
应用免疫冷冻超薄切片技术观察罗非鱼肠道M样细胞
2015-10-20 00:59李莉萍等
江苏农业科学 2015年9期
关键词:罗非鱼
李莉萍等
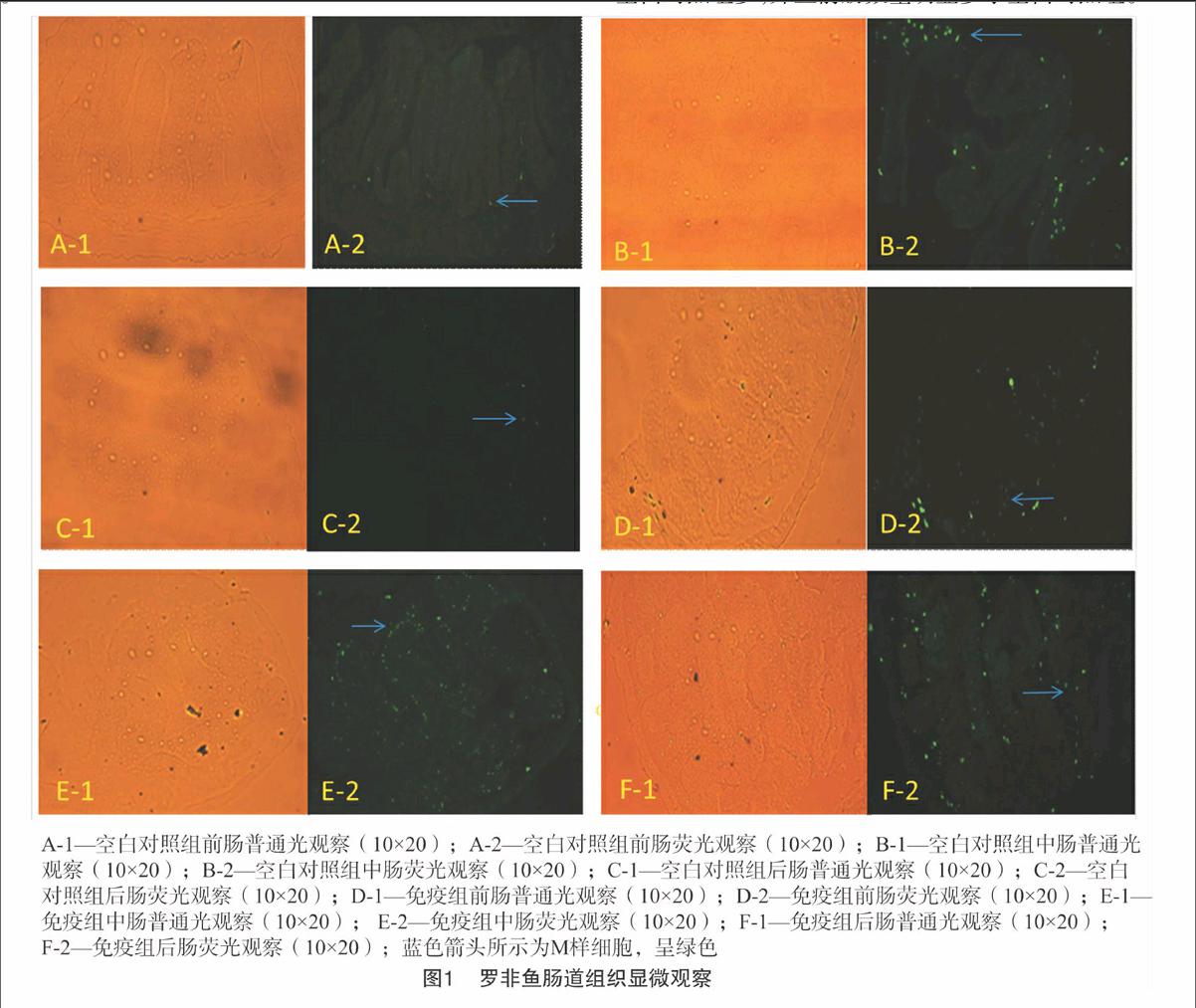
摘要:为鉴定罗非鱼肠道M样细胞及各肠道分布情况,利用FITC标记的荆豆凝集素(UEA-1)对罗非鱼前、中、后肠冰冻切片进行直接免疫荧光显微观察。结果发现,在罗非鱼前、中、后肠均观察到M样细胞;中肠和后肠M样细胞数量比前肠明显多。免疫组M样细胞数量显著比空白对照组多,且免疫组前肠数量明显多于空白对照组前肠。研究表明,在罗非鱼前、中、后肠存在M样细胞,肠道接触无乳链球菌弱毒抗原之后,在数量、分布上均发生变化,初步表明M样细胞参与肠道免疫应答。
关键词:罗非鱼;M细胞;荆豆凝集素
中图分类号: S917;S942.5文献标志码: A文章编号:1002-1302(2015)09-0264-03
通信作者:陈明,博士,副研究员,主要从事水产疫苗与免疫技术研究。E-mail:cm990919@163.com。近年来,链球菌病对我国罗非鱼养殖危害日趋严重,药物治疗不能有效控制该病,反而易产生耐药性,污染环境,同时还存在食品安全隐患[1]。而疫苗预防是控制该病最有效方法之一。其中注射疫苗免疫效果好,保护期长,但操作麻烦,在实际生产中难以大面积推广应用[2]。口服疫苗操作方便、对鱼体伤害小,适合大规模鱼集体免疫,但其免疫保护率低,保护期短[3],其原因之一可能是罗非鱼肠道黏膜细胞摄取、吸收和呈递抗原有限。因此,有必要对鱼类肠道黏膜抗原递呈进行研究。
在肠道黏膜上皮内有一种特殊类型的M样细胞,也叫微皱褶细胞(microfold cell)[4-5],它是肠道免疫组织吸收微生物或抗原的主效应细胞,是激发肠道局部免疫反应首要门户。目前普遍认为,M样细胞能有效地摄取和转运肠腔内各种大分子和抗原至上皮下集合淋巴结的免疫细胞,并将抗原进一步加工和提呈,从而激活特异性B淋巴细胞和T淋巴细胞。而近年来有关肠道免疫反应研究表明,绝大部分口服疫苗都需经由M样细胞吸收,它是许多细菌和病毒致病以及黏膜疫苗作用关键[6],其摄取效率决定疫苗免疫效果。目前在人、鸡、兔、鼠、猪、牛、猴、犬等动物与淋巴组织相关联的表面上皮中均发现有M样细胞存在[7-8]。在鱼类只见虹鳟报道有M样细胞[9],其他未见报道。
本试验利用FITC标记荆豆凝集素(UEA-1)对罗非鱼前、中、后肠道冰冻切片进行直接免疫荧光染色显微观察,寻找罗非鱼肠道内形态上类似于哺乳动物的M样细胞及分布情况,为下一步研究罗非鱼肠道M样细胞摄取和呈递抗原作用机理及无乳链球菌口服疫苗研发提供理论依据。
1材料与方法
1.1材料
1.1.1试验鱼试验鱼来自广西水产科学研究院罗非鱼良种场的吉富罗非鱼,平均体质量250~300 g,分成2组,每组20尾,暂养于600 L大缸桶中,24 h充氧,在28±5 ℃条件下饲养2周,使之适应饲养环境。每3 d换水2/3。每天饲喂2次(09:00和16:00),日投喂量占鱼体质量的2%。
1.1.2试验试剂Lectin from ulex europaeus(FITC conjugate)购自Sigma公司;胎牛血清购自浙江天杭生物科技有限公司;DMEM培养基购自Gibco公司;血平板河南购自郑州安图绿科生物工程有限公司;TSB培养基购自杭州微生物试剂有限公司;无乳链球菌弱毒菌株YM001为本实验室筛选获得;其他生物、化学试剂均为从商业公司购买的分析纯产品。
1.2实验方法
1.2.1疫苗制备从-80 ℃冰箱中取出无乳链球菌弱毒菌株YM001在血平板上划线复苏,24 h后转到TSB培养基中扩大培养。取120 mL菌液10 000 r/min离心10 min,弃上清,PBS定容至20 mL,用喷壶均匀喷至120 mL饲料。晾干 5 min,投喂免疫组;空白对照组用不添加弱毒株饲料饲喂。饲喂3 h后采样。
1.2.2取样先取空白对照组鱼肠道,然后再取免疫组鱼肠道。将活鱼剖腹后,迅速取肠的前、中、后段,约4 cm左右。前段在胃后端至肠的第1个弯曲之间的中间取样;中段在肠的第1个弯曲至最后1个弯曲的中间取样;后段在最后1个弯曲至肛门的中间取样[10]。
1.2.3冰冻切片制备样品用PBS冲洗3次,滤纸吸干;迅速置于4 ℃预冷4%多聚甲醛溶液中1 h。用含10% FBS细胞培养液冲洗3次,滤纸吸干;立即放在液氮中进行冷冻1 h以上。取样品修成0.3 cm×0.3 cm,厚度0.3 cm小块,用OCT在金属锡箔盒内进行包埋,把包埋组织置于冰冻切片机内,Leica CM3050冰冻切片机横切,厚为5 μm,用防脱落载玻片粘片,置于4 ℃保存。
1.2.4免疫荧光染色与观察将冰冻切片置于37 ℃的FITC-UEA-1溶液(1 ∶80)避光作用1 h后,PBS冲洗3次,每次5 min,滴加少量50%的PBS甘油溶液封片,置于荧光显微镜下检测,肠道上皮组织内出现绿色荧光的判定为M样细胞。
2结果与分析
由图1可知,在免疫组与空白对照组罗非鱼前、中、后肠均观察到M样细胞;从数量上看,不管是免疫组,还是空白对照组,中肠和后肠M样细胞比较多,前肠比较少;并且M样细胞主要集中在中肠和后肠。免疫之后,M样细胞数量明显比空白对照组多,并且前肠数量明显多于空白对照组。
3讨论
抗原成功黏附并被 M 样细胞吸收是其诱发黏膜免疫反应关键,这种作用主要依赖于细菌表面凝集素与M样细胞表面表达的糖基相互作用。M样细胞膜表面覆盖着糖蛋白中的岩藻糖,可以被荆豆凝集素(UEA-1)特异性识别并结合[11-12],因此有人提出UEA-1可以作为鉴定M样细胞特异指标[13-15]。陈洁等分别利用FITC和胶体金颗粒(10 nm) 标记的UEA-1对正常小鼠小肠Peyers集合淋巴小结滤泡相关上皮中M样细胞形态结构进行观察,通过荧光显微镜和免疫透射电镜观察,证明UEA-1可特异性地识别M样细胞[16]。以前的报道称在硬骨鱼类上没有M样细胞,但最近研究发现在鲑鱼中肠的第2肠段部分存在与哺乳动物M细胞相似的M样细胞。Fuglem等应用FITC标记的UEA-1和rho-damine标记的WGA分别对虹鳟中肠的第2肠段细胞进行观察,结果发现仅UEA阳性细胞具有哺乳动物M样细胞特征[9]。本研究也采用FITC结合UEA-1特异方法检测M样细胞,成功从罗非鱼肠道中观察到M样细胞,该结果有力地证明罗非鱼前、中、后肠道中存在M样细胞。试验整个过程只需4 h,相对于采用电镜、胶体金等方法鉴定组织中细胞,具有操作快,步骤简单,组织抗原保存良好,敏感度高、特异性强,结果可靠等优点[17]。
鱼类肠道上皮没有类似哺乳动物Peyer氏淋巴集结,但是有着相当多的淋巴细胞,研究表明鱼类淋巴细胞主要分布在肠道中后部[18]。陈洁等用伤寒减毒活疫苗Ty21a免疫小鼠后观察发现该疫苗可诱导M样细胞数量增加[19]。徐佳用免疫荧光方法检测口服Ag85A DNA疫苗在小鼠肠道M细胞表达,发现Ag85A DNA疫苗在小鼠肠道局部M细胞有表达,并且该DNA疫苗可以被肠道M细胞摄取[20]。黄玉章等研究黄芪多糖对罗非鱼肠绒毛形态结构及肠道免疫细胞的影响时发现,同一组不同肠段上皮内淋巴细胞数量中肠最多,后肠次之,前肠最少[21]。此外,在同一组的不同肠段中,罗非鱼肠道内黏液细胞的数量顺序为中肠>后肠>前肠。在大西洋鲑中,普遍认为前肠主要执行消化功能,而后肠才是主要抗原摄取以及免疫应答部位[22]。在舌齿鲈肠道中,越是靠近肛门,淋巴细胞数量就越多,表明其后肠具有更高免疫相关性[23]。Companjen等也证实了鱼类后肠为抗原的主要吸收和加工部位,抗原可在后肠被巨噬细胞吞噬,诱导产生免疫应答[24]。本研究采用无乳链球菌弱毒株拌料投喂罗非鱼,发现其肠道在接触抗原之后,M样细胞在数量、分布上均发生了明显变化,免疫组M样细胞数量比空白对照组增多,并且免疫组前肠数量明显多于空白对照组前肠,初步表明M样细胞参与免疫应答过程,但目前诱导M样细胞数量升高的机制尚不清楚,还有待于进一步探索。从分布上看,无论是免疫组还是空白对照组,M样细胞主要分布在中肠和后肠,推测罗非鱼前肠主要对外来物质进行初步识别消化,中后肠主要对抗原进行摄取及产生免疫应答。因此,本试验结果为深入研究罗非鱼肠道免疫机理及口服疫苗开发提供理论基础。
参考文献:
[1]李莉萍,王瑞,黄婷,等. 广西罗非鱼链球菌病流行菌株PCR鉴定和PFGE基因型分析[J]. 水产学报,2013,37(6):927-935.
[2]徐增辉,陈汉忠,陈明,等. 海豚链球菌疫苗对罗非鱼免疫功能的影响[J]. 大连水产学院学报,2008,23(6):413-418.
[3]Vandenberg G W. Oral vaccines for finish academic theory or commercial reality?[J]. Anim Health Res Rev,2004,5(2):301-304.
[4]Ei Bahi S,Caliot E,Bens M,et al. Lymphoepithelial Interactions trigger specific regulation of gene expression in the M cell-containing follicle-associated epithelium of Peyers patches [J]. J Immunol,2002,168(8):3713-3720.
[5]Kitagawa H,Hosokawa M,Takeuchi T,et al. The cellular differentiation of M cells from crypt undifferentiated epithelial cells into microvillous epithelial cells in follicle-associated epithelia of chicken cecal tonsils [J]. J Vet Med Sci,2003,65(2):171-178.
[6]赵燕,王霞,蔡美英. M细胞——启动黏膜免疫应答的入口[J]. 微生物学免疫学进展,2001,29(2):89-92.
[7]Fujimura Y,Iida M. A new marker for cup cells in the rabbit small intestine:expression of vimentin intermediate filament protein[J]. Medical Electron Microscopy,2001,34(4):223-229.
[8]Neutra M R,Frey A,Kraehenbuhl J P. Epithelial M cells:gate ways for mucosal infection and immunization[J]. Cell,1996,86(3):345-348.
[9]Fuglem B,Jirillo E,Bjerks I,et al. Antigen-sampling cells in the salmonid intestinal epithelium [J]. Developmental and Comparative Immunology,2010,34(7):768-774.
[10]房慧伶,曾文宗. 尼罗罗非鱼肠道的细微结构及组织化学的研究[J]. 广西农业大学学报,1992,11(3):121-126.
[11]Man A L,Prieto-Garcia M E,Nicoletti C. Improving M cell mediated transport across mucosal Barriers:do certain bacteria hold the keys?[J]. Immunology,2004,113(1):15-22.
[12]Mills D J,Tuohy K M,Booth J,et al. Dietary glycated protein modulates the colonic microbiota towards a more detrimental composition in ulcerative colitis patients and non-ulcerative colitis subjects [J]. J Appl Microbiol,2008,105(3):706-714.
[13]Clark M A,Jepson M A,Simmons N L,et al. Differential expression of lectin-binding sites defines mouse intestinal M cell[J]. J Histochem Cytochem,1993,41(11):1679-1687.
[14]Falk P,Roth K,Gorden J I. Lectins are sensitive tools for defining the differentiation programs of epithelial cell lineages in the developing and adult mouse gastrointestinal tract[J]. Am J Physiol,1994,266(6 Pt 1):G987-G1003.
[15]陈洁,高杰英,常昕,等. 正常小鼠小肠微皱褶细胞的形态学观察[J]. 免疫学杂志,2002,18(4):246-249.
[16]陈洁,高杰英,常昕,等. 应用免疫冷冻超薄切片技术观察小鼠小肠微皱褶细胞[J]. 上海免疫学杂志,2003,23(1):10-12.
[17]张翠薇,刘勇,张旭,等. 肾活检标本冰冻切片与石蜡切片免疫荧光染色结果的比较[J]. 泸州医学院学报,2013,36(1):31-34.
[18]Press C M,Evensen O. The morphology of the immune system in teleost fishes[J]. Fish Shellfish Immunology,1999,9(4):309-318.
[19]陈洁,高杰英,彭虹,等. 伤寒减毒活疫苗Ty21a免疫后小鼠小肠微皱褶细胞数量变化的观测[J]. 上海免疫学杂志,2002,22(2):102-106.
[20]徐佳. 口服Ag85A DNA疫苗在小鼠肠道M细胞的表达[J]. 沈阳医学院学报,2010,12(4):197-198.
[21]黄玉章,林旋,王全溪,等. 黄芪多糖对罗非鱼肠绒毛形态结构及肠道免疫细胞的影响[J]. 动物营养学报,2010,22(1):108-116.
[22]Vervarcke S,Ollevier F,Kinget R,et al. Mucosal response in African catfish after administration of Vibrio anguillarum O2 antigens via different routes[J]. Fish Shellfish Immunol,2005,18(2):125-133.
[23]Abelli L,Picchietti S,Romano N,et al. Immunohistochemistry of gut-associated lymphoid tissue of the sea bass Dicentrarchus labrax (L.) [J]. Fish Shellfish Immunology,1997,7(4):235-245.
[24]Companjen A R,Florack D E,Bastiaans J H,et al. Development of a cost-effective oral vaccination method against viral disease in fish[J]. Dev Biol,2005,121:143-150.
猜你喜欢
当代水产(2023年2期)2023-05-19
当代水产(2022年6期)2022-06-29
今日农业(2021年15期)2021-11-26
当代水产(2021年7期)2021-11-04
当代水产(2020年3期)2020-06-15
当代水产(2019年4期)2019-05-16
当代水产(2018年8期)2018-11-02
当代水产(2018年12期)2018-05-16
农家之友(2018年12期)2018-03-12
农家科技中旬版(2016年12期)2016-04-16
