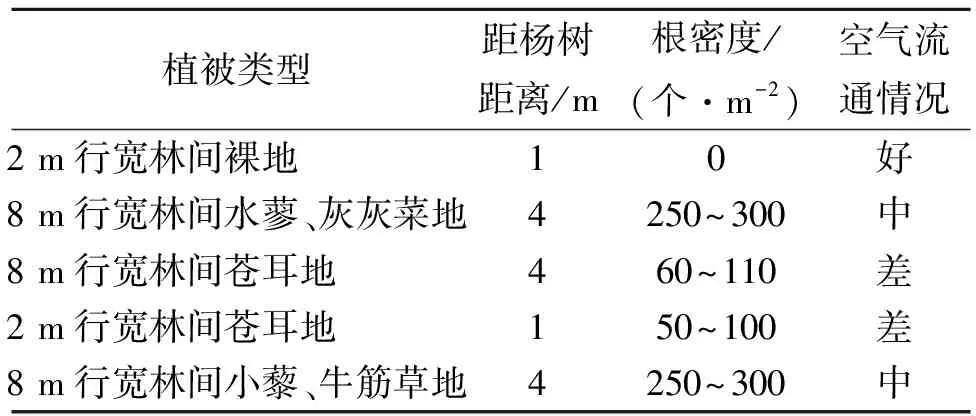
表3 不同植物群落土地概况
4.2不同植物群落下土壤呼吸对土壤温湿度的响应
土壤温度和湿度是影响土壤呼吸的主要因素,由于土壤温度和湿度的不同配置引起土壤呼吸的季节波动[21]。目前关于土壤呼吸与土壤温度或土壤含水量之间单因素关系模型的研究较多,而揭示土壤呼吸与土壤温湿度二者之间关系的双变量模型较少。本研究采用双因素关系模型Rs=aebTsWc对土壤呼吸与土壤温度和土壤含水量之间的关系进行拟合,结果显示土壤温度和土壤含水量共同解释了5种植物群落土壤呼吸的49.1%~86.7%,这与杨玉盛等[16]、王小国等[14]的研究结果相似。采用双因素模型对土壤呼吸与土壤温度和土壤含水量进行拟合,其结果明显优于土壤呼吸与土壤温度或土壤含水量单因素模型。
温度是土壤呼吸的关键影响因素,由于其具有强烈的时空变异性特点,因此它表现为明显的季节变化与日变化幅度大的特征[22]。5种不同植物群落土壤呼吸季节变化特征明显,随季节变化的趋势基本一致,土壤呼吸全年总体变化均呈现先增大而后减小的趋势,与土壤温度的变化趋势完全一致;土壤呼吸最大值均出现在夏季(7月份左右),其最小值一般出现在温度较低的冬季(12月份或1月份)。这是因为土壤温度是土壤呼吸的主要影响因素,在夏季空气温度偏高,从而导致土壤温度相对较高,土壤呼吸值也处于全年最高水平;而在温度较低的冬季,土壤呼吸值均较小,甚至接近于0。这是因为,土壤呼吸不仅与温湿度等环境因子相关,同时还受土壤酶、土壤微生物活动等的影响,而土壤温度对土壤酶的活性及土壤生物的活动有至关重要的影响;在低温条件下,土壤酶活性较低,土壤生物的活动也受到抑制,因此土壤呼吸较低,随着温度的升高,二者活性逐渐升高,土壤呼吸随之增加。陈宝玉等[23]通过对川西亚地区针叶林土壤呼吸的研究发现,针叶林不同土层的土壤呼吸均为春秋低、夏季高,从7月份到11月份呈下降趋势;周海霞等[24]针对中国东北地区次生林与落叶松林的土壤呼吸的研究结果表明,天然次生林和落叶松人工林土壤呼吸速率的季节变化呈单峰曲线,从5月份开始,其土壤呼吸速率逐渐增加,7—8月份达到最大值,然后逐渐下降;这些研究所得出的结果与本研究的结论是一致的。
5种不同植物群落下,土壤呼吸对土壤温度的敏感程度均高于对土壤含水量的。其中,8 m行宽林间小藜—牛筋草地与2 m行宽林间苍耳地土壤呼吸与土壤温度的拟合程度明显高于其余三者,且二者土壤呼吸值最大;二者的土壤呼吸年变化与土壤温度年变化基本保持一致,土壤呼吸与土壤温度最高值均出现在7月份左右。此时,土壤含水量相对较低,可见,土壤呼吸随土壤温度的升高呈现出递增趋势,而随着土壤含水量的增加土壤呼吸会受到一定限制。8 m行宽林间小藜—牛筋草地由于植株体积较小,地表植物群落较为稀疏,受太阳辐射大,土壤温度受影响较大,导致其土壤呼吸比其他四种植物群落相对较高;此外,8 m行宽林间小藜—牛筋草地植被根系密度较大,对土壤呼吸也具有一定的加强作用。2 m行宽林间苍耳地虽然根系密度较小,空气流通较差,但其距离杨树人工林距离较小,杨树林的根系对土壤呼吸有加强的作用,所以其土壤呼吸比8 m行宽林间苍耳地要大。2 m行宽林间裸地地表无植物群落,其土壤呼吸本应小于其余四种植物群落下的土壤呼吸,但由于其靠近杨树人工林,在杨树根系呼吸作用及枯落物的影响下,其土壤呼吸值比8 m行宽林间苍耳地略大,可见杨树人工林对土壤呼吸的贡献值要大于地表植被苍耳根系的呼吸作用。
4.3土壤呼吸对温度的敏感性
Q10是反映土壤呼吸对温度敏感性的重要指标,其意义为土壤温度每升高10℃时土壤呼吸增加的倍数[24]。本研究中得到的土壤呼吸Q10值大小顺序为:8 m行宽林间小藜—牛筋草地(1.46~4.62)>2 m行宽林间苍耳地(1.28~3.67)>2 m行宽林间裸地(1.4~3.39)>8 m行宽林间水蓼—灰灰菜地(1.31~3.56)>8 m行宽林间苍耳地(1.32~3.49),说明8 m行宽林间小藜—牛筋草地土壤呼吸对温度最为敏感。
5种不同植物群落土壤呼吸Q10值均表现为春季最大,冬季次之,夏季最小;与土壤温度呈显著负相关,即随着土壤温度的升高土壤呼吸Q10值逐渐降低,土壤呼吸对土壤温度的敏感性逐渐下降。土壤呼吸Q10值最大值出现在春季主要是由于春季为生长季的初期,土壤微生物活动剧烈,表层植被根系呼吸作用明显增强,造成春季土壤呼吸Q10值最大;而在温度较高的夏季,可能由于空气温度较高,导致植物气孔关闭,从而降低植被呼吸作用对土壤呼吸的影响,且较高温度条件下各种生物活性均较高,对土壤呼吸产生的影响较为复杂,可能导致土壤温度对土壤呼吸的影响作用降低。因此,在温度较高的夏季土壤呼吸Q10值反而变小,温度敏感性降低。许多研究表明,Q10值存在一定的时间和空间变异,它随着季节变化而变化[14,25]。Kutsch和Kappen对德国北部的一个玉米单作地和一个轮作地的土壤呼吸的研究结果表明,在温度较低的冬季Q10值明显高于较高温度夏季的[26];张鸽香等[27]对南京市公园绿地不同植被类型植被的土壤呼吸研究发现,温度较低时土壤呼吸对温度更为敏感,而在温度升高时土壤呼吸对温度的敏感性反而下降;为消除其他因子如土壤含水量和植被枯落物质量对土壤呼吸的影响,Kirschbaum综合了许多室内控制培养试验后发现Q10值在低温下较高,而在高温下较低[28];这些研究结果与本研究得到的结论完全一致。
此外,还有研究发现,除了温度影响土壤呼吸Q10值外,土壤含水量对其也有一定影响。Davidson等对北美硬木林的研究结果表明,有良好排水条件的试验点Q10值较低,而在土壤含水量经常较高的试验点Q10值较高[12],这与本研究的结果略有不同;本研究中土壤含水量在夏季较大,而Q10值在此时确为一年中的最小值,Q10最大值反而出现在土壤含水量相对较小的春季,这可能是因为在本研究中土壤含水量不是对Q10值的主要因素。土壤含水量的改变势必会影响植物根系分布、根系呼吸及土壤微生物活动的改变,但本研究中土壤含水量在一年中变化范围不大,其对土壤呼吸Q10值的影响小于土壤温度的影响,可能导致其影响作用被其他影响因素所覆盖,从而导致以上结果的出现。5种不同植物群落所得土壤呼吸平均Q10值分别为2.47,2.3,2.7,2.59,2.22(表2),而房秋兰等[29]认为全球土壤呼吸的Q10值一般在2.0~2.4,平均值为2.0;牟守国[30]对温带针阔叶林及针阔混交林的研究得到阔叶林的土壤呼吸Q10值为3.6,针叶林为3.8,针阔混交林为3.2;张鸽香[27]等对南京市公园绿地不同植被类型植被的土壤呼吸研究发现,近自然林、疏林、草坪土壤呼吸Q10值分别为2.24,1.78,1.76;本研究中得到的Q10值与以往的研究结果基本一致。目前,Q10模型被广泛应于不同尺度土壤呼吸对温度变化的反映上,且大尺度的对碳循环进行估测主要通过模型推算,因此,Q10模型参数值的精确量化对生态系统碳循环研究具有重要的现实意义。
5 结 论
5种不同植物群落土壤呼吸速率最大的为8 m行宽林间小藜—牛筋草地,最小的为2 m行宽林间裸地。土壤呼吸日变化最大值均出现在11:00—14:00,最小值出现在早晨5:00—8:00。土壤呼吸在全年尺度上的变化与土壤温度的变化是一致的,最大值出现在7月份左右,最小值出现在冬季的12月份或1月份。
土壤温湿度共同解释了土壤呼吸速率季节变化的49.1%~86.7%,表明不同植物群落的土壤呼吸受土壤温度和土壤含水量的共同影响。5种不同植物群落Q10值:8 m行宽林间小藜—牛筋草地(1.46~4.62)>2 m行宽林间苍耳地(1.28~3.67)>2 m行宽林间裸地(1.4~3.39)>8 m行宽林间水蓼—灰灰菜地(1.31~3.56)>8 m行宽林间苍耳地(1.32~3.49),季节变化均表现为春季>冬季>秋季>夏季。
土壤呼吸时一个复杂的生物化学过程,土壤呼吸具有很大时空差异性,本研究只是针对不同植物群落对土壤呼吸的现有影响进行了研究,尚且无法揭示土地利用变化对土壤呼吸的影响机理及预测未来变化趋势。因此,探讨土地利用变化对土壤呼吸的影响将是未来碳循环研究的一个重要方向。
[1]Bahn M, Janssens I A, Reichstein M, et al. Soil respiration across scales:towards an integration of patterns and processes[J]. New Phytologist, 2010,186(2):292-296.
[2]Bond-Lamberty B, Thomson A. A global database of soil respiration data[J]. Biogeosciences, 2010,7(6):1915-1926.
[3]周广胜.全球碳循环[M].北京:气象出版社,2003.
[4]Singh J S, Gupta S R. Plant decomposition and soil respiration in terrestrial ecosystems[J]. The Botanical Review, 1977,43(4):449-528.
[5]Schlesinger W H, Andrews J A. Soil respiration and the global carbon cycle[J]. Biogeochemistry, 2000,48(1):7-20.
[6]Lloyd J, Taylor J A. On the temperature dependence of soil respiration[J]. Func. Ecol., 1994,8(3):315-323.
[7]Buchmann N. Biotic and abiotic factors controlling soil respiration rates in Picea abies stands[J]. Soil Biology and Biochemistry, 2000,32(11):1625-1635.
[8]Dilustro J J, Collins B, Duncan L, et al. Moisture and soil texture effects on soil CO2efflux components in southeastern mixed pine forests[J]. Forest Ecology and Management, 2005,204(1):87-97.
[9]Li H, Yan J, Yue X, et al. Significance of soil temperature and moisture for soil respiration in a Chinese mountain area[J]. Agricultural and Forest Meteorology, 2008,148(3):490-503.
[10]Betts R A. Offset of the potential carbon sink from boreal forestation by decreases in surface albedo[J]. Nature, 2000,408(6809):187-190.
[11]Cox P M, Betts R A, Jones C D, et al. Acceleration of global warming due to carbon-cycle feedbacks in a coupled climate model[J]. Nature, 2000,408(6809):184-187.
[12]Davidson E C, Belk E, Boone R D. Soil water content and temperature as independent or confounded factors controlling soil respiration in a temperate mixed hardwood forest[J]. Global Change Biology,1998,4(2):217-227.
[13]Kao W Y, Chang K W. Soil CO2 efflux from a mountainous forest-grassland ecosystem in central Taiwan[J]. Botanical Studies, 2009,50(3):337-342.
[14]王小国,朱波,王艳强,等.不同土地利用方式下土壤呼吸及其温度敏感性[J].生态学报,2007,27(5):1960-1967.
[15]乌俊杰,许建初,李云驹.西双版纳三种植物群落的旱季土壤呼吸[J].植被分类与资源学报,2012,34(4):357-365.
[16]杨玉盛,陈光水,王小国,等.中国亚热带森林转换对土壤呼吸动态及通量的影响[J].生态学报,2005,25(7):1684-1690.
[17]黄承才,葛滢,常杰,等.中亚热带东部三种主要木本群落土壤呼吸的研究[J].生态学报,1999,19(3):324-328.
[18]董彬.土壤温度和湿度对鲁东南杨树人工林土壤呼吸的协同影响[J].生态环境学报,2012,21(5):864-869.
[19]You W, Wei W, Zhang H, et al. Temporal patterns of soil CO2efflux in a temperate Korean Larch (LarixolgensisHerry.) plantation, Northeast China[J]. Trees, 2013,27(5):1417-1428.[20]马涛,周金星.滩地人工杨树林土壤呼吸变化规律与环境因子的关系研究[J].水土保持研究,2011,18(6):31-41.
[21]杨玉盛,董彬,谢锦升,等.森林土壤呼吸及其对全球变化的响应[J].生态学报,2004,24(3):583-591.
[22]李元,时伟宇,闫美杰,等.土壤呼吸影响因素概述与展望[J].水土保持研究,2013,20(5):311-315.
[23]陈宝玉,刘世荣,葛剑平,等.川西亚高山针叶林土壤呼吸速率与不同土层温度的关系[J].应用生态学报,2007,18(6):1219-1224.
[24]周海霞,张彦东,孙海龙,等.东北温带次生林与落叶松人工林的土壤呼吸[J].应用生态学报,2007,18(12):2668-2674.
[25]Janssens I A, Carrara A, Ceulemans R. AnnualQ10of soil respiration reflects plant phenological patterns as well as temperature sensitivity[J]. Global Change Biology, 2004,10(2):161-169.
[26]Kutesch W L, Kappen L. Aspects of carbon and nitrogen cycling in soils of Bornhoved Lake districtⅡ. Modelling the influence of temperature increase on soil respiration and organic carbon content in arable soils under different management[J]. Biogeochemistry,1997,39(2):207-224.
[27]张鸽香,徐娇,王国兵,等.南京城市公园绿地不同植被类型土壤呼吸的变化[J].生态学杂志,2010,29(2):274-280.
[28]Kirschbaum M U F. The temperature dependence of soil organic matter decomposition, and the effect of global warming on soil organic C storage[J]. Soil Biology and Biochemistry,1995,27(6):753-760.
[29]房秋兰,沙丽清.西双版纳热带季节雨林与橡胶林土壤呼吸[J].植物生态学报,2006,30(1):97-103.
[30]牟守国.温带阔叶林、针叶林和针阔混交林土壤呼吸的研究比较[J].土壤学报,2004,41(4):564-570.
Study on Soil Respiration of Five Plant Communities in North of Huai River
LIU Bin1,2, LU Shaowei2, SHI Yuan1, LI Shaoning2, CHEN Bo2
(1.College of Forestry, Agricultural University of Hebei, Baoding, Hebei 071000, China;2.InstituteofForestryandPomology,BeijingAcademyofAgricultureandForestrySciences,Beijing100093,China)
We studied the soil respiration of five plant communities in poplar plantation ecosystem in Xiping of He′nan Province by an automated soil CO2efflux measurement system (LI-8100, LI-COR, Lincoln, NE, USA), and compared their differences of soil respiration and analyzed its affecting factors. The results showed that: (1) the soil respiration of five plant communities was significantly different, diurnal variation and annual variation of soil respiration had the same tendency with the unimodal curve, and soil temperature variation followed with soil respiration. The sequence of soil respiration was: 8 m wide woodland with Chenopodium serotinumEleusineindica(L). Gaertn >2 m wide woodland withXanthiumsibiricumPatrin >8 m wide woodland withPolygonumhydropiper-ChenopodiumalbumLinn >8 m wide woodland withXanthiumsibiricumPatrin >2 m wide woodland with nothing except a little litter; (2) single factor model was used to analyze the effect of soil temperature and moisture on the soil respiration (Rs=aebTsorRs=aW+b): soil temperature and moisture gave the explanation of 41%~79% and 2.3%~21% of seasonal variations of soil respiration with five plant communities, respectively; but double-factor model (Rs=aebTsWc) was used to analyze the relation, soil temperature and moisture commonly gave the explanation of 49.1%~86.7% of seasonal variations, which showed that soil respiration in five plant communities was influenced by soil temperature and moisture together; (3) with the respect to the soil respiration analyzed byQ10=e10bmodel, the sequence ofQ10values was: 8 m wide woodland with Chenopodium serotinumEleusineindica(L). Gaertn (2.47) >2 m wide woodland withXanthiumsibiricumPatrin (2.3) >2 m wide woodland with nothing except a little litter (2.7) >8 m wide woodland withPolygonumhydropiper-ChenopodiumalbumLinn (2.59) >8 m wide woodland withXanthiumsibiricumPatrin (2.22), and seasonal variation ofQ10value was: spring>winter>autumn>summer, indicating that temperature sensitivity of soil respiration would decrease with rise of soil temperature.
soil respiration; different plant communities;Q10value; soil temperature
2014-12-19
2014-12-30
林业公益性行业科研专项“森林生态服务功能分布式定位观测与模型模拟”(201204101)
刘斌(1989—),男,河北行唐县人,在读硕士,主要从事水土保持研究。E-mail:binl1989x@163.com
鲁绍伟(1969—),男,河北秦皇岛人,博士,研究员,主要从事水土保持研究。E-mail:hblsw8@163.com
S154.1; S181
A
1005-3409(2015)04-0100-07
猜你喜欢
艺术大观(2023年17期)2023-07-10 06:23:41
——林间
文化纵横(2022年3期)2022-09-07 11:43:22
哈哈画报(2022年6期)2022-07-09 09:46:08
阅读(低年级)(2021年11期)2021-03-08 10:47:52
幼儿智力世界(2020年10期)2020-10-28 02:31:47
学生天地(2019年30期)2019-08-25 08:53:26
中学时代(2018年6期)2018-07-19 09:16:18
中学语文(2018年5期)2018-03-03 02:26:55
歌剧(2017年11期)2018-01-23 03:41:09
小学生(看图说画)(2017年6期)2017-11-06 06:48:00
