
甘蔗苗期不同叶位叶绿素荧光特性研究
2015-10-21 01:04安东升魏长斌曹娟窦美安
热带作物学报 2015年11期
安东升 魏长斌 曹娟 窦美安
摘 要 為明确不同叶位和叶片不同部位叶绿素荧光特性的分布规律及其随生长时间的变化规律,进而实现甘蔗光合潜力的快速评价,本研究通过桶栽试验对3个甘蔗主栽品种(ROC22、ROC16、桂柳05-136)不同叶位和叶片不同部位的叶绿素荧光特性进行比较分析,并研究叶绿素荧光参数对生长时间的响应。结果表明:光系统II(PSII)量子效率(ΦII)随着倒数叶位的增加而下降,在叶片不同部位之间表现为叶中部>叶尖端>叶基部,相同叶位不同品种间ΦII的差异不显著;可调节性能量耗散(ΦNPQ)则随着倒数叶位的增加而上升;非调节性能量耗散(ΦNO)在不同叶位(除倒四叶)及品种间并无显著差异。随着生长时间的推进,上部叶位具有较高的ΦII和较低的ΦNPQ;中部叶位的ΦII和光化学猝灭(qL)变化很小;下部叶位在叶片生长的前两周,能量分配在ΦII和ΦNPQ之间进行,ΦNO基本保持不变;而超过3周后,3个供试品种的ΦII、qL和最大光能利用效率(Fv/Fm)均有不同程度的下降,降幅为ΦII>qL>Fv/Fm,同时伴随着ΦNO的增加。上述结果表明光能利用效率以上部叶位最高,中部叶位稳定,而下部叶位最低且差异较为显著。而不同叶位相对电子传递速率(rETR)对生长时间的响应研究表明,在苗期上部第一片全展叶的中部叶位光能转化能力最强,可选择该叶片的中部叶位进行叶绿素荧光特性横向比较。本研究初步明确了不同叶位光能的利用效率及其随生长时间的变化趋势,为如何选择合适的叶位进行叶绿素荧光检测,进而开展甘蔗冠层光合潜力的快速评价奠定基础。
关键词 甘蔗;叶绿素荧光;叶位;生长时间
中图分类号 S314 文献标识码 A
Abstract To investigate the light excitation energy allocation characters in photosystem II (PSII) of different leaf position and growth time for three tested sugarcane cultivars(ROC22, ROC16 and GUI LIU 05136),several chlorophyll fluorescence parameters were measured and analyzed by using pot experiments. Results showed that, the quantum efficiency of PSII (ΦII) decreased with the leaf counting from the top downward, for different leaf position in three regions of 4 leaves were in order of the middle, the apical, the basal, but there was no significant difference among cultivars. The down-regulated energy dissipation (ΦNPQ) showed an opposite trend compared with ΦII while the non-light induced energy dissipation (ΦNO) remained stable in different cultivars and leaf position. With leaf growth, the upper leaf position showed a relatively high ΦII and low ΦNPQ while the lower leaf position showed a complicate scenario: the light excitation energy distributed between ΦII and ΦNPQ at the first two weeks with stable ΦNO, but over three weeks, ΦII, qL and Fv/Fm declined with different degree(ΦII>qL>Fv/Fm)accompanied the inclined ΦNO, which implied that the middle leaves remained constant, the upper leaves possess the high light use efficiency(LUE) while the lower leaves had low LUE significantly among different leaf position. The analysis of relative electron transport rate(rETR) in different growth time indicated that the middle of the first fully expanded leaf from the top had the maximum rETR in the same period, thus indicating that this leaf could be chosen for chlorophyll fluorescence parameters comparison and evaluation as functional leaf. The results clarified the light use efficiency distributed among canopy, thus could be used in choosing appropriate leaf position for chlorophyll fluorescence diagnosis and set up the basis for the quick evaluation of the photosynthetic potential of sugarcane canopy.
Key words Sugarcane;Chlorophyll fluorescence;Leaf position;Growth time
doi 10.3969/j.issn.1000-2561.2015.11.016
甘蔗是中国最重要的糖料作物,种植面积和总产量居世界第三,但中国甘蔗的单产较低[1]。中国甘蔗的植期以春植和秋植为主,气候变化导致西南华南地区季节性干旱频发[2],极大的影响新植蔗出苗率和幼苗的生长,因此,通过合理的光合作用指标对甘蔗品种光合潜力及耐旱性进行快速评价,是从光能利用角度对甘蔗进行灾损评估的基础性工作,也是筛选高光效甘蔗品种、提高甘蔗产量的重要途径之一。
光合作用是作物产量形成的基础,其中叶绿素荧光参数被广泛用于检测植物光系统的运转状况[3-4]。常用的叶绿素荧光参数基于对光系统两种假设得出,Puddle-model假设天线色素和光系统II(PSII)反应中心是一一对应的,以此推导出荧光猝灭qP和非光化学猝灭NPQ[5];而Lavergne和Trissl等人[6]的研究认为天线色素系统作为一个整体为PSII反应中心提供电子,Kramer等人[7]在此基础上推导出qL及两个新的叶绿素荧光参数:可调节性能量耗散(ΦNPQ)和非调节性能量耗散(ΦNO),与PSII量子效率(ΦII)一同描述激发光能的分配特性。从定义和计算结果来看,ΦPSII等同于由Puddle-model计算的PSII实际光化学效率(ΦPSII)和基于Lake-model计算的PSII量子效率(ΦII)。对于包括甘蔗在内的C4喜光作物,当光合作用处于稳态时,PSII光化学量子产额(ΦPSII)和CO2同化量子产额(ΦCO2*)的比值在一个相当大的光合有效辐射(PAR)(40~2 700 μmol/m2·s)和CO2(30~370 μL/L)变化范围内保持恒定,因作物品种而异[8]。其中ΦPSII由“开放”的光反应中心捕获电子的效率(Fm′-Fo′)/Fm′和开放的PSII反应中心所占的比例 (Fm′-Fs′)/(Fm′-Fo′)共同决定[9]。以上研究表明,通过叶绿素荧光参数估算光合作用是可行的。
前人研究表明,因叶片的光学和光化学性质不同而导致不同叶位在不同发育时间内的光合能力有所差异[10]。因此,只有了解不同叶位的叶绿素荧光参数对发育时间的动态响应,才能选择合适的叶片进行数据观测,进而实现基于叶绿素荧光参数的胁迫诊断和光合作用的评价。在大宗农作物上,选取对产量形成有绝对贡献的功能叶片作为叶绿素荧光参数的研究对象,已得到较为一致的结论,如在小麦[11]和水稻[12]上均选用旗叶来研究PSII高温伤害及生理活性对干物质积累和转运的影响,在棉花[13]上则选择对位叶研究棉铃干物质积累的影响等。而针对C3作物(棉花[14]、黄瓜[15]、小麦[16])和C4作物(玉米、高粱)[17]中不同叶位的光合或叶绿素荧光特性,也都有较为系统的研究。在甘蔗上,虽然杨丽涛等[18]和罗俊等[19]分别针对糖蔗和能源蔗不同叶位的叶片形态和光合特性进行了较为系统的研究,但尚未对叶片不同部位和不同生长时间内叶绿素荧光特性进行比较分析,因而不能对甘蔗光合能力进行快速评价,相关研究并未见报道。
甘蔗是全茎秆收获作物,不同于棉花的对位叶和稻麦的旗叶对产量形成起决定作用,因此任何单一叶片的测定均不能反映整株光合潜力。本研究通过对3个主栽品种甘蔗苗期不同叶位叶片在不同生长时间内叶绿素荧光参数的测定分析,明确不同叶位叶绿素荧光参数对不同生长时间的响应,为利用叶绿素荧光参数进行光合潜力的快速評价,进而为实现甘蔗高光效品种的鉴定与筛选和灾损评估奠定一定的理论基础。
1 材料与方法
1.1 试验材料与试验地概况
对于试验材料的选择,由于目前针对甘蔗叶绿素荧光特性研究较少,在材料选择方面缺乏参考,为了使研究结果有较好的代表性,本研究采用3个主栽品种进行试验研究。供试品种为新台糖16号(ROC16)、新台糖22号(ROC22)和桂柳05-136。
试验地点为中国热带农业科学院湛江实验站,供试土壤为红壤土,速效N、P、K和有机质分含量别为65.81 mg/kg、19.26 mg/kg、80.11 mg/kg和21.04 g/kg。栽培方式为桶栽,栽培桶规格为桶口直径40 cm、桶底直径32 cm、桶高40 cm,栽培桶参考大田甘蔗种植距离摆放,摆放密度为6桶/m2,每桶种植种茎3段,每段1个芽,每个品种种植25桶。栽培桶置于基地硬化地面上,无遮阳措施。
1.2 方法
1.2.1 试验设计 不同叶位和叶片不同部位的叶绿素荧光特性研究试验:于2014年5月12~14日试验,试验期间多云天气。于甘蔗生长至六片真叶期,选择长势一致的植株作为试验材料,每个品种3个重复,于每日上午8:00~11:00测试,每天测试1个品种,叶位选择为倒数第一、二、三、四片全展叶片,分为叶基部、叶中部和叶尖端3部分。
不同叶位叶绿素荧光参数对不同生长时间的响应试验:于2014年10月8日开始,11月5日结束,试验期间天气晴朗有时多云。选择幼苗长势一致的植株,针对不同叶位进行定株观测,每个品种3个重复,每周测量1次,叶位选择为第1~6片真叶的中部叶位,观测于每日上午8:00~11:00进行。
1.2.2 叶绿素荧光参数的测定 采用德国PAM公司生产的Mini-PAM于每日早8:00进行30 min暗适应后进行暗反应参数的测定(测试时避开主叶脉):首先给出1个微弱的非光化光,测量初始荧光(Fo);接着给出1个饱和光,测量最大荧光(Fm)。接着于上午9:00在自然光下进行30 min光适应,于9:30~11:00使用光叶夹补充人工光源测量光化光下(强度约为290 μmol/m2·s)光反应参数:光化光控制时间为90 s,测定稳态荧光(Fs);接着给出1个饱和光,测定稳态最大荧光(Fm′)。试验一和试验二测量时刻温度变化范围分别为31~34 ℃和27~29 ℃,光合有效辐射(PAR)范围为285~295 μmol/m2·s。根据Kramaer[7]公式计算出基于Lake-model的叶绿素荧光参数: PSII量子效率ΦI= (Fm′-Fs)/Fm; 光化学猝灭qL=(Fo′/Fs)(Fm′-Fs)/(Fm′-Fo′); 非调节性能量耗散ΦNO=1/[NPQ+1+qL(Fm/Fo-1)], 其中NPQ(Fm/Fm′-1)为非光化学猝灭; 可调节性能量耗散ΦNPQ=1-ΦII-ΦNO;相对电子传递速率rETR=PAR×0.84×0.5×ΦII。其中, 0.84为经验性吸光系数, 0.5为假设天线色素吸收的光能被两个光系统平均分配[20]。最大光能利用效率Fv/Fm=(Fm-Fo)/Fm;稳态最小荧光根据Oxborough和Baker[21]的公式计算得出Fo′=Fo/[(Fv/Fm)+(Fo/Fm′)]。
1.3 数据处理
采用SPSS统计分析软件中Duncan多重比较分析方法进行统计分析。
2 结果与分析
2.1 光系统实际光化学效率在品种间、叶位间和叶片不同部位间的变化
2.1.1 相同品种和叶位叶片不同部位ΦII的变化 ΦII反映限定光强下PSII的量子效率,可用来衡量PSII实际光化学效率[3]。对不同叶片部位的ΦII的分析表明,除了ROC16倒二叶的叶尖端大于叶中部(但并无显著差异)之外,3个供试品种不同叶位的叶片均表现为叶中部>叶基部>叶尖端。ROC22除倒四叶之外的叶片差异不显著;ROC16叶基部与另外两个部位差异显著;桂柳05136则表现出各部位间倒三和倒四叶差异较为显著。ROC16叶基部的ΦII是3个品种中最低的,而叶尖端的ΦII则是3个品种中最高的(图1)。
2.1.2 相同叶位和叶片部位不同品种ΦII的变化 对相同叶位和叶片部位不同品种间的ΦII分析结果表明,3个供试品种之间的差异均表现为不显著(图2)。
2.1.3 相同品种和叶片部位不同叶位ΦII的变化 对各个供试品种叶片相同部位不同叶位的ΦII进行比较,结果显示3个供试品种不同叶位间基本随着倒数叶位的增加而减小。对于ROC22的叶基部和叶中部,倒四叶与其他3片叶差异显著,叶尖端4个叶位差异不显著;ROC16叶基部和叶中部不同叶位间差异不显著,叶尖端表现为倒一倒二叶大于倒三倒四叶;桂柳05136则表现出中部叶位的倒四叶与其他3片差异显著,叶基部4个叶位无显著差异,叶尖端倒三叶和倒四叶显著下降(图3)。
2.1.4 ΦII在品种间、叶位间和叶片不同部位间的变化 对品种、叶片部位、叶位的差异显著性分析表明,3个供试品种间差异不显著;叶片部位表现为叶中部>叶尖端>叶基部,差异显著;叶位表现为倒一>倒二>倒三>倒四,倒一倒二叶与倒三倒四叶之间差异显著(图4)。
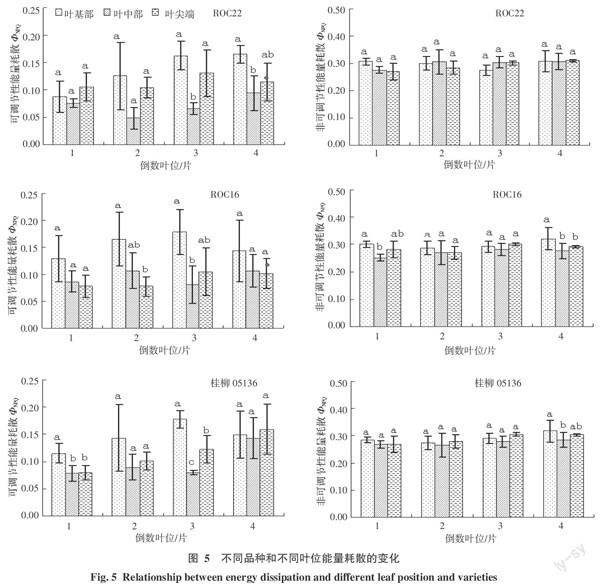
2.2 光系统能量耗散在品种间、叶位间和叶片不同部位间的变化
光系统对能量耗散的水平可以反映植物光保护能力的强弱,叶绿素荧光参数中非光化学猝灭是反映这一水平的指标。基于Lake-model计算出的非光化学猝灭分为反映光系统中可调节性的能量耗散部分(ΦNPQ)和其他非光化学猝灭途径不可调节的能量耗散部分(ΦNO)[7]。研究表明,对于ROC22和桂柳05136,ΦNPQ的总体趋势表现为随着倒数叶位的增加而递增,叶片部位间变化为叶基部>叶尖端>叶中部;而ROC16不同叶位的ΦNPQ则相对稳定,且叶片不同部位间仍以叶基部能量耗散水平最强,叶尖端>叶中部(倒三叶除外)。除桂柳05136的倒四叶、ROC22的倒一和倒四叶之外,ΦNO在各个叶片部位之间的差异不显著,且随着叶位的递增并无明显变化(图5)。相同叶位和叶片部位,不同品种之间ΦNPQ和ΦNO均表现为差异不显著(数据未列出)。
2.3 不同叶位叶绿素荧光特性对不同生长时间的响应
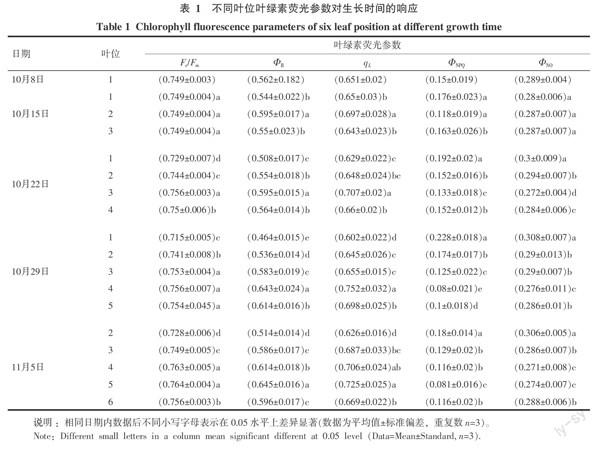
通过不同叶位和叶片不同部位的葉绿素荧光特性研究试验,表明叶绿素荧光参数在品种间、叶片部位间和叶位间的差异以叶片部位间最显著,其中叶片中部实际光化学效率最高,而叶绿素荧光参数在3个供试品种间的变化规律趋于一致,且差异不显著。因此,在叶绿素荧光参数对不同生长时间的响应试验中,选取3个品种中部叶位共9张叶片的平均值做比较分析。
基于Lake-model计算的PSII量子效率(ΦII)、可调节性能量耗散(ΦNPQ)和非调节性能量耗散(ΦNO)共同描述PSII中激发能分配特性(ΦII+ΦNPQ+ΦNO=1)[7]。结果表明:ΦII随着正数叶位的增加呈现出上升趋势,以观测叶片的倒二叶(即叶鞘完全展开的倒一叶)最大;正一叶和正二叶的ΦII随着时间的推移而递减,正三叶基本维持恒定,正四叶和正五叶则呈现上升趋势(正四叶由10月29日至11月5日略有下降)。
ΦNPQ则刚好表现出与ΦII完全相反的变化趋势;除正一叶和正二叶的ΦNO分别在10月29日和11月5日略有增加,其余叶位的的ΦNO在不同生长时间内并无显著变化。正一叶和正二叶在展叶初的2周ΦNPQ的上升弥补了ΦII的下降,但是随着时间的持续推移ΦNO逐渐上升;3周后的正一叶和正二叶的ΦII分别降了9.8%和8.1%,ΦNPQ和ΦNO则分别增加7.8%、6.2%和1.9%、2%。正三叶到正六叶则保持相对较高的ΦII和较低的ΦNPQ,而ΦNO则基本保持不变(表1)。
基于Lake-model计算出的光化学猝灭qL描述处于开放状态的PSII反应中心所站比例[7],Fv/Fm用来衡量PSII潜在最大光能利用效率[3]。qL和Fv/Fm的变化趋势均与ΦII趋于一致,但是变化幅度为ΦII>qL>Fv/Fm(以正一叶为例,ΦII、qL和Fv/Fm分别下降了9.8%、4.9%和3.4%)。以上结果说明,上部叶片具备较高的实际光化学效率,而下部叶片则表现为实际光化学效率下降,同时伴随着能量耗散的增加,表明植株叶片是通过增加能量耗散和降低PSII反应中心的开放程度来防止多余的光能造成的损伤。
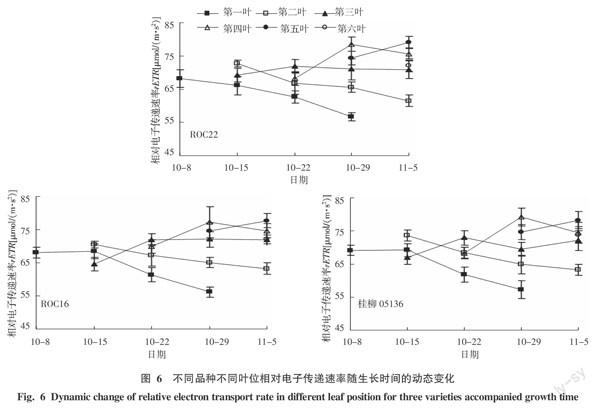
2.4 3个品种不同叶位线性电子传递速率的动态变化
rETR用来描述PSII中线性电子传递速率,即PSII在额定光强下的单位量子产额,反映PSII光能转化能力[3]。对叶片中部位点的rETR在不同生长时间内动态变化的研究表明,3个供试品种的正一叶和正二叶均呈现出下降趋势,且正一叶下降幅度大于正二叶;正三叶的rETR较为稳定;正四叶和正五叶分别表现为先上升后下降和上升趋势。对于每次测量rETR的最大值出现在测量当天的倒数第二叶位,而此时的倒数第一叶位为新发出叶片,叶鞘并未完全展开(图6)。因此,最上部的全展叶的中部叶位具备最大的的相对电子传递速率,代表植株最大光能转化能力,可选做进行叶绿素荧光特型横向比较的叶位。
3 讨论
3.1 叶位对不同甘蔗品种叶绿素荧光特性的影响
C4植物光合碳同化途径从光系统形态结构和生化功能角度进行适应性调整(如利用苹果酸循环浓缩可利用的CO2,消除加氧反应以降低光呼吸),补充完善C3途径,以应对强光、高温和干旱的气候条件[22]。在成熟的C4叶片中,消除加氧反应使得CO2同化成为线性电子流的主要去路,在很大的光强范围内,PSII实际光化学效率和CO2同化速率线性相关[23]。甘蔗品种的选择需要同时考虑高光效和适应性,ROC16和ROC22从定植至伸长初期光合速率并无显著差异[24],这与本研究得出的ΦII在品种间差异不显著的结论相吻合,且桂柳05136在苗期的光合活性与两个ROC品种处于同等水平。而引种试验报告表明,ROC22和ROC16的产量和蔗糖分分别为99.63 t/hm2、90.66 t/hm2和14.15%、14.49%[25]。这说明3个主栽品种在经济性状上的差异并非叶片光合潜力的影响造成的,而是可能与干物质分配和品种对环境的适应性等因素有关。
王燕鹏等人[26]对玉米C4光合叶不同部位解剖结构和光抑制特性的比较分析认为,维管束鞘和细胞叶绿体数量、光合速率及光抑制的恢复能力均表现为中部>顶部>基部,我们的试验结果表明ΦII对叶位的响应总体表现为叶中部>叶尖端>叶基部,这与王燕鹏等[26]的研究趋于一致。但是对于3个甘蔗供试品种,ROC22相同叶位不同叶片部位间差异不显著,而对于ROC16和桂柳05136,个别叶位出现叶尖端与中部叶位持平或大于中部叶位的现象,这可能是由于相对玉米叶片,甘蔗叶片短、窄、直挺的甘蔗基因型冠层辐射特征参数大(包括中下部叶位),有利于光能的吸收和利用[27]。
PSII转化的能量除了供给电子传递之外,还用于进行能量耗散。对玉米光合叶的研究表明,叶绿素a/b的含量随着距叶耳距离的增加而减小,且叶基部最易发生光抑制现象且修复能力差[28],这也解释了本研究得出的ΦNPQ表现为叶基部最大的结论。同时,虽然玉米叶片不同部位叶绿素含量差异显著,但是正常光照条件下,相同位点PSII最大光化学效率之间并无显著差异[26],这说明叶片不同位点叶绿素中PSII均保持完好的功能。而安东升等人[29]对基于Lake-model的叶绿素荧光参数的研究进一步表明,甘蔗叶片PSII处于正常状态和对胁迫的应激反应状态时,在不同光温背景下ΦNO会表现出相对稳定,ΦNPQ与ΦII刚好呈现出相反的变化趋势,这同样可以解释本研究中ΦNO在不同品种叶位和不同的叶片部位间差异总体上差异并不显著,而ΦNPQ与ΦII刚好呈现出相反的变化趋势的原因。
3.2 生长时间对不同叶位叶绿素荧光特性的影响
对玉米和高粱不同叶位SPAD的研究表明,随着发育进程的推进,下部叶位SPAD的值呈递减趋势,中部叶位则表现为先上升后下降,而上部叶位则出现上升趋势,这是由于不同叶片由于生长发育的先后顺序不同所致[30]。本研究对于苗期生长时间不同叶位相对电子传递速率变化的研究表明,正一叶和正二叶的rETR呈下降趋势,正三叶和正四叶基本呈先上升后下降的趋势,正五叶呈上升趋势。这与前人对不同叶位SPAD随发育进程的演变相一致,即不同生长时间内,叶片传递电子能力的大小与不同叶位的叶绿素含量有关[30]。且对于正一叶和正二叶在展叶一周至两周,叶片中入射光的能量分配在ΦII和ΦNPQ之间进行;而第三周后,ΦNO开始逐渐上升,并伴随着qL和Fv/Fm用的下降。其中qL的变化幅度小于ΦII的变化幅度,说明在限定光强下上部叶位的PSII仍能维持较高的开放程度,而此时中下部叶位表现出的非调节性能量耗散增加和潜在最大光能利用效率降低的现象,可能是由于N素的再分配而导致的叶绿素含量的降低,从而引起的光抑制现象所致。
参考文献
[1] 李如丹, 張跃彬, 刘少春,等. 国内外甘蔗生产技术现状和展望[J]. 中国糖料, 2009(3): 54-56.
[2] 刘定辉, 刘永红, 熊 洪,等. 西南地区农业重大气象灾害危害及监测防控研究[J]. 中国农业气象, 2011, 32(add1): 208-212.
[3] Papageorgiou G C, Govindjee(eds). Chlorophyll a Fluorescence: A Signature of Photosynthesis[M]. Springer-Verlag, New York Inc, 2005.
[4] Suresh K, Nagamani C, Ramachandrudu K, et al. Gas-exchange characteristics, leaf water potential and chlorophyll a fluorescence in oil palm(Elaeis guineensis Jacq.)seedlings under water stress and recovery[J]. Photosynthetica, 2010, 48(3): 430-436.
[5] Krause G, Vernotte C, Briantais J. Photo-induced quenching of chlorophyll fluorescence in intact chloroplasts and algae resolution into two components[J]. Biochim Biophys Acta, 1982, 679: 116-124.
[6] Lavergne J, Trissl H W. Theory of fluorescence induction in Photosystem II: derivation of analytical expressions in a model including exciton-radical-pair equilibrium and restricted energy transfer between photosynthetic units[J]. Biophysical journal, 1995, 68: 2 474-2 492.
[7] Kramer D M, Johnson G, Kiirats O, et al. New fluorescence parameters for the determination of QA redox state and excitation energy fluxes[J]. Photosynthesis Research, 2004, 79: 209-218.
[8] Edwards G E, Baker N R. Can CO2 assimilation in maize leaves be predicted accurately from chlorophyll fluorescence analysis[J]. Photosynthesis Research, 1993, 37: 89-102.
[9] Genty B, Birantais J, Baker N R. The relationship between the quantum efficiencies of photo-systems I and II in pea leaves[J]. Biochem Biophys Acta, 1989, 990: 87-92.
[10] Filippo B, Martina P, Chiara C, et al. Reto JS(2011) Conclusive remarks. Reliability and comparability of chlorophyll fluorescence data from several field teams[J]. Environmental and Experimental Botany, 2011(73): 116-119.
[11] 郭翠花, 高志强,苗果园. 花后遮阴对小麦旗叶光合特性及籽粒产量和品质的影响[J]. 作物学报, 2010, 36(4): 673-679.
[12] 杨卫丽, 黄福灯,曹珍珍, 等. 高温胁迫对水稻光合PSII系统伤害及其与叶绿体D1蛋白间关系[J]. 作物学报, 2013, 39(6):1 060-1 068.
[13] 胡 宏, 张文静, 陈兵林, 等. 棉铃对位叶C/N的变化及其与棉铃干物质积累和分配的关系[J]. 作物学报, 2008, 34(2):254-260.
[14] 李志博, 魏亦农, 张荣华,等. 棉花不同叶位叶绿素荧光特性初探[J]. 棉花学报, 2005, 17(3): 189-190.
[15] 李 伟, 眭晓蕾, 王绍辉, 等. 黄瓜幼苗不同叶位叶片光合特性对弱光的响应[J]. 中国农业科学, 2008, 41(11): 3 698-3 707.
[16] 谭雪莲, 郭文天, 张国宏,等. 氮素对小麦不同叶位叶片叶绿素荧光参数的调控效应[J]. 麦类作物学报, 2009, 29(3): 437-441.
[17] 姜 英, 林叶春, 许和水, 等. 两种C4作物不同叶位光合及叶绿素荧光特性比较[J]. 中国农业大学学报, 2012, 17(3): 34-42.
[18] 杨丽涛, 陈超君, 李杨瑞. 田间生长条件甘蔗光合特性的研究[J]. 西南农业学报, 2001, 14(7): 69-74.
[19] 罗 俊, 张 华, 陈由强, 等. 能源甘蔗不同叶位叶片形态、光合气体交换及其与产量关系[J]. 应用与环境生物学报, 2006, 12(6): 754-760.
[20] Schreiber U, Bilger W, Neubauer C. Chlorophyll fluorescence as a non intrusive indicator for rapid assessment of in vivo photosynthesis. In: Schulze E-D, Caldwell M(Eds.) Ecophysiology of Photosynthesis[M]. Berlin: Springer-Verlag, 1994, 100: 49-70.
[21] Oxborough K, Baker N. R. Resolving chlorophyll a fluorescence images of photosynthetic efficiency into photochemical and non-photochemical components-calculation of qP and Fv/Fm without measuring Fo′[J]. Photosynthesis Research, 1997, 54: 135-142.
[22] Sage R. F. The evolution of C4 photosynthesis[J]. New Phytologist, 2004, 161: 341-370.
[23] Baker N. R. Chlorophyll Fluorescence: A Probe of Photosynthesis In Vivo[J]. Annual review of plant biology, 2008, 59: 89-113.
[24] 唐仕云, 劉海斌, 贤 武. 甘蔗新品种(系) 种性与光合特性的相关关系[J]. 中国糖料, 2008(3): 7-9.
[25] 周 会, 雷敬超, 王伦旺, 等. 2008~2009年广西甘蔗品种区域试验报告[J]. 南方农业学报, 2012, 43(3): 306-310.
[26] 王燕鹏, 崔震海, 朱延姝, 等. 玉米C4光合叶不同部位解剖结构和光抑制特性的比较[J]. 植物生理学报, 2012, 48(6): 571-576.
[27] 罗 俊, 张 华, 邓祖湖,等. 甘蔗不同叶位叶片形态与冠层特征的关系[J]. 应用与环境生物学报, 2005, 11(1): 28-31.
[28] 刘 畅, 崔震海, 张立军, 等. 玉米C4光合叶不同部位“花环”结构及叶绿素含量的变化[J]. 玉米科学, 2012, 20(6): 60-62, 69.
[29] 安东升, 曹 娟, 黄小华,等. 基于Lake模型的叶绿素荧光参数在甘蔗苗期抗旱性研究中的应用[J]. 植物生态学报, 2015, 39(4): 398-406.
[30] 黄瑞冬, 王进军,许文娟. 玉米和高粱叶片叶绿素含量及动态的比较[J]. 杂粮作物, 2005, 25(1): 30-31.
猜你喜欢
农业与技术(2023年11期)2023-06-19
儿童故事画报·自然探秘(2022年12期)2022-11-24
干旱地区农业研究(2022年4期)2022-07-28
四川蚕业(2022年1期)2022-06-06
基层中医药(2021年3期)2021-11-22
特别健康(2018年3期)2018-07-04
山东农业科学(2017年2期)2017-03-15
中国糖料(2016年1期)2016-12-01
西南农业学报(2016年5期)2016-05-17
现代园艺(2016年2期)2016-03-15
