
Oviposition Site Selection in the Malayan Giant Frog (Limnonectes blythii) in Singapore: Conservation Implications
2015-10-31 10:56VishnuVardhanSRIDHARandDavidBICKFORD
Asian Herpetological Research 2015年3期
Vishnu Vardhan SRIDHARand David BICKFORD
Department of Biological Sciences, National University of Singapore, 14 Science Drive 4, Singapore 117543, Republic of Singapore
Oviposition Site Selection in the Malayan Giant Frog (Limnonectes blythii) in Singapore: Conservation Implications
Vishnu Vardhan SRIDHAR*and David BICKFORD
Department of Biological Sciences, National University of Singapore, 14 Science Drive 4, Singapore 117543, Republic of Singapore
Amphibians require specific habitats for breeding and loss or degradation of such habitats can negatively affect reproductive success. Oviposition site selection within a habitat is also important as site quality is linked to larval survivorship and metamorphic success. We investigated oviposition site p
of the stream-breeding frog Limnonectes blythii in Singapore through surveys and habitat measurements of breeding and non-breeding sites (N = 30 and 32, respectively). The study species L. blythii is classified as Near Threatened (NT) in the IUCN red list and is associated with medium sized forest streams. L. blythii appeared to prefer streams with higher water pH and shallower water depths for oviposition. Our findings have implications in conservation management as it provides the baseline for habitat restoration for creation of new and for preserving existing breeding habitat of L. blythii.
amphibians, anura, oviposition, reproduction, restoration, streams
1. Introduction
Amphibians being biphasic are particularly sensitive to changes in their environment (Wells, 2007). Throughout the tropics, loss of habitat and degradation constitute the greatest threat to biodiversity conservation and in recent years has been attributed as a major cause for global amphibian crisis (Wilson, 1992; Sodhi et al., 2008). While amphibians are going extinct elsewhere, in much of South and South East Asia, new species continue to be reported (Kohler et al., 2005; Meegaskumbura et al., 2002). For effective conservation mitigation, the knowledge about the life history characteristics are needed but information is currently limited. Studies on habitat usage of species at risk would provide valuable information to ensure successful conservation efforts.
Habitat selection, defined as the process of choosing resources is especially evident in ectotherms like amphibians as they may require specific microhabitats and microclimates for normal functioning of biologicalprocesses (Johnson, 1980; Bickford, 2010). These include reproductive processes such as oviposition site selection that influence hatching success and larval performance in turn affecting the species’ requirements for survival, growth and development of juveniles (Glos et al., 2008).
To understand underlying mechanisms of habitat selection, studies focussing on habitat associations of species have advanced to quantitative studies of habitat use (Heyer et al., 1994; Beard et al., 2000). By understanding what habitats organisms use, and why they use them, we might be able to better ameliorate the effects of large scale habitat loss by protecting the most important habitats. Investigating patterns and mechanisms of habitat selection could form the foundation for conservation and management plans. On the selection of parameters for investigation, Rudolf and Rodel (2005) demonstrated the importance of including multiple factors such as water presence, sediment and water depth in analysis of oviposition site selection. Other similar habitat and breeding site characterisation studies of anurans have provided valuable information on the typical factors affecting breeding habitat characterisation among stream and river breeding anurans (Alfonso and Eterovick, 2007; Fukuyama and Kusano,1992; Gillespie et al., 2004; Kupferberg, 1996) as well as their potential applications for conservation.
This study documents the breeding habitat requirements of the stream breeding frog, L. blythii (Boulenger, 1920). Our primary objectives were to identify potential breeding sites in the national parks of Singapore and identify the environmental factors underlying suitability of habitat. This would provide baseline information for subsequent conservation interventions like habitat restoration and natural resource management in Singapore and elsewhere since breeding sites are the most important aspect of variables that explain frog abundance in Singapore’s fragmented forest system (Bickford et al., 2010).
2. Materials and Methods
In this study, we sampled six small streams in Singapore located near Bukit Timah Nature Reserve, Venus Drive and Bukit Batok Nature Park. We carried out microand macro-habitat analyses for a more comprehensive picture of possible management intervention areas that could target habitat restoration via oviposition site determination. We characterised egg clutches and microhabitat in the immediate vicinity (1 m area surrounding the eggs). We then compared these habitats with random stream habitats, where egg clutches were not observed. In addition to microhabitat comparisons, we broadly characterised streams where egg clutches were observed and compared them with streams that showed no signs of L. blythii eggs during the study period. Due to the relatively small length of the streams we were not able to conclusively determine non-breeding sites in streams that contained L. Blythii eggs. Moreover, the second analysis (stream characterisation) showed streams to be broadly similar and hence allowed breeding and non-breeding habitat comparisons across streams. There is no breeding seasonality for frogs in Singapore, and all fieldwork was done between September 2012 and March 2013 (Bickford et al., 2010). Additionally, previous studies on L. Blythii have demonstrated that reproductive activity of these species is continuous (Inger, 1969).
2.1 Study species The study species Limnonectes blythii is commonly known as the Malayan Giant Frog, and belongs to the family Dicroglossidae. It is the largest native frog in Singapore, growing up to a snout-vent length of 26 cm (Chan and Goh, 2010), and is associated with medium sized streams in forested areas. Found in Peninsular Malaysia, Singapore, Borneo, Thailand and Sumatra, L. blythii is classified as Near Threatened (NT)in the IUCN red list status (van Dijk and Iskandar, 2004). L.blythii has unusual characteristics compared to most other frogs in that males are larger than females and have disproportionately large heads (Emerson et al., 1992). Additionally, these are voiceless frogs with males lacking advertisement calls (Inger, 1969). They breed throughout the year (Inger and Bacon, 1968). Since L. blythii are large, strictly terrestrial species with a home range between 6 m and 9 m (Inger, 1969), it can be assumed that population dynamics will closely mirror the suitability of the site for oviposition.
2.2 Study site Singapore is a 710.2 km2tropical island located just south of Peninsular Malaysia. Only 5% of Singapore’s forests remain due to urbanisation (Corlett, 1997). Abundant rainfall and high humidity are typical throughout the year, although dry periods occur. Though seasonal changes are minimal, there are two monsoon periods with noticeably greater rainfall - the Northeast monsoon from mid-November to March and the Southwest monsoon from mid-June to September.
We selected six streams in Singapore as study sites for visual encounter surveys (VES; Heyer et al.,1994) of L.blythii egg clutches. We selected the streams randomly aiming to cover as many streams as possible within the given time period. Due to accessibility constraints, we were not able to cover remaining streams. Four of six streams were in Bukit Timah Nature Reserve (BTNR). These were: Jungle Falls, Soraya stream, Ases stream and Fern valley. The remaining two streams were at Venus Drive and Bukit Batok Nature Park.
2.3 Microhabitat analysis Upon locating an egg clutch during a VES, we characterised the habitat as close to the clutch as possible, preferably within the clutch area. These include the following habitat parameters: air and water temperatures, pH, canopy cover, depth of water, water flow speed, relative humidity, organic matter surrounding the clutch, water colour, water turbidity, overhanging vegetation (if any) and substrate type. In breeding sites, we measured the above parameters as close to the clutch as possible and at the center of the stream in non-breeding sites.
We measured air temperature and relative humidity using a thermohygrometer, pH and water temperature using a calibrated pH probe. We used a spherical densiometer to measure canopy cover, a ruler to measure water depth. Categories like extent of organic matter surrounding the clutch, water color and water turbidity were categorized through visual observation. We categorised organic matter surrounding the clutch into: (1) little to none; (2) surrounds the clutch; and (3) clutchenveloped with debris. We categorised water colour into: (1) colourless; (2) light brown; and (3) brown. We categorised water turbidity into: (1) clear; (2) murky; and (3) extremely turbid. Each of the above three categories were assigned numerical between 1 to 3 based on the above. We classified overhanging vegetation based on presence or absence (1 or 0) 1 m above water surface. We classified the substrate on which the clutches were present in a qualitative manner (soil, leaf, organic matter, gravel and rocks). We sampled each stream at least three times before classifying it as breeding (one where eggs were observed) or non-breeding (one where eggs were not observed). We obtained 30 breeding sites.
In order to obtain non-breeding sites, we marked streams every 5 m for 100 m (Emerson and Inger, 1992). For streams where no L. blythii clutches were observed, we randomly generated four points along the stream and characterised the habitat at these points. This was done twice in each stream and since four out of the six streams were non-breeding sites, we obtained 32 nonbreeding points. We compared these non-breeding sites with breeding sites using a linear mixed effects model in order to determine significant differences in habitat parameters. Since streams ranged from 100 m to a few kilometres in length, we used linear mixed effects models (Oberg et al., 2007) to account for correlations between multiple observations per stream and to broaden inference to the entire population rather than those samples just participating in the trial (Oberg et al., 2007). We assigned streams as random effects because this allows an important source of variation by adding an additional variation source beyond the residual error. We subjected each of the parameters mentioned earlier to the linear mixed effects model to test significance and obtain correlation values.
2.4 Stream comparison After 3 VESs per stream, we designated streams as either “breeding” or “non-breeding”streams based on presence or absence of eggs. In order to study possible macroscopic differences between streams, we broadly characterised habitat in a stream at three evenly spaced points 0 m, 50 m and 100 m along each stream and averaged parameter values to obtain a better understanding of the macrohabitat. We carried out the process twice with readings taken a month apart. For this analysis, all measurements were taken at the centre of the stream. We compared streams in which breeding was observed (Venus Drive and Bukit Batok Park) with the four streams at BTNR with no breeding and tested for any significant differences. We characterised the following parameters: air and water temperatures, pH, canopy cover, depth of water, relative humidity, extent of organic matter, water colour, water turbidity, overhanging vegetation (if any).
Since this involved comparing the habitat parameters of the six streams and searching for the most significant parameter difference, a generalised linear model (GLM) was used.
3. Results
Out of the six streams surveyed, we found 16 L. blythii egg clutches at Bukit Batok stream and 14 clutches at Venus Drive stream. We observed clutches throughout the sampling period. The clutches were in various stages of development with some being only one to two days old while others were in advanced stages of development along with newly hatched tadpoles. No clutches were found in any stream sections at Bukit Timah Nature Reserve. The number of eggs in clutches varied (ranging from 100 to as high as almost 400 eggs).
We found the eggs only in those shallow areas where water flowed very slowly or was almost stationary. Breeding sites had significantly higher pH and shallower depth of water than non-breeding micro-habitats despite the fact that at the larger scale, streams were almost identical. We found L. blythii eggs on a wide variety of substrates including sand, silt, gravel and organic matter such as leaves (dead or alive) and thin roots. It is significant to note that out of the thirty egg clutches observed, none were found on rocks since this plays a role in determining the breeding habitat preference for L. blythii.
3.1 Microhabitat analysis Most egg clutches observed during sampling were in areas adjacent to or in shallow water (< 1.8 cm deep). The mean ± SD of pH and water depth in breeding sites were 6.1 ± 0.7 (5.06–6.83) and 0.7 ± 0.5 cm (0–1.8 cm) respectively as compared to those in non breeding sites pH= 4.4 ± 0.6 (3.52–5.45) and water depth=7.2 ± 4.0 cm (1.7–18 cm). The differences between them was also found to be statistically significant for pH (P = 0.0478, N = 62) and water depth (P = 0.000847, N = 62). The P, mean ± SD, range and correlation for these parameters are provided (Table 1). The ‘correlation’values in (Table 1) are interpreted the same way as coefficients from a traditional regression.
Other factors like organic matter surrounding the clutch, colour and turbidity of water as well as overhanging vegetation were not significant factors (P > 0.05) in oviposition site selection (Table 1). The mean ± SD of organic matter surrounding the clutches variedwidely and was 1.7 ± 0.8 in breeding sites and 1.8 ± 0.8 in non-breeding sites. Some clutches were found almost completely enveloped in organic debris like leaves and roots whereas others were found on a shallow portion on the stream on sand with no surrounding organic matter. Similarly colour of water (1.3 ± 0.5 in breeding sites and 1.1 ± 0.3 in non-breeding sites) and water turbidity (1.3 ± 0.4 in breeding sites and 1.1 ± 0.3 in non-breeding sites) were almost identical between breeding and non-breeding sites and varied widely depending on the substrate, extent of organic matter and presence of vegetation around the clutch. Some clutches were found in turbid, brown waters whereas others were found in extremely clear sections of shallow water. A summary of these values can be found in Table 1.
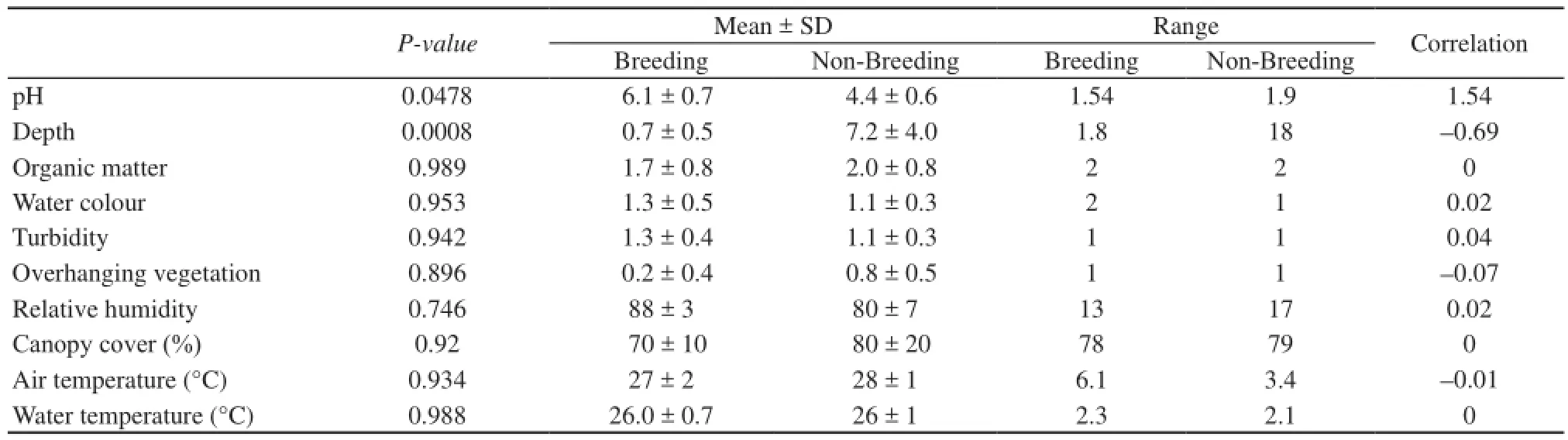
Table 1 Microhabitat analysis.
3.2 Stream comparison In order to compare breeding and non-breeding sites across streams, we analysed the habitat of a stream as a whole and compared across streams to investigate for inherent differences using a GLM. However, no significant differences were observed. Thus, parameters that we measured across streams are broadly identical without any significant differences. This suggests that though streams might be identical on a large scale, L.blythii have specific preferences on a smaller scale which affect oviposition suitability of specific sites.
4. Discussion
4.1 pH pH of breeding sites was significantly higher than pH of non-breeding sites. Minimum pH in breeding sites was 5.06. This agrees with previous findings where amphibians as a group suffer increasing mortality around pH 5 and below (Pierce, 1985; Freda and Dunson, 1986). A correlation value of 1.54 for pH indicates that an increase in pH increases chance of finding an egg clutch if all other factors were equal.
The low pH of the streams at BTNR (non-breeding sites) might be because the main body of the hill is granite (Corlett, 1988). Granite rocks tend to have a low buffering capacity against acidity (Dangles et al., 2004) explaining the low pH of these streams.
4.2 Water depth and substrate The depth of water in breeding sites was significantly lower than that in nonbreeding sites. The correlation for water depth is –0.694. This negative correlation between water depth and egg presence indicates that L. blythii prefer shallow water in streams. Previous studies have found that L. blythii deposits eggs on a wide substratum like gravel bars (Dring, 1979). Subsequently Emerson and Inger (1992) found that the egg clutches were deposited on shallow streams with gravel bed. Findings from our study partly corroborate these findings. Though clutches were found only in shallow sections of the stream, the water was not only slow fowing but we observed egg clutches on other substrates like sand, dead organic matter like leaves and roots and leaves from overhanging vegetation that were floating on the stream in addition to gravel. Since L. blythii eggs are not adhesive to the substrate, choosing shallow and choosing slow moving stream section as oviposition sites would be a strategy to ensure optimal conditions for hatchability.
This research suggests that pH and water depth are key localised factors infuencing the choice of oviposition sites by L. blythii in Singapore. This knowledge has applications in the local conservation of L. blythii by focussing on the most significant factors necessary to improve breeding success. It can also provide information to help in relocating the frogs to regions where it is endangered in order to improve its numbers in those areas.
For future studies we feel it will be extremely valuableto study the other aspects of L.blythii’s reproductive ecology such as factors affecting the success rate of emerging tadpoles using data loggers and time lapse cameras over an extended measurement period. Other oviposition site selection studies should also incorporate data on whether selection is made by the male or female. This would further aid conservation and relocation efforts of these and other similar endangered species.
Acknowledgements This study was done under National Parks Board Permit Number NP/RP12-057A and supported by National University of Singapore Undergraduate Science Research Programme funding. We would like to thank members of the Evolutionary Ecology and Conservation Lab for helping us carry out this research. We would like to thank Seshadri K.S for reviewing drafts of the manuscript and Dr. Roman Carrasco for his inputs in the statistical analyses. We would also like to thank Sheila Poo, Daniel Ng,Christopher Ziyang, Prashanth.T.R, Prasanna.V, Aswath. K.B and Vaghul Balaji for help on data analysis and field assistance.
Afonso L. G., Eterovick P. C. 2007. Spatial and temporal distribution of breeding anurans in streams in southeastern Brazil. J Nat Hist, 41: 949-963
Beard K. H., McCullough S., Eschtruth A. K. 2000. Quantitative assessment of habitat preferences for the Puerto Rican terrestrial Frog, Eleutherodactylus coqui. J Herpetol, 37(1): 10–17
Bickford D., Ng T. H., Qie L, Kudavidanage E. P., Bradshaw C. J. 2010. Forest fragment and breeding habitat characteristics explain frog diversity and abundance in Singapore. Biotropica, 42: 119–125
Boulenger G. A. 1920. A monograph of the South Asian, Papuan,Melanesian, and Australian frogs of the genus Rana. Rec Indian Mus, 20: 1–226
Chan S. H., Goh C. 2010. Frogs of Sungei Buloh wetland reserve(Amphibia: Anura). Nat Singap, 3: 103-116
Corlett R. T. 1997. The vegetation in the nature reserves of Singapore. Gard bull, 49: 147–159
Corlett R. T. 1988. Bukit Timah: The history and significance of a small rain-forest reserve. Environ Conserv, 15(1): 37–44
Dangles O.,Gessner M. O., Guerold F., Chauvet E. 2004. Impacts of stream acidification on litter breakdown: Implications for assessing ecosystem functioning. J Appl Ecol, 41: 365–378
Dring J. 1979. Amphibians and reptiles from northern Trenggaru Malaysia with descriptions of two new geckos: Cnemaspis and Cyrtodactylus. Bull Br Mus Nat Hist, 34: 181–241
Emerson S. B., Inger R. F. 1992. The comparative ecology of voiced and voiceless bornean frogs. J Herpetol, 26(4): 482–490
Freda J., Dunson W. A. 1986. Effects of low pH and other chemical variables on the local distribution of amphibians. Copeia, 1986(2): 454–466
Fukuyama K., Kusano T. 1992. Factors affecting breeding activity in a stream-breeding frog, Buregeria Buergeri. J Herpetol, 26: 88–91
Gillespie G. R., Lockie D., Scroggie M. P., Iskandar D. T. 2004. Habitat use by stream-breeding frogs in south-east Sulawesi, with some preliminary observations on community organization. J Trop Ecol, 20: 439–448
Glos J., Wegner F., Dausmann K. H., Linsenmair K. E. 2008. Oviposition-site selection in an endangered Madagascan frog: Experimental evaluation of a habitat model and its implications for conservation. Biotropica, 40: 646–652
Heyer W. R., Donnelly M. A., McDiarmid R. W., Hayek L. C., Foster M. S. 1994. Measuring and monitoring biological diversity: Standard methods for amphibians. Washington, D. C., USA: Smithsonian Institution Press
Inger R. F. 1969. Organization of communities of frogs along small rain forest streams in Sarawak. J Anim Ecol, 38(1): 123–148
Inger R. F., Bacon J. P. 1968. Annual reproduction and clutch size in rain forest frogs from Sarawak. Copeia, 1968(3): 602–605
Johnson D. H. 1980. The comparison of usage and availability measurements for evaluating resource preference. Ecology, 61: 65–71
Kohler J., Vieities D. R., Bonett R. M., Garcia F. H., Glaw F., Steinke D., Vences M. 2005. New amphibians and global conservation: A boost in species discoveries in a highly endangered vertebrate group. BioScience, 55(8): 693–696
Kupferberg S. J. 1996. Hydrologic and geomorphic factors affecting conservation of a river-breeding frog (Rana boylii). Ecol Appl, 6: 1332–1344
Meegaskumbura M., Bossuyt F., Pethiyagoda R., Manamendra-Arachchi K., Bahir M., Milinkovitch M. C., Schneider C. J. 2002. Sri Lanka: An amphibian hot spot. Science, 298(5592): 379
Oberg A.L., Mahoney D. W. 2007. Linear mixed effects model. Method Mol Biol, 404: 213–234
Pierce B. A. 1985. Acid tolerance in amphibians. BioScience, 35(4): 239–243
Rudolf V., Rodel M. 2005. Oviposition site selection in a complex and variable environment: The role of habitat quality and conspecific cues. Oecologia, 142(2): 316-325
Sodhi N. S., Bickford D., Diesmos A. C., Lee T. M., Koh L. P., Brook B. W., Sekercioglu C. H., Bradshaw C. J. A. 2008. Measuring the Meltdown: Drivers of Global Amphibian Extinction and Decline. PLoS One, 3(2): e1636
van Dijk P. P., Iskandar D. 2004. Limnonectes blythii. The IUCN Red List of Threatened Species. Version 2014.3. <www. iucnredlist.org>. Downloaded on 09 February 2015.
Wells K. D. 2007. Ecology and behaviour of amphibians. Chicago, USA: The University of Chicago Press Ltd.
Wilson E. O. 1992. The diversity of life. New York, USA: Norton and Company
Mr. Vishnu Vardhan SRIDHAR, alumnus from National University of Singapore with his research focusing on habitat ecology, conservation, man-wild conficts, and integrating science with conservation management involving local communities.
E-mail: vishna8691@gmail.com
3 October 2014 Accepted: 18 May 2015
 Asian Herpetological Research2015年3期
Asian Herpetological Research2015年3期
- Asian Herpetological Research的其它文章
- A New Species of Japalura (Squamata: Sauria: Agamidae) from Upper Lancang (Mekong) Valley of Eastern Tibet, China
- Seasonal Dynamics of Male and Female Reproductive Systems in the Siberian Salamander, Salamandrella keyserlingii (Caudata, Hynobiidae)
- Comparative Studies on Sperm Ultrastructure of Three Gecko Species, Gekko japonicus, Gekko chinensis and Hemidactylus bowrigii (Reptilia, Squamata, Gekkonidae)
- Genetic and Morphological Variations within Laudakia microlepis (Blanford, 1874) (Sauria: Agamidae) Populations in Southeastern Iran with Description of a New Subspecies
- Potential Distribution Modeling and Morphology of Pelias barani (Böhme and Joger, 1983) in Turkey
- Characterization of the Genetic Diversity of Trachemys dorbigni and Phrynops hilarii