
藏东南高寒森林大型真菌物种增补及生态特点1)
2015-09-18 11:12:28王术荣
东北林业大学学报 2015年11期
王术荣
(山西农业大学,太谷县,030801)
汤昕明
(中华全国供销合作总社昆明食用菌研究所)
王琦 李玉
(吉林农业大学食药用菌教育部工程研究中心)
生物多样性处于生态系统稳定性和生产力的中间环节,并且三者之间形成反馈环[1],生物多样性丧失会严重影响生态系统的功能[2-5]。真菌具有重要的生态功能,是纤维物质的初级分解者和循环者,是土壤食物网的重要组分部分,也是从细菌到哺乳动物的许多物种的食物来源。真菌与90%~95%的陆地植物、昆虫[6]和藻类[7]形成共生关系。大型真菌物种多样性是全球生物多样性的重要组成部分。我国大型真菌多样性调查和保护面临较多问题。如:生物物种资源家底不清、调查和编目任务繁重、生物多样性监测和预警体系尚未建立、生物多样性投入不足、基础科研能力较弱、应对生物多样性保护新问题的能力不足等,这些突出问题都极大限制了大型真菌多样性的研究。大型真菌“understudied”是世界菌物学家的共识[8-9]。系统研究大型真菌多样性、物种组成和生态特点,对研究真菌在生态系统中的作用以及整个生态系统的发展具有深远的意义。
由于西藏高寒森林地区海拔高、交通不便,真菌资源更少人问津。臧穆[10]于1980年发现了西藏担子菌类的多个新种。1990年前后,中国科学院青藏高原综合考察队对包括南迦巴瓦峰在内的西藏地区做过较为详细的调查[11-13],卯晓岚[14]对西藏大型真菌进行调查后出版了《西藏大型经济真菌》。图力古尔[15]对西藏真菌进行了增补,邓利君等[16-17]对色季拉山食用菌和丝膜菌的种类进行了调查,但相对于西藏的地域和大型真菌资源来说,这些工作远远不够。本研究拟通过对西藏东南高寒森林地区典型植被类型物种资源调查和生态特点分析,旨在进一步探明此地区的大型真菌的种类和资源状况,加强对其生态特点的认识。
1 材料与方法
1.1 材料
供试大型真菌子实体于2012年6—9月和2013年6—8月在西藏东南高寒森林地采集,凭证标本存放于吉林农业大学菌物标本馆(HMJAU)。
1.2 取样、记录和鉴定方法
随机选取西藏色季拉山坡和波密县高寒森林地多个采样点,以10 d为一个周期进行标本采集和物种调查,所有种类都详细记录其生境和生态特点。每份大型真菌标本的生态照片包括标尺照片、非标尺生态照片,以及正面、侧片、剖面照等。生态特点的记录主要依据其生境、营养方式和基物的选择性。根据传统分类研究方法,对所采集到的1100余份标本进行形态特征观察,并做详尽的野外记录,拍摄野外生态照片。
鉴定方法采用宏观特征、显微特征和化学试剂反应相结合的方法。形态观察依靠放大镜和实体解剖镜,宏观特征依靠新鲜材料和干标本,颜色判断主要依据野外记录和当时的彩色照片,颜色名称根据Kornerup[18]的颜色名称手册确定。显微观察主要依靠光学显微镜(重庆光电H6303),将徒手切片材料分别置于蒸馏水、5%KOH和Melzer试剂中进行。数量特征以3~5个子实体为依据,孢子大小测定40个成熟孢子。部分标本的显微特征用手绘的方法,少量标本的担孢子表面显微特征喷金后,用扫描电镜观察[19]。气象数据来源于色季拉山和波密县气象局的气象资料。
2 结果与分析
2.1 西藏地区物种增补
通过分类鉴定,共确定鉴定到种的大型真菌标本2门4纲 16目 44科 118属 239种[20],子囊菌门3纲5目10科12属17种,担子菌门1纲11目34科106属222种。其中22种为西藏新记录种(见表1)。
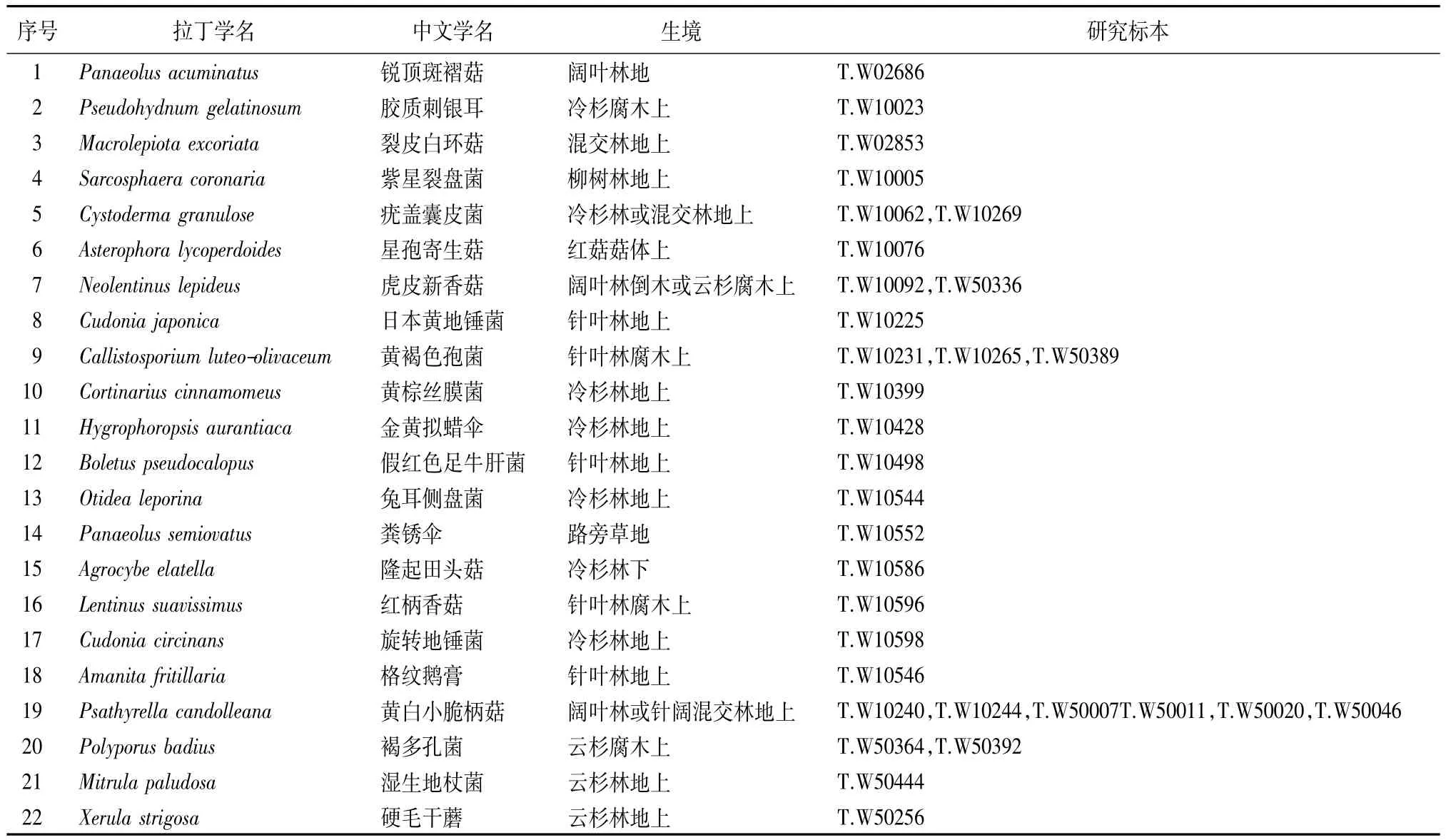
表1 22个西藏新记录种
2.2 科属分布特点
大型真菌种类最多的科是伞菌科(Agaricaceae)和红菇科(Russulaceae),分别有24种和23种,占全部种类的19.67%;其次为牛肝菌科(Boletaceae)和多孔菌科(Polyporaceae),各有16种,占全部种类的13.39%;然后依次是丝膜菌科(Cortinariaceae)、球盖菇科(Strophariaceae)、口蘑科(Tricholomataceae)。这些科共有种数118种,占所采集大型真菌总数的49.37%。
西藏高寒森林地区大型真菌共有118个属,种类(包括种下等级)超过或者等于5个种的属有6个,共有种类51种,占西藏高寒森林地区大型真菌种数的21.34%,而属的数目仅占全部属数的5.08%。6个属中有世界分布属2个,北温带分布属4个。反应出本地区大型真菌是以世界分布属和北温带分布属为主的特点。这些科属中的很多种类都是重要的食药用菌资源,为我们更好地保育、开发利用大型真菌提供了基础资料。
西藏高寒森林地区大型真菌的分布类型研究表明,世界分布种69种和北温带分布种134种是西藏高寒森林地区大型真菌的主要分布型,占总种数的87.12%。在西藏高寒森林地区,欧亚大陆分布种(包括亚洲分布种)、中国-日本共有种、温带-亚热带、热带分布种等占总数的11.60%,其他分布型所含种数比例较小,在一定层次上反映了本地区的大型真菌区系特点。
2.3 营养方式和基物选择
在生态系统中,按照大型真菌对碳源的利用策略,可以分为3种生活类型:起分解作用的腐生真菌、具有共生特点的菌根真菌、极少量的寄生真菌[21]。他们在分解木质有机物、增加植物营养吸收、提高植物抗性和改善土壤结构中发挥着重要作用。根据实际采集情况记录了西藏高寒森林地区大型真菌的营养方式和基物选择情况。除营大型真菌寄生的星孢寄生菇外,其他种均为起分解作用的腐生真菌和菌根真菌。主要的基物偏好为腐木、活木、枯落物、土壤。腐生真菌的物种数量约占总数的39.38%,菌根真菌的物种数量为60.18%。从基物选择来看,营土壤生活的物种数量为63.00%,营木质物生活的为34.80%,营枯落物生活的物种数量为1.76%,在菇体上生长的为0.44%。
2.4 降水和温度对大型真菌发生的影响
根据采集时间(每10 d为一个计算周期)对西藏林芝地区色季拉山和波密县采集到的大型真菌的发生种类进行统计分析。

图1 波密县和色季拉山大型真菌发生与降水量的关系

图2 波密县和色季拉山大型真菌发生和温度变化的关系
由图1可知,波密县大型真菌物种发生的数量与降水量具有一致性,大型真菌物种发生的数量较降水时间约有10 d的延后期,色季拉山的物种发生基本也具有此特点。
由图2可知,色季拉山和波密县平均温度变化和大型真菌种类发生关系不大。
3 结论和讨论
通过对西藏的物种多样性调查和室内鉴定,共发现了1个新种[22]和22个西藏新记录种。22种是鉴定到种的大型真菌种类的1/10,也就是说采集10种大型真菌就有1种是未报道过的西藏菌类,说明对西藏大型真菌资源调查工作的不足。物种组成特征分析表明伞菌科和红菇科的种类最多,其次是牛肝菌科和多孔菌科。营养方式上,腐生真菌的物种数量占总数的39.38%,而菌根真菌的物种数量占总数的60.18%。基物选择上,主要营土壤生活的物种数量为63.00%,主要营木质物生活的约为34.80%,主要营枯落物生活的物种数量为1.76%,在菇体上生长的为0.44%。营养和生活特点的把握对西藏大型真菌资源的保育和开发利用具有重要意义。
西藏高寒地区新记录种的发现填补了生物演化、物种迁移的重要环节。由于西藏独特的地理环境条件,西藏大型真菌标本子实体的形态与其他地点的标本形态有许多差异。尽管采集到的大多数种类属于全球分布种或者北温带分布种,但在鉴定中,发现该地区的标本具有寒带、温带的标本特点,甚至某些种类具有热带种类的形态特点,这可能与西藏高寒地区多变的气候有关。
葡萄牙东北部栗树林大型真菌多样性研究表明,降水量变化对大型真菌的发生具有显著影响,大型真菌发生前3 d的平均气温对大型真菌的发生具有影响,10 d前或更长时间以前的平均温度对大型真菌的发生没有影响[23]。Baptista等[24]对板栗树和大型真菌多样性的关系研究中,也发现物种多度和丰富度年间波动和气候条件有关(主要是降水)。研究表明大型真菌的发生与降水量具有相关性,并且大型真菌发生较降水有一定的延后期,可能是大型真菌的发生需要一定的湿度积累。温度变化对大型真菌的发生影响不明显。这些发现有利于提高我们对大型真菌发生特点和发生因素的深刻理解和认识,同时,对加强大型真菌多样性保护也具有重要的参考价值。
[1]Worm B,Duffy J E.Biodiversity,productivity and stability in real food webs[J].Trends in Ecology & Evolution,2003,18(12):628-632.
[2]Kinzig A P,Pacala S W,Tilman D.The functional consequences of biodiversity:empirical progress and theoretical extensions[M].Princeton:Princeton Univ Pr,2001.
[3]Loreau M,Naeem S,Inchausti P,et al.Biodiversity and ecosystem functioning:current knowledge and future challenges[J].Science,2001,294:804-808.
[4]Loreau M,Naeem S,Inchausti P.Biodiversity and ecosystem functioning:synthesis and perspectives[M].Oxford:Oxford University Press,2002.
[5]Hooper D,Chapin IF,Ewel J,et al.Effects of biodiversity on e-cosystem functioning:a consensus of current knowledge[J].Ecological Monographs,2005,75(1):3-35.
[6]Kendrick B.The fifth kingdom[M].Sidney:Mycologue Publications,1985.
[7]Hawksworth D.The variety of fungal-algal symbioses,their evolutionary significance,and the nature of lichens[J].Botanical journal of the Linnean Society,1988,96(1):3-20.
[8]Hawksworth D.The fungal dimension of biodiversity:magnitude,significance,and conservation[J].Mycological Research,1991,95(6):641-655.
[9]Mueller G M,Bills G F,Foster M S.Biodiversity of fungi:inventory and monitoring methods[M].Burlington:Academic Press,2004.
[10]臧穆.我国西藏担子菌类数新种[J].微生物学报,1980,20(1):29-34.
[11]王云章,臧穆,马启明.西藏真菌[M].北京:科学出版社,1983.
[12]中国科学院青藏高原综合科学考察队,杨逸畴.南迦巴瓦峰登山综合科学考察[M].北京:科学出版社,1993.
[13]李渤生.南迦巴瓦峰地区生物[M].北京:科学出版社,1995.
[14]卯晓岚.西藏大型经济真菌[M].北京:科学技术出版社,1993.
[15]图力古尔,李玉.西藏真菌增补[J].植物研究,2001,21(2):191-194.
[16]邓利君,徐阿生,刘小娇.西藏色季拉山食用菌资源[J].食用菌学报,2011,18(3):91-99.
[17]邓利君,徐阿生,刘小娇.西藏色季拉山丝膜菌属真菌资源[J].中国食用菌,2011,30(5):6-7.
[18]Kornerup A,Wanscher J.Methuen handbook of colour(3rd)Methuen[M].London:Methuen and Co.Ltd.,1978.
[19]Bozzola J J,Russell L D.Electron microscopy:principles and techniques for biologists[M].Boston:John & Bartlet Publishers,1999.
[20]王术荣.西藏高寒森林地区大型真菌多样性研究[D].长春:东北师范大学,2014.
[21]Schmit J P,Mueller G M.An estimate of the lower limit of global fungal diversity [J].Biodiversity and Conservation,2007,16(1):99-111.
[22]Wang S R,Wang Q,Wang D L,et al.Gastroboletus thibetanus:a new species from China[J].Mycotaxon,2014,129(1):79-83.
[23]Gange A,Gange E,Sparks T,et al.Rapid and recent changes in fungal fruiting patterns[J].Science,2007,316:71-71.
[24]Baptista P,Martins A,Tavares R M,et al.Diversity and fruiting pattern of macrofungi associated with chestnut(Castanea sativa)in the Trás-os-Montes region(Northeast portugal)[J].Fungal E-cology,2010,3(1):9-19.
猜你喜欢
学与玩(2022年7期)2022-10-31 01:59:22
科学(2020年2期)2020-08-24 07:56:56
收藏界(2018年1期)2018-10-10 05:23:08
小学科学(学生版)(2018年8期)2018-08-21 12:05:50
创新作文(小学版)(2018年31期)2018-05-16 03:00:30
小猕猴智力画刊(2017年12期)2017-12-27 17:30:31
摄影之友(影像视觉)(2017年1期)2017-07-18 11:12:16
海峡姐妹(2017年4期)2017-05-04 04:03:51
剑南文学(2016年11期)2016-08-22 03:33:36
小学阅读指南·低年级版(2016年6期)2016-05-14 21:39:20
