
自主跑轮运动对C57BL/6小鼠骨骼肌线粒体蛋白输入(PIM)组件的影响
2015-10-18 10:11王海燕丁树哲
体育科学 2015年2期
张 喆,崔 迪,王海燕,孙 易,丁树哲
线粒体是真核细胞内重要的细胞器,关键的能量代谢“工厂”。线粒体有一定的特殊性,其具有基因半自主性,即线粒体蛋白的合成受细胞核基因(nDNA)和线粒体基因(mtDNA)的双重调控。大部分线粒体蛋白需由nDNA编码,随后在细胞质中经历转录、翻译、修饰等过程,再通过存在于线粒体膜上的蛋白输入机制(protein import machinery,以下简称PIM)进入线粒体的不同区域,从而行使其特定职能[20]。因此,线粒体PIM对于线粒体、细胞、乃至个体的健康和稳态都至关重要。
PIM组件主要包括线粒体外膜转位酶/线粒体内膜转位酶(TOM/TIM)复合物,超过半数的线粒体蛋白输入选择此途径。其中TOM复合体主要包括Tom 70,Tom 40,Tom 22等,TIM 复合体则 主要有 Tim22,Tim23等[6]。除此之外,已有研究者对线粒体PIM进行了大量的基础性研究,绘制了线粒体蛋白输入路线图[2],研究大多集中于PIM经典路线,但关于线粒体PIM的应用性探讨和PIM非经典路线的研究较少。因此,基于前期的基础研究成果,本研究拟针对线粒体PIM组件在运动方面的应用性研究进行初探,同时,还关注了运动对线粒体PIM非经典路线组件的影响。
1 研究材料与方法
清洁级C57BL/6雄性小鼠20只,4周龄,体重24~27g,购于上海斯莱克动物实验中心,实验动物生产许可证号:SCXK(沪)2007-0005;实验动物使用许可证号:SYXK(沪)2004-0001。国家标准啮齿类动物常规饲料及垫料均由上海斯莱克实验动物有限公司提供。饲养期间,小鼠自由摄食、饮水,动物房温度为22±1℃,相对湿度为50%±10%,明暗周期12∶12,遵循小鼠生物节律。
购入小鼠适应性喂养1周后,随机分为2组:对照组(N组,n=10)和自主跑轮运动组(E组,n=10)。运动方案:E组小鼠采用为期8周的自主跑轮运动,每日进行跑轮圈数的记录与清零。实验结束后,过夜禁食12h,于次日麻醉断颈处死,取完整的左、右股四头肌置于液氮中速冻,随后移入-80℃超低温冰箱保存待用。
1.1 指标检测
1.1.1 相关指标转录水平的检测
用TRIZOL法提取总RNA。取一侧股四头肌约100 mg剪碎,加入TRIZOL1mL提取骨骼肌RNA,使用超微分光光度计测OD260/OD280比值,以此检验纯度;琼脂糖凝胶电泳检测RNA完整性,将提取的RNA用TOYOBO FSQl01试剂盒反转录为cDNA,随后使用实时荧光定量PCR仪检测P53(Protein 53)、Bip、HSPA1、Tom 20、Tom 40、Tim23、Tim22、VDAC3的 mRNA水平。实验所用引物由上海生物工程有限公司合成(表1)。

表1 本研究目标基因引物序列一览表Table 1 The Target Gene Primers
1.1.2 相关指标蛋白表达水平的检测
取另一侧股四头肌约100mg剪碎,加入1mLRIPA裂解液磁珠匀浆,离心取上清,用BCA法进行蛋白定量,用裂解液将各样品的蛋白浓度调平后,加入SDS-PAGE蛋白上样线冲液(5×),蛋白变性后进行SDS-PAGE电泳,转膜至PVDF膜上。随后用5%脱脂牛奶TBST室温封闭1h,洗膜后加入相应一抗(Tom 40抗体购于Santa Cruz、Tim22抗体购于abcam、HSPA1抗体购于abcam、P53抗体购于Santa Cruz,GAPDH抗体购于杭州贤至,tubline抗体购于Santa Cruz)于4℃孵育过夜。次日TBST洗膜3次,每次10min,加入二抗(购于碧云天)室温孵育2h,再次洗膜3次,用ECL显色和曝光,用目的蛋白条带与内参(GAPDH或tubline)的灰度值之比反映目的蛋白表达量。
1.2 统计学分析
所有数据均用平均数±标准差表示,用GraphPad Prism 5.0分析,组间采用独立样本T检验,以P<0.05为显著性差异,P<0.01为极显著性差异。
2 结果
2.1 小鼠的体重变化
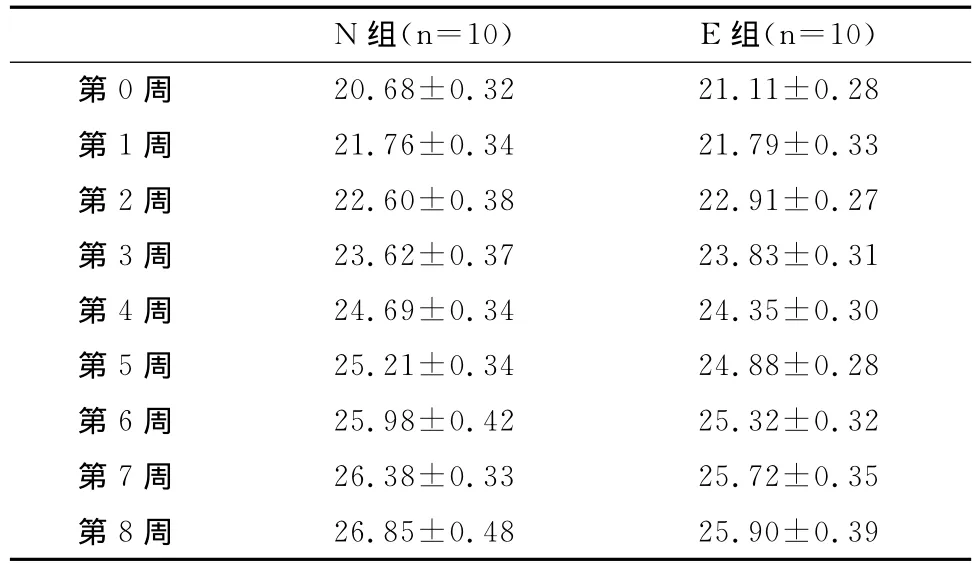
表2 本研究N组与E组小鼠的体重变化一览表Table 2 The Changes of the Body Weight in Group N and Group E
2.2 P53、Bip、HSPA1的转录水平变化
运动组与安静组相比,自主跑轮未明显改变P53的转录水平(图1A),但Bip的mRNA含量明显升高(P<0.05,图1B),且热休克蛋白70(HSPA1)的转录水平显著下降(P<0.05,图1C)。
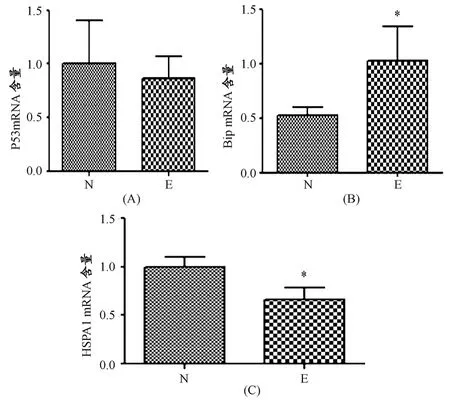
图1 本研究股四头肌P53、Bip和HSPA1的转录水平变化示意图Figure 1.A,B and C Representing the Transcription Level of P53,Bip and HSPA1Respectively
2.3 PIM组件的转录水平变化
自主跑轮运动增加了PIM组件Tom20(P<0.05,图2A)、Tom 40(P<0.01,图 2B)和 VDAC3(P<0.05,图2D)的转录水平,但Tim22的mRNA含量未发生明显变化(图2C)。
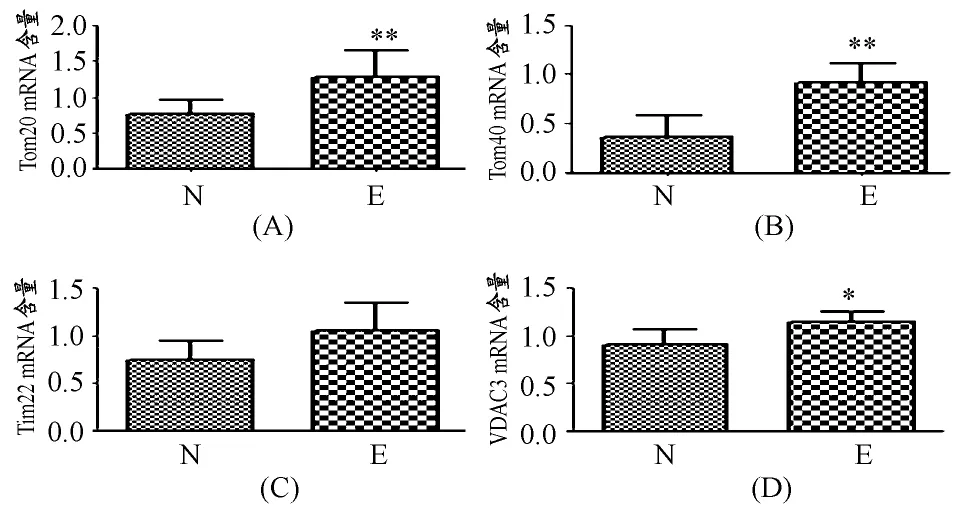
图2 本研究Tom20、Tom40、Tim22、VDAC3的转录水平变化示意图Figure 2.A,B,C and D Representing the Transcription Level of Tom20,Tom40,Tim22and VDAC3Respectively
2.4 P53、Tom 40、Tim22、HSPA1的蛋白表达量变化
自主跑轮运动未使P53(图3A)和Tom40(图3B)的蛋白表达量发生明显变化,但增加了HSPA1的蛋白含量(P<0.05,图3D,图4),显著降低了Tim22的蛋白表达量(P<0.01,图3C,图4)。

图3 本研究P53、Tom40、Tim22和HSPA1蛋白表达量变化示意图Figure 3.A,B,C and D Representing the Protein Expression of P53,Tom40,Tim22and HSPA1Respectively

图4 本研究P53、Tom40、Tim22、HSPA1以及相应内参的表达示意图Figure 4.The Protein Expression of P53,Tom40,Tim22,HSPA1and Corresponding Endogenous References
3 讨论
本团队前期已进行了相关线粒体PIM组件和功能方面的研究,其中,重点研究了线粒体生物发生关键分子PGC-1α和Bax/Bak与线粒体PIM经典途径组件蛋白表达以及PIM功能的关系。前期的研究已发现,PGC-1α基因缺失对骨骼肌线粒体PIM经典路线组件表达的影响呈多样化,对PIM功能无显著影响,但会造成细胞质环境改变影响PIM功能。该研究还发现运动可改善因Bax/Bak敲除所引起的PIM功能受损,且可能与细胞质内质网应激(ER STRESS)、未折叠蛋白反应(unfolded protein response,UPR)等生理过程密切相关[1]。基于前期的研究,本研究拟进一步探讨骨骼肌PIM经典途径组件在运动方面的应用性,同时,还关注运动对线粒体PIM非经典路线组件的影响,从而为更全面地了解线粒体PIM与运动的关系打下基础。
3.1 自主跑轮运动对PIM经典途径组件的影响
线粒体内绝大部分的线粒体蛋白是由核基因编码,这一类蛋白在细胞质核糖体合成,随后经PIM进入线粒体,因此,PIM对于维持线粒体功能和真核细胞的生存能力至关重要[6]。在PIM的途径中,最经典的路线是导肽途径[13],这条途径被超过半数的线粒体蛋白使用[21]。携带了导肽的蛋白质前体物质经线粒体外膜转位酶(TOM)复 合 体 (包 括 Tom20、Tom40、Tom70 等 )识别[16]。经TOM渠道,可分裂的前体蛋白被转移至线粒体内膜转位酶23(Tim23)。随后,ATP依赖的热休克蛋白70(HSP70,HSPA1)等可促使蛋白前体转位进入线粒体基质[12]。因此,本研究重点关注此经典PIM路线。研究者发现,小鼠经过8周的自主跑轮运动后,骨骼肌PIM组件Tom20、Tom40的转录水平显著升高,尤其是Tom40的mRNA含量升高极显著,尽管如此,Tom40的蛋白表达变化却无统计学差异,可能是因为自主跑轮运动强度较低,激活了Tom40的转录水平,但蛋白表达未能出现明显变化。说明在此经典途径中,相较于Tom20,Tom40可能对跑轮运动有更高的敏感性。另外,Joseph等人[9]的研究发现,骨骼肌经过长期的收缩活动后,进入线粒体的Tom40增加,且TOM复合体呈收缩时间依赖性募集。这与本研究结果类似,据此可推测若加大运动强度或延长自主跑轮周期,Tom40的蛋白表达可能会发生变化。
上述提及PIM经典途径中有一个重要的伴侣分子HSPA1,HSPA1是依赖ATP的热休克蛋白,其作为伴侣蛋白家族的重要成员之一,有一项重要功能就是介导其他蛋白正确装配,加速正确的肽链折叠,维持某些肽链的伸展状态以利于其跨膜转位,HSPA1在线粒体等不同区域内发挥该作用[3,7]。此外,HSPA1还是细胞质伴侣复合体的重要组成部分,该复合体可在蛋白前体物质经过TOM复合体时促使蛋白前体进入线粒体基质。在此过程中,此复合体主要与重要受体Tom70对接传递目标蛋白至线粒体[4]。本研究发现,虽然运动组的HSPA1的转录水平出现下降,但8周的自主跑轮运动可显著增加HSPA1的蛋白表达量,其表达的增高也间接说明自主跑轮运动可一定程度促进线粒体蛋白输入。有研究报道,长时间亚极量运动后,大鼠下肢比目鱼肌、腓肠肌和左心室肌 HPS70表达增高[8,18]。Liu等人[11]的研究也发现,在4周的递增负荷运动中,HSP70类蛋白表达增加。本研究采用的是8周自主跑轮运动,但HSPA1的表达仍出现显著增加,说明尽管8周的自主跑轮运动强度较小,但HSPA1对运动的作用有较敏感的反应。此外,不仅HSPA1是PIM经典途径的重要伴侣分子,电压依赖性离子通道(VDAC)亦是重要的PIM组件,VDAC有3种同工型,分别是 VDAC1、VDAC2、VDAC3。有研究指出,VDAC与Tom40结构类似,两者均为β桶状跨膜蛋白,且VDAC与Tom20和Tom22相互作用有利于蛋白前体物质经TOM复合体输入[10]。本团队前期的研究结果显示,8周的自主跑轮运动未能使VDAC复合体表达发生明显变化[1],而本研究发现,运动组VDAC3的转录水平明显升高,推测运动改变PIM组件可能体现在不同的VDAC同工型上。
综上所述,8周的自主跑轮运动增加了PIM组件Tom40和VDAC3的转录水平,同时,HSPA1的蛋白表达也显著增加,表明运动对于线粒体经典输入途径有一定促进作用,且跑轮运动对PIM经典途径组件中的HSPA1的影响较显著。
3.2 自主跑轮运动对Tim22的影响
除了经典的PIM路线外,一些疏水性代谢物载体的前体物质需经过非经典途径输入线粒体。这类蛋白的前体物质在细胞质中与伴侣分子结合,转移至Tom70受体,经TOM复合体转位后,随后在线粒体内膜Tim22的作用下嵌入线粒体膜中[6]。遗憾的是,目前关于运动结合PIM的研究多集中于PIM经典途径,关于运动对非经典PIM途径的影响的研究甚少。故本研究关注了PIM组件Tim22,发现运动组Tim22的蛋白表达显著下降,说明运动并未促进该PIM组件的表达,反而使其表达降低。由此推测,低强度运动对不同的PIM路线的作用效果可能不同,抑或是自主跑轮运动作为一种应激状态,在该应激状态下,线粒体对蛋白输入的种类有选择性。因此,不同运动方式或强度对不同PIM途径的影响值得进一步研究。
3.3 自主跑轮运动对P53和Bip的影响
P53是重要的线粒体生物发生分子,同时,也是运动适应的敏感分子。Saleem等人[17]研究了在一次性急性运动和耐力运动中P53激活线粒体生物发生和细胞自噬/线粒体自噬的必要性,发现P53基因敲除的小鼠运动能力显著受损,且线粒体生物发生下游分子表达降低,说明P53对于运动适应和线粒体生物发生有重要作用。关于运动过程中P53变化的研究结果却不尽相同,Patki等人[14]以帕金森疾病模型小鼠为研究对象,发现长期运动可减少细胞色素C的释放和增加P53的表达。Qi等人[15]就发现,糖尿病大鼠经过8周的跑台运动后P53的表达水平明显下降。
本研究试图将线粒体生物发生与线粒体蛋白输入相联系,研究结果显示,自主跑轮运动未使P53的转录水平和蛋白表达发生明显变化,可能是因为8周的自主跑轮运动强度较低,不足以引起P53的变化,若增加运动负荷和延长运动时间P53发生何种变化值得研究。
本团队前期的研究关注了线粒体生物发生另一个关键分子——PGC-1α,发现PGC-1α基因敲除的小鼠PIM组件和功能未发生明显变化,但却引起了内质网应激重要分子Bip的变化[1],而Bip是一种胞浆内质网分子伴侣蛋白,又名Grp87,属于HSP70蛋白家族,是一种钙离子结合分子伴侣,主要位于内质网,其对保证蛋白进行正常糖基化和折叠,膜蛋白和分泌蛋白的正确定位,以及阻截异常蛋白具有非常重要的作用[2]。Bip分子表达的增高往往提示内质网非折叠蛋白反应(UPR)增加,细胞质环境发生改变。Zhang等人[22]推测,UPR的发生可能在某种程度上影响胞浆内蛋白生成与折叠质量,而蛋白折叠状态直接影响输入线粒体的效率。并非发生UPR就使得PIM受损,在一定应激下,UPR被适度激活是对细胞的一种保护作用,且已有研究指出,6周的自主跑轮运动即可使Bip表达水平显著增高,同时改善PIM功能[22]。
本研究检测了Bip分子的变化,发现自主跑轮运动可显著增加Bip的转录水平,说明8周的自主跑轮运动适度激活了UPR,改变了细胞质环境,这很可能也是运动促进PIM经典路线组件以及伴侣分子表达的机制之一。
尽管关于运动与PIM的研究较少,但在PIM经典路线组件方面,Joseph等人[9]的研究就发现,骨骼肌经过长期的收缩活动后,Tom40进入线粒体增加,且TOM复合体呈收缩时间依赖性募集,这些变化与PIM关键蛋白(包括 Tim17、Tim23、Tom20和 HSP90)的表达增加一致。Gordon等人[5]和 Takahashi等人[19]的研究也发现,长期肌肉收缩活动引起的骨骼肌适应性改变,部分原因是由于其同时改变了参与PIM的多种蛋白的表达,这些改变体现在细胞质伴侣蛋白Hsp70以及线粒体内伴侣蛋白mtHsp60、mtHsp70等蛋白表达水平的增加,该研究还发现,大鼠经过7天的电极刺激肌肉收缩后,Tom20的蛋白水平即显著升高。
本研究则发现,自主跑轮运动可明显增加Tom20、Tom40以及VDAC3的转录水平,同时,HSPA1的蛋白表达明显增加,故与上述研究结果基本一致。线粒体是细胞内的能量代谢场所,运动可促进线粒体生物发生和ATP生成[17],虽然,本团队前期研究发现线粒体生物发生相关分子PGC-1α敲除对PIM组件的影响有限,但也推测线粒体生物发生可能通过改变细胞质环境或其他途径影响PIM[1],而本研究亦发现,细胞质内质网应激分子Bip转录水平升高,故推测运动促进线粒体生物发生和ATP生成,改变细胞质环境,促进PIM经典路线组件表达,而PIM组件的变化也可能是运动引起骨骼肌收缩从而导致的肌肉适应性改变的机制之一。这为后续研究PIM组件与运动适应性的关系提供了一定实验依据。目前,关于运动对PIM非经典路线的影响的研究较少,而本研究发现,自主跑轮运动显著降低了PIM非经典路线组件Tim22的蛋白表达,故不同的PIM路线对运动的不同敏感性值得研究,且在运动作用下,线粒体对输入蛋白的种类可能存在选择性。
4 结论
自主跑轮运动使PIM经典途径组件Tom40和伴侣分子VDAC3的转录水平升高,并且显著增加了另一伴侣分子HSPA1的蛋白表达水平。提示,运动有助于促进PIM经典途径。运动亦使内质网应激关键分子Bip的转录水平增高,提示,自主跑轮运动可适度激活UPR,改变细胞质环境,从而促进PIM经典途径组件的表达,改善PIM经典路线蛋白输入。此外,自主跑轮运动显著降低了非经典PIM途径组件Tim22的蛋白表达量,说明自主跑轮运动对不同的PIM途径可能有不同的影响,抑或是在运动情况下,线粒体对输入蛋白的种类有一定选择性。
[1]张媛.应激对骨骼肌线粒体PIM组件蛋白表达及功能调节的作用机制研究[D].上海:华东师范大学博士学位论文,2013.
[2]COE H,MICHALAK M.Calcium binding chaperones of the endoplasmic reticulum[J].Gen Physiol Biophys,2009,28:F96-F103.
[3]ELLIS R J.The molecular chaperone concept[J].Semin Cell Biol,1990,1(1):1-9.
[4]FAOU P,HOOGENRAAD N J.Tom34:a cytosolic cochaperone of the Hsp90/Hsp70protein complex involved in mitochondrial protein import[J].Biochim Biophys Acta,2012,1823(2):348-357.
[5]GORDON J W,RUNGI A A,INAGAKI H,et al.Effects of contractile activity on mitochondrial transcription factor A expression in skeletal muscle[J].J Appl Physiol,2001,90(1):389-396.
[6]HARBAUER A B,ZAHEDI R P,SICKMANN A,et al.The protein import machinery of mitochondria-a regulatory hub in metabolism,stress,and disease[J].Cell Metab,2014,19(3):357-372.
[7]HARDWICK K G,PELHAM H R.ERS1aseven transmembrane domain protein from Saccharomyces cerevisiae[J].Nucleic Acids Res,1990,18(8):2177.
[8]IWAKI K,CHI S H,DILLMANN W H,et al.Induction of HSP70in cultured rat neonatal cardiomyocytes by hypoxia and metabolic stress[J].Circulation,1993,87(6):2023-2032.
[9]JOSEPH A M,HOOD D A.Plasticity of TOM complex assembly in skeletal muscle mitochondria in response to chronic contractile activity[J].Mitochondrion,2012,12(2):305-312.
[10]KRIMMER T,RAPAPORT D,RYAN M T,et al.Biogenesis of porin of the outer mitochondrial membrane involves an import pathway via receptors and the general import pore of the TOM complex[J].J Cell Biol,2001,152(2):289-300.
[11]LIU Y,MAYR S,OPITZ-GRESS A,et al.Human skeletal muscle HSP70response to training in highly trained rowers[J].J Appl Physiol,1999,86(1):101-104.
[12]MAPA K,SIKOR M,KUDRYAVTSEV V,et al.The conformational dynamics of the mitochondrial Hsp70chaperone[J].Mol Cell,2010,38(1):89-100.
[13]NARGUND A M,PELLEGRINO M W,FIORESE C J,et al.Mitochondrial import efficiency of ATFS-1regulates mitochondrial UPR activation[J].Sci,2012,337(6094):587-590.
[14]PATKI G,LAU Y S.Impact of exercise on mitochondrial transcription factor expression and damage in the striatum of a chronic mouse model of Parkinson's disease[J].Neur Lett,2011,505(3):268-272.
[15]QI Z,HE J,ZHANG Y,et al.Exercise training attenuates oxidative stress and decreases p53protein content in skeletal mus-cle of type 2diabetic Goto-Kakizaki rats[J].Free Radic Biol Med,2011,50(7):794-800.
[16]SAITOH T,IGURA M,OBITA T,et al.Tom20recognizes mitochondrial presequences through dynamic equilibrium among multiple bound states[J].EMBO J,2007,26(22):4777-4787.
[17]SALEEM A,CARTER H N,HOOD D A.p53is necessary for the adaptive changes in cellular milieu subsequent to an acute bout of endurance exercise[J].Am J Physiol Cell Physiol,2014,306(3):C241-C249.
[18]SKIDMORE R,GUTIERREZ J A,GUERRIERO V J,et al.HSP70induction during exercise and heat stress in rats:role of internal temperature[J].Am J Physiol,1995,268(1Pt2):R92-R97.
[19]TAKAHASHI M,CHESLEY A,FREYSSENET D,et al.Contractile activity-induced adaptations in the mitochondrial protein import system[J].Am J Physiol,1998,274(5Pt1):C1380-C1387.
[20]TAKAHASHI M,HOOD D A.Protein import into subsarcolemmal and intermyofibrillar skeletal muscle mitochondria.Differential import regulation in distinct subcellular regions[J].J Biol Chem,1996,271(44):27285-27291.
[21]VOGTLE F N,WORELKAMP S,ZAHEDI R P,et al.Global analysis of the mitochondrial N-proteome identifies a processing peptidase critical for protein stability[J].Cell,2009,139(2):428-439.
[22]ZHANG Y,IQBAL S,O'LEARY M F,et al.Altered mitochondrial morphology and defective protein import reveal novel roles for Bax and/or Bak in skeletal muscle[J].Am J Physiol Cell Physiol,2013,305(5):C502-C511.
猜你喜欢
临床肺科杂志(2022年3期)2022-11-26
分子催化(2022年1期)2022-11-02
中华实用诊断与治疗杂志(2022年1期)2022-08-31
能源工程(2022年2期)2022-05-23
中国卒中杂志(2021年7期)2021-11-29
商用汽车(2021年4期)2021-10-13
电力设备管理(2020年4期)2020-12-05
无线互联科技(2020年10期)2020-08-14
中国体育科技(2018年6期)2018-12-13
新民周刊(2018年8期)2018-03-02
