
尖孢镰刀菌致病性菌株遗传多样性的ISSR分析
2015-10-21 16:47肖荣凤陈燕萍朱育菁詹洪刘波葛慈斌
热带作物学报 2015年11期
关键词:遗传多样性
肖荣凤 陈燕萍 朱育菁 詹洪 刘波 葛慈斌
摘 要 致病性尖孢镰刀菌(Fusarium oxysporum)是一种世界性分布的作物土传病原真菌,为明确其不同寄主及专化型菌株遗传差异性和亲缘关系,利用ISSR分子标记技术对来源于不同寄主(专化型)及地理分布的43个菌株进行了遗传多样性分析。结果表明,供试的15条引物通过ISSR-PCR 扩增,共扩增出147条条带,其中多态性条带为144条,平均多态性条带达98.0%。43个菌株的聚类结果表明,各菌株间的遗传相似系数为0.67~0.97,且遗传多样性与其寄主和地理来源没有明显的相关性。进一步将供试菌株中葫芦科与茄科作物尖孢镰刀菌分别进行聚类分析,当遗传相似系数为0.84时,16个葫芦科的菌株可分为6个ISSR类群;当遗传相似系数为0.77时,16个茄科的菌株亦可分为6个ISSR类群;且均表现为相同寄主或专化型菌株因地理来源不同,被归属于不同的ISSR类群,说明寄主或专化型相同的菌株与地理来源存在一定的相关性。
关键词 尖孢镰刀菌;专化型;ISSR-PCR;遗传多样性
中图分类号 S43;Q78 文献标识码 A
Abstract Pathogenic Fusarium oxysporum is one of the most common soil-borne fungal pathogens all over the world. To specify genetic difference and phylogenetic relationship of F. oxysporum strains,the genetic diversity of 43 strains with different host and geographic origin or formae speciales strains was analyzed using the ISSR-PCR molecular marker technique. The results showed that 147 fragments were amplified from 43 strains by 15 tested primers. Among them, the polymorphic band was 144,which accounted for 98.0% in the total amplified fragments. The ISSR analysis showed that genetic similarity coefficient was 0.67-0.97, suggesting a high genetic variability within 43 strains. However, the results did not reveal a clear relationship between variability and the host/geographic origin. The isolates from cucurbitaceae and solanaceae crops were further performed the cluster analyses of genetic variability, respectively. At the level of 0.84,16 isolates from cucurbitaceae crops could be obviously divided into six ISSR groups. At the level of 0.77, 16 isolates from solanaceae crops could be also obviously divided into six ISSR groups. Moreover, these F. oxysporum isolates from the same host or formae speciales strains could be divided into different ISSR groups due to their different geographic origin. The results indicated that there was a relationship in a certain degree between variability and geographic origin within the isolates from the same hosts or the same formae speciales strains.
Key words Fusarium oxysporum;Formae speciales;ISSR-PCR;Genetic diversity
doi 10.3969/j.issn.1000-2561.2015.11.014
由尖孢镰刀菌(Fusarium oxysporum)引起的作物枯萎病是一种世界性分布的土传病害,也是造成作物连作障碍的主要原因之一。该病菌可危害各种作物,一般造成20%~30%产量损失,严重者可导致绝收[1-4]。该病原菌在遗传上十分复杂,且极易发生变异,导致菌株间的差异较大,仅靠传统的形态学分类和致病性检测进行种间及种内鉴定,难以准确揭示物种的进化关系。目前,ISSR分子遗传标记技术由于其稳定性和重复性较好、操作简单快速、所需DNA量少、不需要活体材料、也没有组织特异性等特点[5-6],在多种植物病原菌的遗传多样性研究中得到应用[7-10]。
在尖孢镰刀菌的研究方面,相关的ISSR遗传多样性研究表明其与寄主、地理来源和毒力等方面可能存在一定的相关性。段会军等[11]研究发现河北省西瓜枯萎病菌的ISSR遗传多样性与生理小种之间存在一定相关性,但与菌株的地理来源无相关性。María等[12]分析了来自不同国家的梨孢镰刀菌的ISSR遗传多样性,结果表明ISSR类群与寄主和地理来源无明显相关性。苑琳等[13]对尖孢镰刀菌胡麻专化型的分析表明,ISSR多样性与地理来源存在相关性。刘东等[14]研究发现尖孢镰刀菌黄瓜专化型菌株ISSR的遗传多样性与菌株地理来源及毒力存在相关性。王建明等[15]和李新凤等[16]的研究表明,尖孢镰刀菌同一類群中菌株之间的遗传相似性与地理来源存在一定的相关性。张述义等[17]研究表明,不同寄主的尖孢镰刀菌的ISSR 类群划分与菌株的地理来源没有明显的相关性,但分离自相同寄主的各菌株间的遗传相似性与菌株的地理来源有一定的相关性。上述相关研究表明,尖孢镰刀菌的遗传变异存在复杂性和多样性,不同寄主与地理来源的菌株在遗传变异上存在较大差异性,可能与菌株进化过程中受自然环境条件影响及与寄主发生协同进化等因素有关,而且菌株的遗传变异也可能引发寄主植物抗性的改变,从而影响该病害的防治。因此,研究不同寄主尖孢镰刀菌的遗传多样性对该病原菌的种系演化和病害防治具有重要的意义。但目前如番茄、辣椒、甜瓜和苦瓜等作物尖孢镰刀菌ISSR的遗传多样性还未见报道,本研究收集了不同寄主(专化型)和地理来源的尖孢镰刀菌43株,拟采用ISSR-PCR技术进行遗传多样性研究,旨在探讨这些菌株的遗传多样性与寄主或专化型、地理分布的关系,为尖孢镰刀菌的遗传分化、系统发育和病害防治等领域的研究提供理论依据。
1 材料与方法
1.1 材料
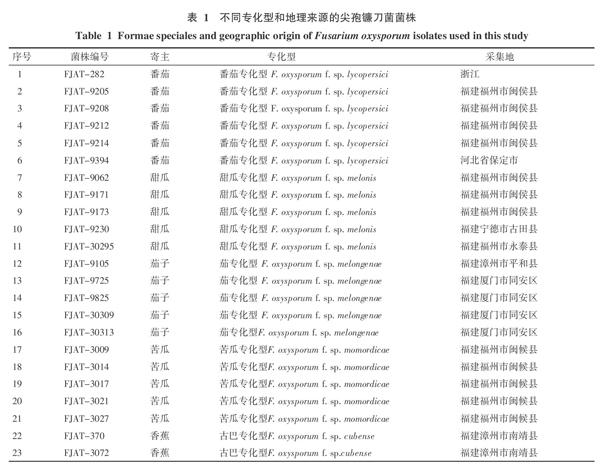
供试菌株:尖孢镰刀菌不同专化型菌株共43株(表1)。除菌株FJAT-282、FJAT-9394和FJAT-3322分别为浙江大学、河北省农林科学院和中国热带农业科学院赠送外,余者均由本研究室从福建省内不同地区采集与分离保存。
培养基:PDA、PDB。
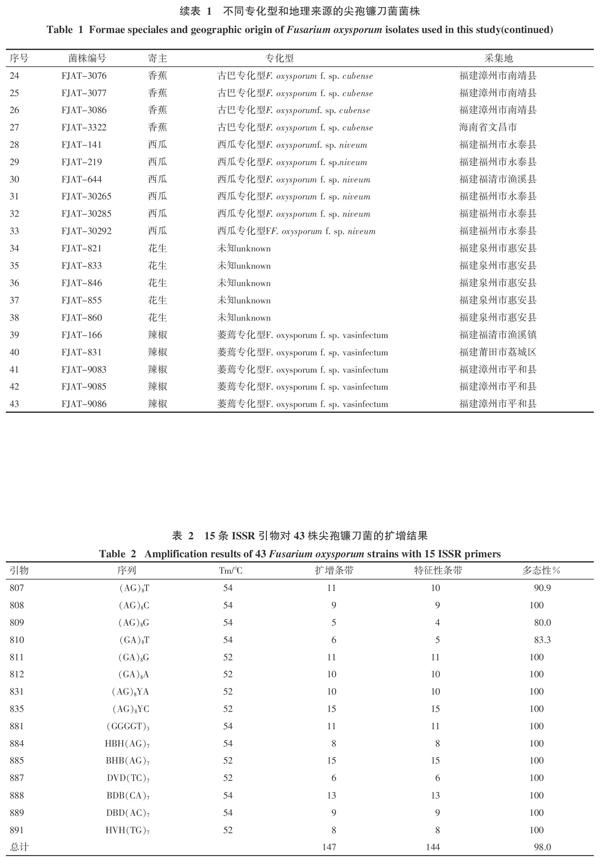
试剂:10 mmol/μL dNTPs、10×Buffer(无Mg2+)、25 mmol/μL MgCl2和5 U/μL Taq酶购自上海博尚生物技术服务有限公司;用于ISSR-PCR 反应的15条ISSR引物参考王建明等文献[15],来自于英国哥伦比亚大学(University of British Columbia)公布的序列(表2),引物均由上海博尚生物技术服务有限公司合成。
仪器:Gel Doc-ItTM UVP凝胶成像系统;Biometra PCR仪;BIO-RAD PowerPacTM Basic电泳仪;Thermo Scientific NanoDrop 2000C分光光度计。
1.2 方法
1.2.1 尖孢镰刀菌菌丝培养及DNA提取 将供试的43株尖孢镰刀菌菌株经PDA平板活化后,分别接种到PDB培养液中,25 ℃恒温振荡培养7 d,过滤获得菌丝,烘干备用。菌株DNA提取方法参考苏明星等[18]方法。制备好的DNA采用Thermo Scientific NanoDrop 2000C 分光光度计测定DNA的浓度,用灭菌的去离子水将DNA浓度统一调节成20 ng/μL,备用。
1.2.2 尖孢镰刀菌的ISSR引物筛选与扩增 ISSR 扩增反应体系参考文献[15,19]。20 μL的PCR反应体积:1.0 U Taq DNA聚合酶,1.2 μL MgCl2(25 mmol/μL),dNTPs 0.3 μL(10 mmol/μL),0.8 μL引物(10 μmol/μL),1 μL模板DNA(20 ng/μL)。PCR循环参数:94 ℃ 5 min;94 ℃ 45 s,52 ℃/54 ℃ 45 s,72 ℃ 2 min,共35个循环;72 ℃ 7 min。
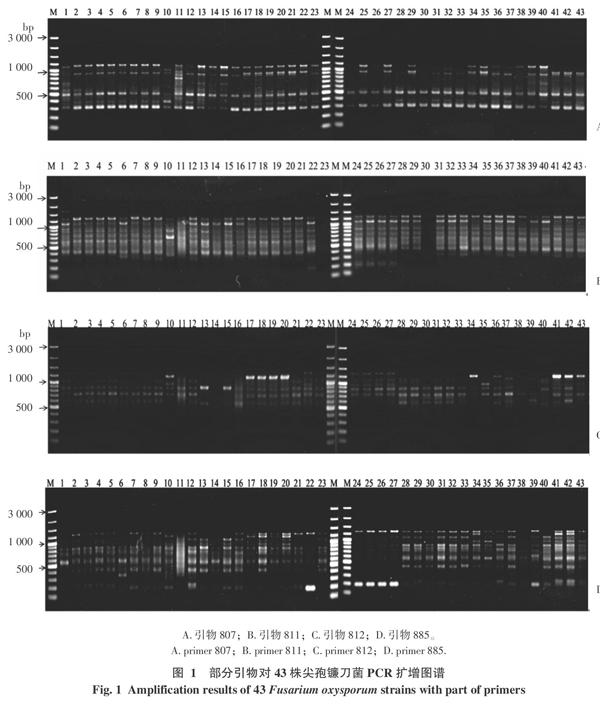
随机选取3个来自不同寄主的菌株基因组DNA为模板,利用合成的15条ISSR引物对其进行扩增,然后比较扩增结果,从中选择多态性和重复性好的引物对43 株供试菌株的基因组进行PCR 扩增。扩增反应结束后,在2%琼脂糖凝胶中分离扩增产物并拍照。
1.2.3 尖孢镰刀菌的ISSR-PCR多态性分析 ISSR引物的PCR扩增产物经电泳分离后,记录每条引物对每个菌株的DNA的扩增结果。在电泳图谱中每条扩增条带都代表引物与模板DNA互补的一对结合位点,可记为1个分子标记。采用Gel-Pro Analyzer软件对ISSR电泳图谱中的条带进行自动识别和记录,并进行人工校对。将PCR扩增的DNA条带转换成0/1矩阵,无带的记为0,有带的记为1,得到ISSR分子标记的原始0/1二元数据矩阵。利用NTSYSpc(Version 2.10e)软件进行数据处理,应用SIMQUAL(Similarity for Qualitative Date)法进行菌株间的遗传相似系数计算,以非加权类平均法(UPGMA)进行聚类分析并构建系统聚类图。
2 结果与分析
2.1 尖孢镰刀菌的ISSR引物的筛选与扩增
15条ISSR引物对43株尖孢镰刀菌的扩增结果表明(表2),所有引物均具背景清晰、信号强、多态性明显的特点,部分扩增结果图片见图1。共扩增出147条条带,多分布在500~2 000 bp之间,其中特征性条带(多态性位点)为144条,多态性平均比例为98%。不同引物扩增条带数不等,条带数在5~15条之间,多数为9~11条之间,平均每条引物产生多态性条带9.8条。因此供试的15条引物均可用于供试的不同寄主的尖孢镰刀菌遗传多态性分析。
2.2 不同寄主专化型尖孢镰刀菌的ISSR-PCR多态性分析
对43株来自8个不同寄主或专化型尖孢镰刀菌的ISSR-PCR扩增,聚类分析结果表明(图2):各菌株间的遗传相似系数为0.67~0.97,且遗传多样性与其寄主和地理来源没有明显的相关性。有些寄主或地理来源相同的菌株间的遗传距离比寄主或地理来源不同的菌株间的遗传距离大,有些则相反。如1号菌(来自浙江)与同寄主专化型的2~5号菌株(均来自福州市闽侯县)遗传距离相差较大;而34号分离自花生的菌株(来自泉州市惠安县)与17~18号分离自苦瓜专化型(福州市闽候县)遗传距离较近。但专化型(寄主)和地理来源相同的菌株却有优先聚到一起的趋势,如番茄专化型菌株2~5号(均来自福州市闽侯县)、茄专化型菌株13~16号(均来自厦门市同安区)和香蕉古巴专化型22~25号(均来自漳州市南靖县)。
为进一步分析这些尖孢镰刀菌的ISSR-PCR多态性及遗传相似性,从中将寄主为葫芦科和茄科作物的尖孢镰刀菌菌株分别进行聚类分析。
葫芦科作物尖孢鐮刀菌的聚类分析结果表明(图3),16株菌株遗传相似系数在0.62~0.97之间。当遗传相似系数为0.84时,供试的16株尖孢镰刀菌可明显地分成6个ISSR类群(ISSR Groups,IGs):甜瓜专化型菌株可分为3个IGs,IGsⅠ为7~9号菌(均来自福州市闽侯县)、IGsⅤ为10号菌(来自宁德市古田县)和IGsⅥ为11株号(来自福州市永泰县),苦瓜专化型菌株均归为IGsⅡ(17~21号菌,来自福州市闽侯县);西瓜专化型菌株分为2个IGs,IGsⅢ为28、29号菌、31~33号菌(均来自福州市永泰县)和IGsⅣ为30号菌(来自福清市渔溪镇)。上述结果表明,相同寄主专化型菌株因地理来源不同,遗传相似距离较大,亲缘关系较远;相同寄主专化型且地理来源相同的菌株,遗传相似距离较小,亲缘关系较近。
茄科作物尖孢镰刀菌的聚类分析结果表明(图4),16株菌株遗传相似系数在0.62~0.94之间。当遗传相似系数为0.77時,供试的16株尖孢镰刀菌可明显地分成6个ISSR类群:番茄专化型菌株可分为2个IGs,IGsⅠ为1号菌(来自浙江),IGsⅡ为2~6号菌(分别来自福州市闽侯县和河北保定);寄主为辣椒的萎蔫专化型可分为3个IGs,IGsⅢ为40号菌株(来自莆田市荔城区),IGsⅣ为41~43号菌(来自漳州市平和县),IGsⅥ为39号菌(来自福清市渔溪镇);茄专化型13~16号菌株归为IGsⅤ(均来自厦门市同安区),但12号茄专化型菌株(来自漳州平和县)例外,与番茄专化型菌株归为IGsⅡ。上述结果同样表明,相同寄主专化型菌株因地理来源不同,遗传相似距离较大,亲缘关系较远;相同寄主专化型且地理来源相同的菌株,遗传相似距离较小,亲缘关系较近;但个别菌株出现例外。
3 讨论与结论
ISSR分子标记通常为显性标记,具有很好的稳定性和多态性[20]。本研究对43个尖孢镰刀菌菌株进行ISSR-PCR扩增,供试的15条引物均具有良好的多态性,且平均多态性条带达98.0%,说明供试菌株的DNA 基因组在SSR区域的遗传差异明显、多态性高,所筛选的引物可用于ISSR-PCR扩增进行尖孢镰刀菌种内遗传差异分析,该结果与段会军等[11]和王建明等[15]的结果一致。
不同寄主与地理来源尖孢镰刀菌的遗传变异存在复杂性和多样性,影响着该病原菌的种系演化和病害防治,明确其遗传变异具有重要的意义。本研究对43株来自于不同寄主或专化型尖孢镰刀菌的ISSR-PCR遗传多态性分析结果表明,各专化型菌株间的遗传相似系数与其寄主和地理来源没有明显的相关性。有些寄主或地理来源相同的菌株间的遗传相似系数比寄主或地理来源不同的各菌株间的遗传相似系数大,有些则相反。说明不同专化型类群菌株中,当寄主差异较大时,利用ISSR-PCR遗传多态性分析并不能很好的反映菌株与寄主或地理来源的相关性,但寄主和地理来源相同的菌株却有优先聚到一起的趋势,该结果与张述义等[17]的研究结果一致。通过进一步将供试菌株进行细化分类重新聚类分析,发现葫芦科与茄科作物尖孢镰刀菌ISSR-PCR遗传多态性结果均表明,相同寄主专化型菌株因地理来源不同,遗传相似距离较大,亲缘关系较远;相同寄主专化型且地理来源相同的菌株,遗传相似距离较小,亲缘关系较近;但有个别菌株会出现例外。该结果可说明在同一类群中不同菌株之间的遗传相似性与菌株的地理来源有一定的相关性,与王建明等[15]的结果有相似性,而且其认为简单地提出分子类群与菌株的地理来源没有相关性是不准确的。Bentley等[21]研究表明,镰刀菌在很小的地理隔离就有多样性,甚至在间隔l m的种植行种系就出现明显的差异。本研究中如甜瓜专化型和萎蔫专化型菌株虽然均来自福建省内,因地理来源于不同市县,均分别归属于不同的ISSR类群,说明不同专化型菌株在遗传变异上有丰富的多样性,同时在相似种群内(均来自葫芦科或茄科)与地理和寄主来源存在一定相关性,这对后续田间作物品种种植的合理布局与病原菌的遗传变异监测具有重要的理论指导,通过对田间菌株的遗传多样性监测,进行作物的轮作、间作和病害防治指导,以减轻病害的发生。此外,本研究中由于供试菌株数量有限,可能导致结果存在一定的局限性。因此,下一步的研究还待收集更多来源广泛的菌株,并对特定的某种专化型菌种进行种下的遗传多态性研究。
参考文献
[1] 杨莹莹, 刘 波, 肖荣凤, 等. 番茄、茄子和辣椒枯萎病原菌分子鉴定及其致病性测定[J]. 热带作物学报, 2012, 33(5): 906-912.
[2] Xiao F, Zhu J, Li D, et al. Studies on vascular infection of Fusarium oxysporum f. sp. cubense race 4 in banana by field survey and green fluorescent protein reporter[J]. ESci J Plant Pathol, 2013, 02(01): 44-51.
[3] Dubey S C, Suresh M, Singh B. Evaluation of Trichoderma species against Fusarium oxysporum f. sp. ciceris for integrated management of chickpea wilt[J]. Biological Control, 2007: 118-127.
[4] Postma J, Luttikholt A J G. Colonization of carnation stems by a nonpathogenic isolate of Fusarium oxysporum and its effect on Fusarium oxysporum f. sp. dianthi[J]. Can, J, Bot, 1996, 74: 1 841-1 851.
[5] Leslie J F, Anderson L L, Bowden R L, et al. Inter- and intra-specific genetic variation in Fusarium[J]. International Journal of Food Microbiology, 2007, 119: 25-32.
[6] Chandra N S, Wulff E G, Udayashankar A C, et al. Prospects of molecular markers in Fusarium species diversity[J]. Applied Microbiology and Biotechnology, 2011,90:1 625-1 639.
[7] 宋培琳, 李子钦, 杨家荣. 棉花黄萎病菌遗传多样性的ISSR分析[J]. 西北农林科技大学学报(自然科学版), 2011, 39(1): 113-125.
[8] 袁洪波, 艾尼江, 赵建军, 等. 棉花黄萎病菌致病力分化与ISSR遗传变异分析[J]. 华北农学报, 2013, 28(5): 84-89.
[9] 郝丽芬, 宋培玲, 李子钦, 等. ISSR标记分析油菜黑胫病原菌遗传多样性[J]. 中国油料作物学报, 2014, 36(1): 098-0 105.
[10] 李挺丹, 彭世文, 王宗华, 等. 应用ISSR-PCR分析福建水稻纹枯菌的遗传多样性[J]. 植物病理学报, 2010(2): 76-84.
[11] 段会军, 张彩英, 李喜焕, 等. 基于RAPD、 ISSR和AFLP对西瓜枯萎病菌遗传多样性的评价[J]. 菌物学报, 2008(3), 27(2): 267-276.
[12] María I D, Eliana C, Sebastián A S. Characterization of a Fusarium poae world-wide collection by using molecular markers[J]. Eur J Plant Pathol, 2014, 140: 119-132.
[13] 苑 琳, 刘姗姗, 路福平, 等. 尖孢镰刀菌胡麻专化型(Fusarium oxysporum f. sp. lini)ISSR标记聚类分析[J]. 中国油料作物学报, 2012, 34(2): 193-200.
[14] 刘 东, 代丽婷, 蒲子婧, 等. 黄瓜枯萎病菌毒力、营养养体亲和性及ISSR分析[J]. 植物病理学报, 2012, 42(5): 456-465.
[15] 王建明, 李蕊倩, 李新凤, 等. 尖孢镰孢菌及芬芳镰孢菌遗传多样性的ISSR分析. 植物病理学报, 2011, 41(4): 337-344.
[16] 李新凤, 张光明, 王建明, 等. 21株马特组镰孢菌遗传多样性的ISSR分析[J]. 应用生态学报, 2012, 23(5): 1 339-1 344.
[17] 张述义, 李新凤, 韦晓艳,等. 33株尖孢镰刀菌遗传多样性的ISSR分析. 生态学杂志, 2013, 32(5): 1 195-1 202.
[18] 苏明星, 肖荣凤, 朱育菁, 等. 苦瓜枯萎病病原的鑒定及植株体内菌量测定[J]. 中国蔬菜, 2010, 16: 62-66.
[19] 李蕊倩, 何 瑞, 张跃兵, 等. 镰刀菌ISSR标记体系的建立及遗传多样性分析[J]. 中国农业科学, 2009, 42(9): 3 139-3 146.
[20] Tsumura Y, Ohba K, Strauss S H. Diversity and inheritance of intersimple sequence polymorphisms in Douglas-fir(Pseudotauga menziesii)and Sugi(Cryptomeria japonica)[J]. Theoretical and Applied Genetics, 1996, 92: 40-45.
[21] Bentley A R, Milgroom M G, Leslie J F, et al. spatial aggregation in Fusarum pseudograminearum populations from the Australian grain belt[J]. Plant Pathol, 2009, 58: 23-32.
