
镉胁迫对麻疯树幼苗生理生化特性的影响
2015-10-21 19:11侯定基等
安徽农业科学 2015年21期
侯定基等


摘要
[目的] 为研究了Cd胁迫处理对麻疯树幼苗生理生化指标的影响。[方法] 通过室内沙培试验,测定丙二醛含量、根系活力、叶绿素含量、细胞膜透性、蛋白质含量、过氧化物酶活性、脯氨酸含量等的变化情况。[结果] 少量低浓度的镉(5 mg/L)对植物的生长无明显毒害作用。但,随着镉浓度的增加(10~200 mg/L),显示出一定的负效应,表现为丙二醛和蛋白质含量的增加、电导率的增大、根系活力的下降、叶绿素和过氧化物酶活性的下降。[结论] 一定浓度的镉会影响或改变麻风树部分生理生化特征,从而对麻疯树的生长造成一定的危害。
关键词 镉胁迫;麻疯树;生理生化指标
中图分类号S158.4文献标识码A文章编号0517-6611(2015)21-018-03
随着社会的发展与进步,重金属污染已越来越普遍。金属冶炼和化工企业生产、灌水(污灌)、固体废弃物、农药和化学残留等都是重金属的主要来源。镉污染是重金属污染中较常见且污染严重的一种。当前我国受镉污染的耕地面积达1.33万hm2,涉及11個省区的25个市县。在国内许多地方,在粮食、蔬菜中已检测出含量超标及严重超标的砷、铅、镉、铜等[1-3],镉大米事件也常见报道。
过量的Cd 在土壤中极易被植物吸收和积累,对植物的生长发育造成较大影响,最终造成农作物产量和品质的下降[4]。镉对植物毒害机理十分复杂。它不但影响植物细胞生长和分裂[4-5],而且对叶绿素的合成和光合作用均有抑制作用[6-7]。研究表明,镉毒害使小麦叶片的光系统I 受损严重;也有研究表明,Cd对光系统Ⅰ和光系统Ⅱ均有影响,但对后者的影响更加显著。其机理主要是镉促使叶绿体中光系统Ⅱ捕光叶绿素蛋白质复合物(LHCHⅡ)的部分寡聚体解聚成单体,且总量减少[8-9]。另外,镉对光合作用色素、光合组织等也有破坏作用[10-12],从而影响植物的光合效率[13-14]。
麻疯树(Jatropha curcus)在分类学上属大戟科(Euphorbiaceae)麻疯树属,用途十分广泛,生长速度很快,可作为饲料、农药、医药、能源等的原料。尤为重要的是,麻疯树具有耐旱瘠的特性,可以作为干热河谷地区的生态林或水源林树种[15],是我国各地均推广的一个树种[16-17]。
近年来,人们越来越重视对重金属污染环境的植物修复方法的研究[18]。笔者采用盆栽试验,研究土壤中不同浓度的镉胁迫污染对麻疯树幼苗生理生化特性的影响,为利用麻疯树修复重金属污染的环境的可行性提供参考。
1 材料与方法
1.1试验材料
供试麻疯树种子从云南壮大科技有限公司购买。
1.2试验方法
1.2.1
种子催芽。先用清水浸泡12 h,再用1∶5 000的KMnO4溶液消毒,时间为10 min。培养皿中垫1层湿润的棉花和2层滤纸,种子平铺在滤纸上,置入培养箱进行恒温培养,培养温度为25 ℃,定时用蒸馏水喷洒,以保持种子水分。
1.2.2
盆栽沙培处理。
1.2.2.1
播种育苗。准备直径8 cm塑料杯作为栽培容器, 装入珍珠岩作为栽培基质。在种子开始露白后,在培养基质上播种。每天以1/2霍格兰培养液浇淋,以保持基质湿润。
1.2.2.2
胁迫处理。设6个不同的镉浓度:0、5、10、50、100、200 mg/L。每个处理10株麻疯树幼苗,要求幼苗长势均匀、大小一致。每个处理用含不同浓度的1/2体积的培养液分别浇淋栽培基质。胁迫7 d后,测定各种生理生化指标。
1.3 测定项目与方法
试验项目参照李玲等方法[19-20]。采用无水丙酮提取、可见光分光光度法测量的方法,测定叶绿素的含量;采用硫代巴比妥酸加热显色法,测定丙二醛(MDA)含量;采用考马斯亮蓝法,测定可溶性蛋白的含量;采用愈创木酚法,测定过氧化物酶(POD)的活性;采用氯化三苯四氮唑(TTC)法,测定根系的活力;通过测定外渗液电导率的变化,测定检测植物细胞质膜的透性;采用酸性茚三酮法,测定脯氨酸含量。
1.4 数据处理
数据的整理、图表绘制都采用 Excel2003软件。
2 结果与分析
2.1镉胁迫对麻疯树幼苗丙二醛含量的影响
丙二醛是膜脂过氧化物的重要产物。它可以和氨基酸、核酸、蛋白质等活性物质交联,形成脂褐素(不溶化合物)沉积,干扰细胞的生命活动。其含量直接反映膜脂过氧化程度[12]。从图1可以看出,在镉胁迫下,随着镉浓度的增加,丙二醛含量增加,而当镉浓度达100 mg/L时丙二醛含量开始下降。这说明低浓度重金属处理能够维持细胞膜的稳定,促进麻风树幼苗的生长,而高浓度则使其受害程度加剧。
2.2镉胁迫对麻疯树幼苗根系活力的影响
根系是植物的重要器官,供应植物生长所必需的矿物质和水分,与植物生长密切相关。根系活力一般指根系的吸收、合成、氧化和还原能力等。根系活力的大小与根系代谢强度有关,根系活力越高,则根系的代谢就越旺盛。这有利于整株植株的生长。从图2可以看出,在镉处理时,麻风幼苗根系活力先降后升,当镉浓度为100 mg/L时根系TTC还原强度为最高。
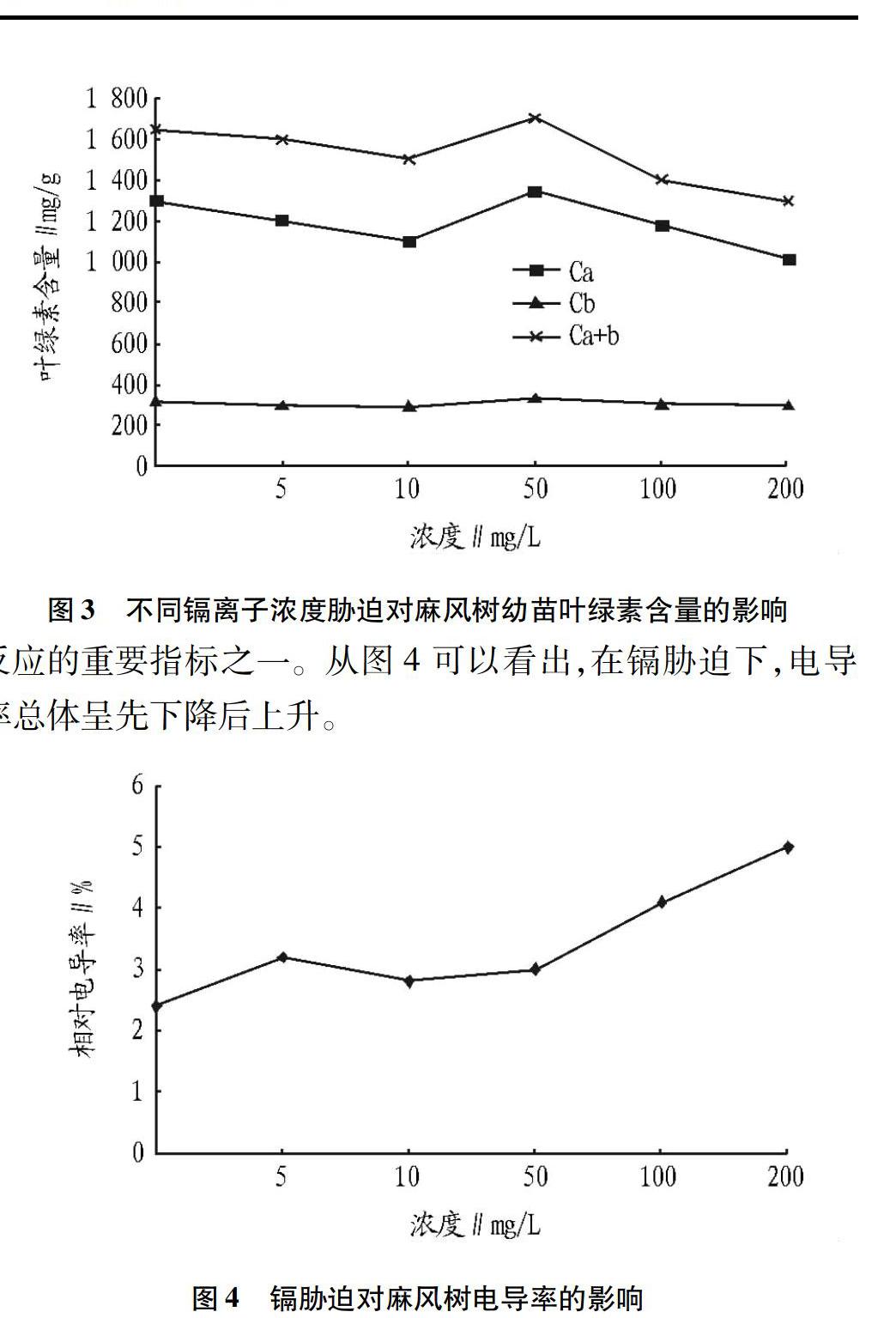
2.3镉胁迫对麻疯树叶绿素含量的影响
叶绿素的作用是进行光合作用,其含量与光合作用水平有正相关。光合作用弱,则植物的湿重会降低[9-10]。从图3可以看出,叶绿素含量随着镉浓度的增加而降低,但当镉浓度为5 mg/L时,叶绿素含量出现上升趋势;当镉浓度超过50 mg/L时,叶绿素又开始呈下降趋势,说明镉破坏了叶绿素结构和功能活性,抑制光合作用,即叶绿素生长。
2.4镉胁迫对麻疯树幼苗电导率的影响
植物细胞中的原生质膜在维持细胞内外离子的浓度差、物质的主动运输中起着非常重要的作用。细胞内外物质的交换和运输均由细胞膜调节和控制。细胞膜的选择透过性是评价植物对污染物反应的重要指标之一。从图4可以看出,在镉胁迫下,电导率总体呈先下降后上升。
2.5镉胁迫对麻疯树幼苗可溶性蛋白质含量的影响
植物体内的可溶性蛋白质大部分是参与各种代谢的酶类。可溶性蛋白质含量是一个重要的生理生化指标。人们可以通过其含量来了解植物体的总代谢水平。可溶蛋白质含量的升高会使细胞渗透浓度升高,也会使功能蛋白数量增加,有利于细胞正常代谢的维持。
从图5可以看出,不同浓度Cd2+胁迫对麻风树幼苗叶片中的可溶性蛋白含量变化存在一定的影响。随着处理浓度的增加,可溶性蛋白的含量也增加,当Cd胁迫浓度达200 mg/L时,麻风树幼苗叶片中可溶性蛋白达最大值。
2.6镉胁迫对麻疯树幼苗过氧化物酶活性的影响
过氧化物酶是一种含铁的酶,能够促进H2O2氧化酶类反应,使得植物免受伤害。其作用类似于氢的受体,在植物体的呼吸代谢中起着重要的作用。
从图6可以看出,在镉胁迫下,POD活性都随浓度升高先缓慢上升后缓慢下降。当镉浓度为10 mg/L时,POD活性达到最高值;当镉浓度为200 mg/L时,POD活性达到最小值。这表明当镉浓度较低时,它能诱导过氧化物酶消除胁迫下产生的活性氧。但是,它维持系统稳定的能力并不是无限的。当胁迫达到一定强度时,这种能力会渐渐丧失。
2.7镉胁迫对麻疯树幼苗脯氨酸含量的影响
脯氨酸的积累与逆境胁迫程度、植物的抗逆性有关。在逆境条件下,植物体内常有游离脯氨酸的积累,参与渗透调节过程,从而对细胞膜和生理代谢过程起到保护的作用。
从图7可以看出,在5.0 mg/L 镉胁迫下,脯氨酸含量增大;当镉浓度为10 mg/L时,脯氨酸含量变小,为对照组的68.32%,说明10 mg/L 镉阻碍了麻风树体内脯氨酸的合成;当镉浓度达50、100、200 mg/L,脯氨酸含量呈上升趋势,分别为对照组的114.68%、165.66%、208.92%。
3讨论
镉对植物毒害机理十分复杂,不但影响植物细胞生长、分裂,而且对植物光合作用产生影响且较复杂。镉对叶绿素的合成和光合作用均有抑制作用。活性氧代谢平衡对植物的生长发育起着至关重要的作用,而镉在植物体内的积累可以破坏活性氧代谢平衡,使得活性氧浓度超过一定阈值,产
生连锁反应,结果使得像脂肪酸之类的大分子物质发生氧化
作用,并且产生重要的脂质过氧化物丙二醛,丙二醛的积累又会引起蛋白质变性失活,破坏核酸等生物大分子, 最终使细胞膜渗透性增加,一些小分子物质及电解质外渗,导致细胞间物质交换失去平衡[17]。
镉使麻疯树幼苗MDA含量增加;抑制根系的发育,导致根系活力降低,从而抑制营养物质的吸收;影响叶绿体的合成,降低叶绿素含量,从而抑制光合作用;破坏叶片膜结构和功能,增大细胞膜透性,提高可溶性蛋白的含量;降低过氧化物酶活性。由于碳源矿物质的积累速度降低,苗期生长速度降低,脯氨酸含量增加,导致膜结构遭到破坏。
综上所述,低浓度镉对作物生长有促进作用,但是随着幼苗的生长,镉在植物体内积累,逐渐表现出抑制作用,使得麻疯树幼苗受到毒害。
安徽农业科学2015年
参考文献
[1]
杨苏才,南忠仁,曾静静.土壤重金属污染现状与治理途径研究进展[J].安徽农业科学,2006,34(3):549-552.
[2] 徐应明,李军幸.新型功能膜材料对污染土壤铅汞镉钝化作用研究[J].农业环境科学学报,2003,22(l):86-89.
[3] 王凯荣.我国农业重金属污染现状及其治理利用对策[J].农业环境保护,1997,16(6):174-178.
[4] 陈怀满.土壤-植物系统中的重金属污染[M].北京:科学出版社,1996:71-125.
[5] 劉海亮,崔世民,李强,等.镉对作物种子萌发、幼苗生长及氧化酶同工酶的影响[J].环境科学,1991,12(6):29-31,37.
[6] 陈桂珠.重金属对黄瓜籽苗发育影响的研究[J].植物学通报,1990,7(1):34-39.
[7] 洪仁远,蒲长辉.镉对小麦幼苗的生长和生理生化反应的影响[J].华北农学报,1991,6(3):70-75.
[8] 慈敦伟,姜东,戴廷波,等.镉毒害对小麦幼苗光合及叶绿素荧光特性的影响[J].麦类作物学报, 2005,25(5):88-91.
[9] 杨丹慧,许春辉,王可玢,等.镉离子对菠菜叶绿体色素蛋白质复合物及激发能分配的影响[J].植物学报,1990,32(3):198-204.
[10] 秦天才,阮捷,王腊娇.镉对植物光合作用的影响[J].环境科学与技术,2000(S1):33-35,44.
[11] GUPTA A,SINGHAL G S.Inhibit ion of PS Ⅱ activity by copper and its effect on spectral properties on intact cells in Anacystis nidulans[J].Environ Exper Bot,1995,35:435-439.
[12] OUZOUNIDOU G.Copperinduced changes on growth,methal content and photosynthetic function of Al yssum montanum plants[J].Environ Experi Bot,1994,34(2):165-172.
[13] 王友保,刘登义.Cu、As 及其复合污染对小麦生理生态指标的影响[J].应用生态学报,2001,12(5):773-776.
[14] 林义章,徐磊.铜污染对高等植物的生理毒害作用研究[J].中国生态农业学报,2007,15(1):201-204.
[15] CARGNELUITTI D,TABALDI L A,SPANEVELLO R M,et al.Mercury toxicity induces oxidative stress in growing cucumber seedlings[J].Chemosphere,2006,5(3): 999-1006.
[16] 何文淑,肖荣贵,杨晓琼,等.麻疯树在贫困地区农村发展和生态建设中的开发前景[J].中国中医药信息杂志,2002,7(10): 56.
[17] 魏琴,赖家业,周锦霞,等.干旱胁迫下麻疯树毒蛋白的Western杂交分析[J].北京林业大学学报,2004,3(5): 23.
[18] 母波,韩善华,张英慧,等.汞胁迫对植物细胞结构与功能的影响[J].中国微生态学杂志,2007,19(1): 112-118.
[19] 李玲.植物生理学模块实验指导[M].北京:科学出版社,2009.
[20] 张志良,瞿伟菁,李小方.植物生理学实验指导[M].北京: 高等教育出版社,2009.
猜你喜欢
阅读与作文(小学低年级版)(2018年9期)2018-10-26
阅读与作文(小学低年级版)(2018年10期)2018-10-25
贵州农业科学(2015年5期)2015-03-21
山东体育学院学报(2014年4期)2014-09-22
现代农业科技(2009年19期)2009-03-20
北方人(2009年12期)2009-02-01
