
植物乳杆菌谷氨酸脱羧酶基因的克隆及原核表达
2015-10-09 13:38时粲等
湖北农业科学 2015年17期
时粲等
摘要:根据GenBank中植物乳杆菌(Lactobacillus plantarum)基因组序列设计引物,利用PCR技术扩增了植物乳杆菌QL-14的谷氨酸脱羧酶编码基因gadB,克隆至表达载体pGEX-4T-3,转化大肠杆菌(Escherichia coli)后进行原核表达,对表达条件进行了优化并对表达蛋白质进了纯化。结果表明,扩增目的gadB基因的开放阅读框(ORF)全长1 407 bp,编码469个氨基酸,氨基酸序列与植物乳杆菌WCFS1同源性为99.6%。通过优化诱导表达条件,得到纯化蛋白质GAD大小为53.6 ku,等电点为5.58,比活力为9.9 U/mg。
关键词:植物乳杆菌(Lactobacillus plantarum);γ-氨基丁酸;谷氨酸脱羧酶;原核表达;GST标签蛋白质
中图分类号:Q78 文献标识码:A 文章编号:0439-8114(2015)17-4328-04
DOI:10.14088/j.cnki.issn0439-8114.2015.17.058
γ-氨基丁酸(GABA)作为一种非蛋白质氨基酸,是目前研究较为深入的哺乳动物脑组织的一种重要的抑制性神经递质[1],具有一系列的生理功能,例如可降血压[2]、治疗癫痫[3]、抗疲劳[4]、提高免疫力[5]等。谷氨酸脱羧酶(GAD)是生物体内催化L-谷氨酸经α-脱羧为GABA的限速酶[6]。
目前,国内外主要采用化学合成法、植物富集法和微生物法制备GABA。化学合成法反应条件剧烈,污染严重,不适合食品添加[7];植物富集法提取的GABA含量较低,提取较为困难[8];微生物发酵法条件温和、安全、成本较低,但后处理过程复杂且生产周期长[9]。通过基因工程技术克隆高活性的GAD进行体外催化是获得大量GABA的潜在技术手段[10,11]。本研究通过克隆植物乳杆菌(Lactobacillus plantarum)QL-14中gadB基因并在大肠杆菌(Escherichia coli)中进行原核表达,借助大肠杆菌生长迅速、易于培养及融合表达质粒pGEX-4T-3可人工调控表达的优点进行高效表达,同时对其进行纯化以期获得有较高活性的GAD,为今后将GAD应用于GABA生产奠定基础。
1 材料与方法
1.1 材料
1.1.1 菌株及质粒 大肠杆菌BL21、植物乳杆菌QL-14和质粒pGEX-4T-3由广西科技大学生物与化学工程学院提供。
1.1.2 试剂 PrimeSTAR HS DNA Polymerase、DNA Marker、T4 DNA连接酶、限制性内切酶BamHⅠ和EcoRⅠ以及TA克隆试剂盒购自TaKaRa公司,质粒小量提取试剂盒、胶回收试剂盒购自北京天根生化科技有限公司,低分子质量蛋白质Marker、氨苄青霉素(AMP)、IPTG购自北京索莱宝科技有限公司。
1.2 方法
1.2.1 引物设计与合成 根据GenBank中发表的植物乳杆菌WCFS1全基因序列(登录号:NC004567),针对其gadB基因用Primer Premier5.0设计1对特异性引物lpgadbs/lpgadba,同时,根据融合表达载体pGEX-4T-3上多克隆位点的特征,在两条引物5′端分别插入限制性内切酶位点(下划线部分),由上海英俊生物工程有限公司合成。引物序列lpgadbs:5′-GGATCCATGGCAATGTTATACGGTA-3′;lpgadba:5′-GAATTCTCAGTGTGTGAATCCGT-3′。
1.2.2 gadB基因的克隆 以植物乳杆菌QL-14菌液为模版,按如下反应物体系进行PCR反应:0.5 μL PrimeSTAR HS DNA Polymerase(2.5 U/μL),10 μL 5×PrimeSTAR Buffer,4 μL dNTP Mixture(各2.5 mmol/L),2 μL模板(菌液),引物lpgadbs/lpgadba(10 mmol/L)各1 μL,添加ddH2O至50 μL。反应条件:94 ℃预变性5 min;94 ℃变性50 s,59 ℃退火90 s,72 ℃延伸2 min,35个循环;72 ℃延伸10 min。1.0%琼脂糖凝胶电泳后进行胶回收。加20 μL A体系 [13.5 μL 胶回收产物,0.5 μL TaKaRa TaqTM(5 U/μL), 2 μL 10×PCR Buffer(Mg2+ free),3 μL dNTP Mixture(各2.5 mmol/L),1 μL MgCl2(25 mmol/L)],72 ℃反应30 min后与pMD18-T载体在16 ℃连接过夜,连接产物转化至感受态细胞,在含100 μg/mL AMP的LB平板上进行抗性筛选。挑选单克隆经PCR、双酶切验证后送至上海英俊生物工程有限公司测序。
1.2.3 融合表达质粒的构建及鉴定 将质粒pMD18-T-gadB和pGEX-4t-3融合质粒分别经限制性内切酶BamHⅠ和EcoRⅠ双酶切,以10∶1的体积比混合后加入T4 DNA连接酶16 ℃连接过夜,将连接产物转化至感受态细胞,在含100 μL/mL AMP的LB平板上进行抗性筛选。挑选单克隆经PCR、双酶切验证后进行测序。
1.2.4 重组蛋白质诱导表达 取新鲜培养的重组菌菌液1.0 mL(含100 μg/mL AMP)接种于100 mL LB液体培养基(含100 μg/mL AMP)上,37 ℃、120 r/min摇床培养至OD600 nm=0.4~0.6时加入异丙基-β-D-硫代吡喃半乳糖苷(IPTG)进行诱导表达。诱导完成后取1.0 mL菌液浓缩至150 μL,于12 000 r/min 离心5 min,去上清液后加入2×样品缓冲液煮沸15 min,然后12 000 r/min离心15 min,取上清液进行10%的SDS-PAGE凝胶电泳。SDS-PAGE凝胶配制及电泳条件参考《蛋白质技术手册》[12]。endprint
1.2.5 融合蛋白质可溶性分析 将诱导后的菌体悬浮于PBS,利用超声细胞破碎仪破碎菌体(35 W,工作2 s间歇1 s,10 min)。将破碎后的菌液6 000 r/min离心10 min,分别取上清液和沉淀进行电泳。
1.2.6 GAD的纯化 将诱导表达的菌液400 mL于12 000 r/min、4 ℃离心15 min,收集菌体,用20 mL PBS缓冲液(pH 7.4)悬浮菌体,超声波破碎细胞,然后经0.45 μm滤膜过滤后上柱,用GST(谷胱甘肽)琼脂糖凝胶柱对重组蛋白质进行纯化,得到纯化后的GST-GAD,通过凝血酶Thrombin将标签蛋白质GST切除掉,得到目的蛋白质GAD。纯化后的蛋白质含量采用Bradford[13]法测定,以BSA(牛血清蛋白质)为标准蛋白质。
1.2.7 GAD活力的测定 采用比色法。配制一定量的底物溶液Macllvaine[14]缓冲体系(0.2 mol/L,pH 4.8,含0.02 mol/L底物L-谷氨酸钠),取400 μL底物溶液和200 μL酶液在37 ℃反应30 min后煮沸终止反应;再加入1.0 mL 6%(V/V)苯酚和1.0 mL次氯酸钠溶液,充分振荡后,沸水浴中反应10 min,迅速在冰浴中冷却显色20 min,待出现蓝绿色后加入60%乙醇溶液2.0 mL,最后在643.5 nm处测定吸光度。酶反应体系中GABA的量以标准曲线确定,一个酶活力单位(U)定义为在测定条件下每分钟产生1 μmol GABA所需的酶量。
2 结果与分析
2.1 gadB基因的克隆及表达质粒的构建
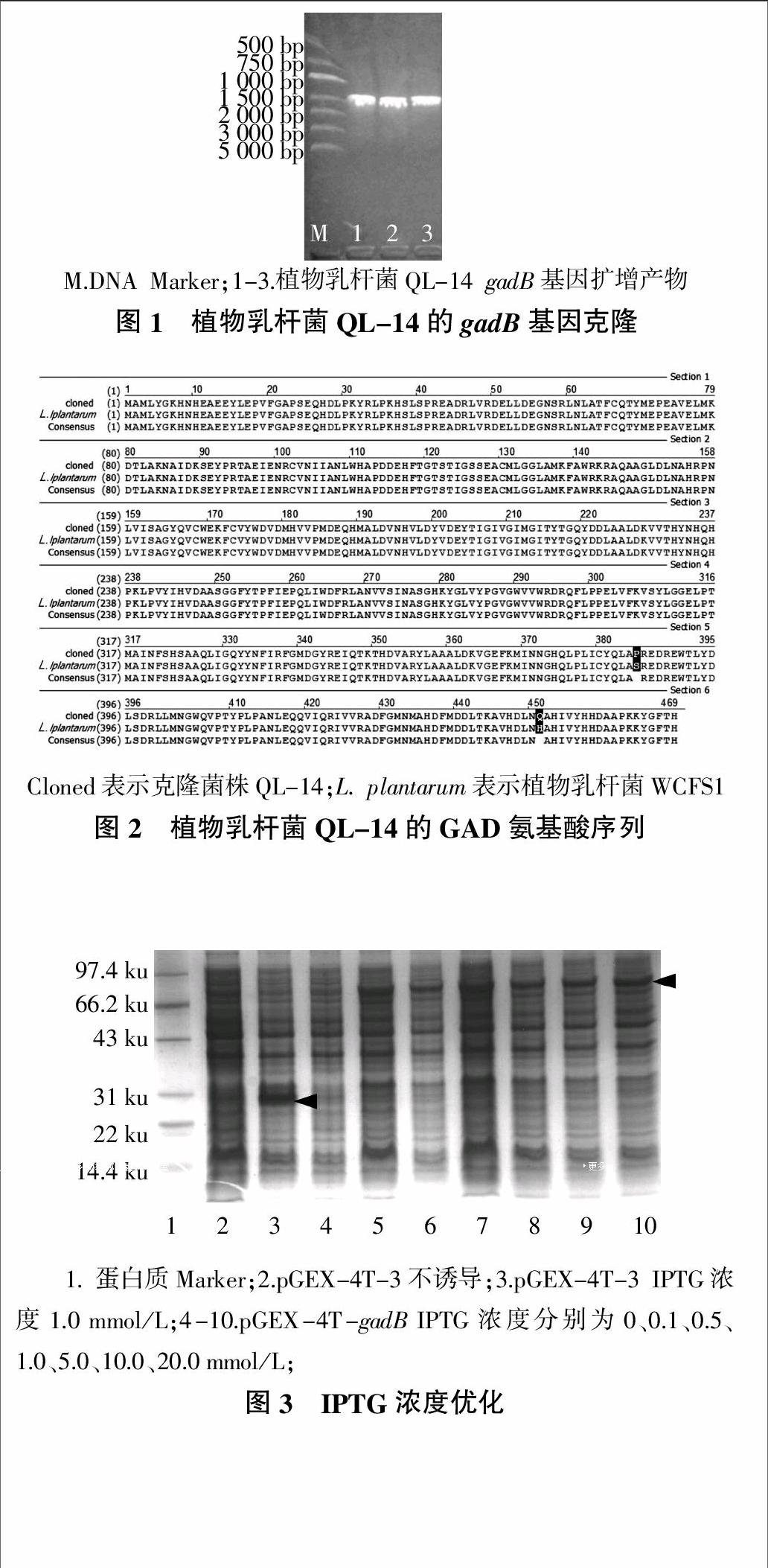
gadB基因PCR产物经琼脂糖凝胶电泳分析后可见一段约1 400 bp的特异性条带(图1)。测序结果显示,该片段长1 410 bp,含全长为1 407 bp的开放阅读框(ORF)编码序列。克隆的gadB基因序列已经提交到GenBank数据库,登录号为KJ542619,氨基酸序列登录号为AHY84727。
pMD-18-gadB与pGEX-4T-3分别经双酶切后连接,转化大肠杆菌BL21感受态细胞,在含100 μg/mL AMP的LB平板上进行抗性筛选。挑选单克隆培养后提取重组质粒,以质粒为模板进行PCR及双酶切验证,测序结果表明目的基因成功地克隆到融合表达载体pGEX-4T-3中。对测序后的氨基酸序列进行多重序列比对,结果如图2所示,发现其序列与植物乳杆菌WCFS1的氨基酸序列仅存在两个氨基酸位点的差异,同源性较高,为99.6%。
2.2 gadB基因重组蛋白质的诱导表达结果
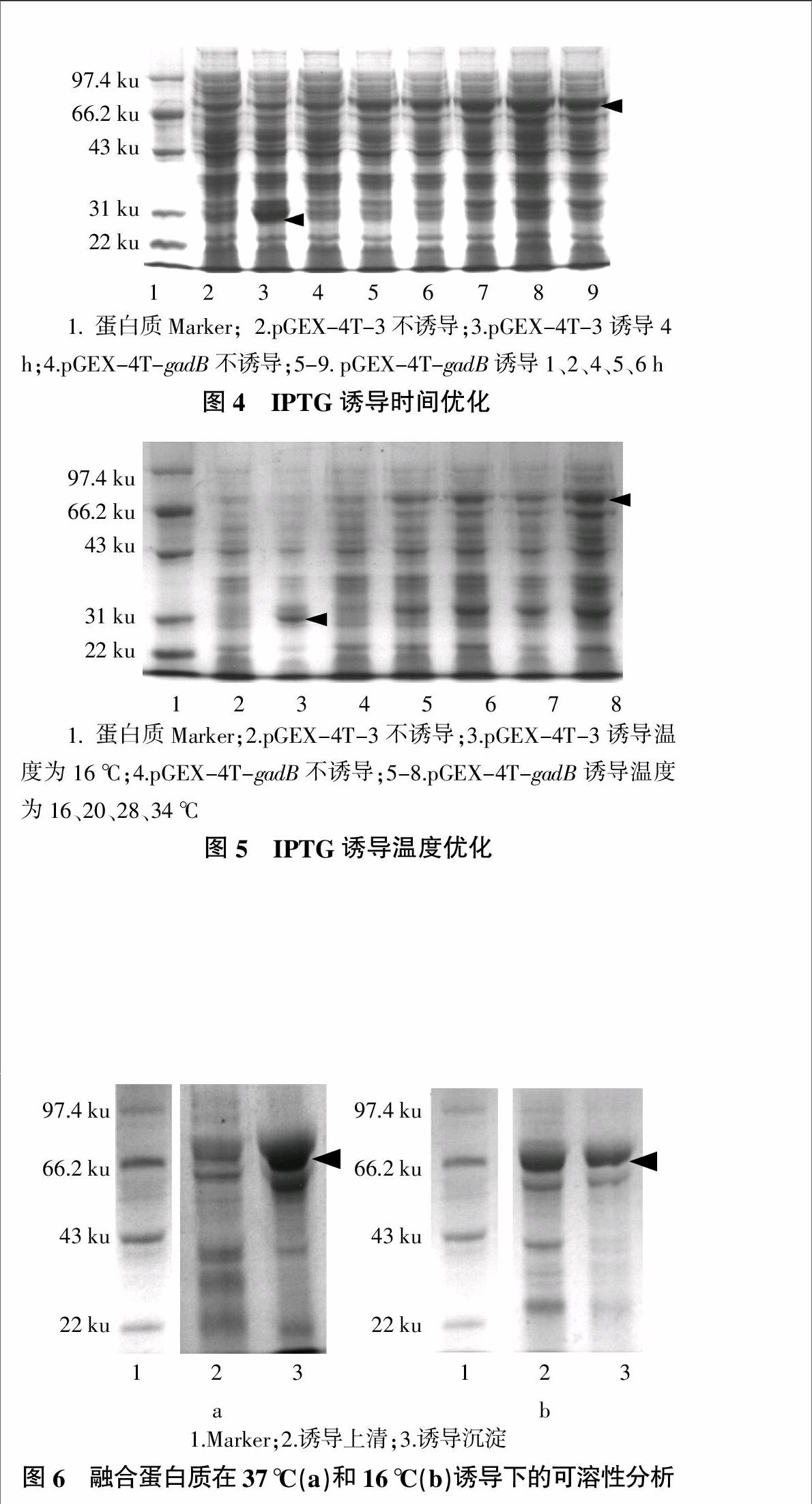
根据“1.2.4”所述,分别从IPTG浓度、诱导时间和诱导温度对重组蛋白质的诱导表达条件进行了优化。从SDS-PAGE电泳结果可以看出,在大约79 ku处有明显条带,说明融合蛋白质已经得到表达,同时以空载体进行诱导表达作为对照。经过上述优化,确定以IPTG浓度1.0 mmol/L(图3)、诱导时间5 h(图4)、诱导温度34 ℃(图5)的条件对重组质粒进行诱导表达并对其表达蛋白质进行可溶性分析。
2.3 融合蛋白质可溶性分析结果
由于使用大肠杆菌pGEX-4t-3高效表达系统,在一定的诱导条件下重组蛋白质产生后来不及进行构象化而迅速积累,常形成不可溶的包涵体,这可能影响目的蛋白质功能[15]。为此,本研究对融合蛋白质的可溶性进行了检测,结果如图6所示。从图6可以看出,37 ℃诱导过程中融合蛋白质大多以包涵体的形式存在于沉淀中(图6a),16 ℃诱导融合蛋白质大多以可溶性的形式存在于上清液中(图6b)。为了得到大量的可溶性蛋白质,本研究目的蛋白质的纯化采用16 ℃过夜诱导表达。
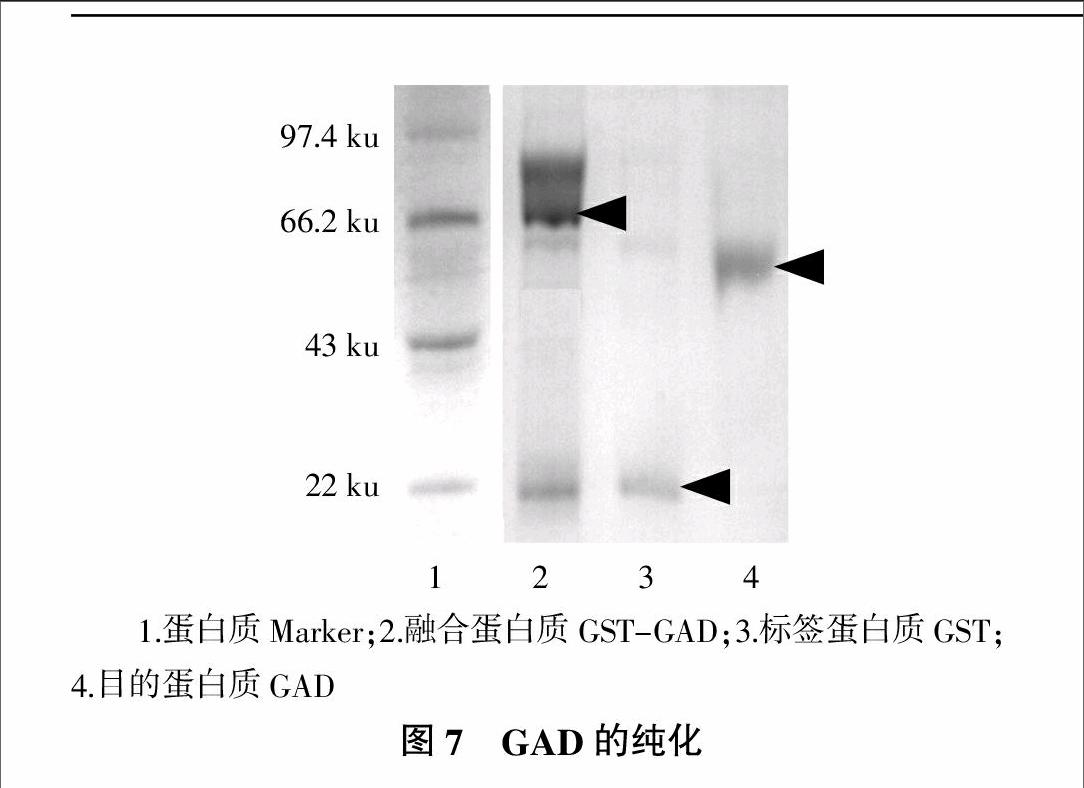
2.4 GAD的纯化结果
重组蛋白质通过GST琼脂糖凝胶柱纯化,通过凝血酶切除标签蛋白质,并经过SDS-PAGE电泳分析,分析结果如图7所示。由图7可知,纯化的融合蛋白质GST-GAD大小在79 ku左右,标签蛋白质GST大小约为26 ku,目的蛋白质GAD为53.6 ku,与报道的目标蛋白质大小基本一致[16]。图7中纯化后得到相对单一的条带,基本确定可以是较纯的GAD,经测定,纯化后GAD蛋白质含量为0.626 mg/mL,等电点为5.58。
2.5 GAD活力的测定结果
纯化的GAD与底物L-谷氨酸钠在一定条件下反应得到GABA,在643.5 nm处检测GABA吸光度,后带入GABA标准曲线。将每分钟产生1 μmol GABA所需的酶量定义为一个酶活力单位(U),得到目的蛋白质的酶活力。纯化后的目的蛋白质GAD有较高的活力,总活力为6.2 U/mL,比活力为9.9 U/mg。
3 结论
通过克隆植物乳杆菌QL-14谷氨酸脱羧酶(共469个氨基酸),发现其与GenBank上已发表的植物乳杆菌L. plantarum WCFS1[17](GenBank登录号:YP 0048490915)的GAD氨基酸序列同源性为99.6%,仅在2个位点有差异,说明在不同的植物乳杆菌菌株中的GAD氨基酸序列略有差异。
经过诱导条件优化,发现IPTG的诱导表达条件与蛋白质的表达量息息相关,其中随着诱导时间的增加表达量增加明显,在诱导5 h时表达量达到最大值,诱导温度的升高致使表达量的增加也是很可观的,而IPTG诱导浓度的增加对目的表达量增加量影响并不大。尹淑琴等[18]采用同样的表达体系进行原核表达所得的最优条件为IPTG浓度0.3 mmol/L、诱导温度32 ℃。说明相同的表达体系中不同的基因表达条件不同。重组蛋白质可溶性分析中,发现融合蛋白质GST-GAD在37 ℃高温诱导条件下易形成不可溶的包涵体,不利于纯化,所以选择在16 ℃下过夜诱导。诱导后蛋白质经GST琼脂糖凝胶柱纯化得到有一定纯度的目的蛋白质GAD(大小53.6 ku,等电点5.58),并用凝血酶切除掉标签蛋白质GST,排除了可能对GAD活性有干扰的因素。通过酶活力测定发现纯化蛋白质GAD总活力为6.2 U/mL,比活力为9.9 U/mg,有较高酶活力,具备工业催化生产GABA的潜在条件。endprint
综合上述,本试验成功克隆了植物乳杆菌gadB基因,并构建了重组表达菌株,对重组蛋白质的表达条件进行了优化并且得到了有较高活性的目的蛋白质GAD,为后续对GAD性质的进一步研究奠定了基础。
参考文献:
[1] ERLANDER M J, TOBIN A J. The structural and functional heterogeneity of glutamic acid decarboxylase:A review[J]. Neurochem Reserch,1991,16(3):215-226.
[2] DE LA GARZA R, ZORICK T, HEINZERLING K G, et al. The cardiovascular and subjective effects of methamphetamine combined with gamma-vinyl-gamma-aminobutyric acid (GVG) in non-treatment seeking methamphetamine-dependent volunteers[J]. Pharmacology biochemistry and behavior,2009,94(1):186-193.
[3] PODELL M, HADJICONSTANTINOU M. Cerebrospinal fluid gamma-aminobutyric acid and glutamate values in dogs with epilepsy[J]. American journal of veterinary research,1997,58(5):451-456.
[4] 茅 原, 杉浦友美.近年のGABA生理机能研究-脑机能改善作用,高血压作用を中心に[J].食品开发,2001,36(6):4-6.
[5] PARK K, KIM N, OH C, et al. Effects of lactic acid bacteria cultures with enhanced levels of GABA on immune cell stimulation[J]. The FASEB Journal,2006,20:270-274.
[6] UENO H. Enzymatic and structural aspects on glutamate decarboxylase[J]. Journal of Molecular Catalysis B: Enzymatic, 2000,10(1-3):67-79.
[7] 李良铸,李明哗.最新生化药物制备技术[M].北京:中国医药科技出版社,2001.79-80.
[8] 许建军.γ-氨基丁酸(GABA)——一种新型的功能性食品因子[J].食品工业科技,2003,24(1):109-110.
[9] CHOI S I,LEE J W,PARK S M,et al.Improvement of γ-aminobutyric acid (GABA) production using cell entrapment of Lactobacillus brevis GABA 057[J]. Microbiol Biotechnol,2006, 16(4):562-568.
[10] LIN Q,YANG S Y.Cloning and expression of glutamate decarboxylase gene from Streptococcus thermophilus Y2[J].The Journal of General and Applied Microbiology,2009,55(4): 305-310.
[11] KIM S H,SHIN B H. Cloning and expression of a full-length glutamate decarboxylase gene from Lactobacillus brevis BH2[J].Biotechnology and Bioprocess Engineering,2007,12(6):707-712.
[12] 汪家政,范 明.蛋白质技术手册[M].北京:科学出版社,2000. 77-108.
[13] BRADFORD M M. A rapid and sensitive method for the quantization of microgram quantities of protein utilizing the principle of protein-dye binding[J]. Anal Biochem,1976, 72(9):248-254.
[14] 许建军,江 波,许时婴.比色法快速测定乳酸菌谷氨酸脱羧酶活力及其应用[J].微生物学通报,2004,31(2):66-71.
[15] BESSETTE P H,ASLUND F,BECKWITH J,et al. Efficient folding of protein with multiple disulfide bonds in the Escherichia coli cytoplasm[J]. Proc Nat1 Acad Sci USA,1999, 96(24):13703-13708.
[16] NOMURA M,NAKAJIMA I,FUJITA Y.Lactococcus lactis contains only one glutamate decarboxylase gene[J]. Microbiology, 1999,145(6):1375-1380.
[17] SIEZEN R J,FRANCKE C,RENCKENS B,et al.Complete resequencing and reannotation of the Lactobacillus plantarum WCFS1 genome[J].J Bacteriol,2012,194(1):195-196.
[18] 尹淑琴,常 泓,范 艳,等.钙激活酶激活蛋白基因的克隆及其在原核生物中表达及条件的优化[J].激光生物学报,2012(2):156-161.endprint
猜你喜欢
环球时报(2022-09-20)2022-09-20
今日农业(2020年24期)2020-12-15
当代陕西(2020年9期)2020-08-04
食品科学(2018年10期)2018-05-23
兽医导刊(2016年12期)2016-05-17
中国工程咨询(2016年6期)2016-01-31
中国卫生(2015年7期)2015-11-08
西南医科大学学报(2015年1期)2015-08-22
中国当代医药(2015年9期)2015-03-01
现代检验医学杂志(2015年4期)2015-02-06
