
不同小生境下小蓬竹形态多样性研究
2015-09-10 16:09李丽霞刘济明骆畅等
江苏农业科学 2015年8期
李丽霞+刘济明+骆畅等
摘要: 为了解小蓬竹资源的适应性及遗传特点,本研究采用野外与室内相结合的方法对小蓬竹6个小生境的14个形态标记指标进行测定分析。结果显示,小蓬竹各表型性状存在不同程度的分化,14个表型性状变异系数均值变化范围为5 7%~68 3%,变异系数除叶长宽比,一级、二级分枝叶面积外,其他性状变异系数都大于14 1%,变异系数最高的是二级分枝数,达68 3%。6个小生境间的小蓬竹形态变异存在差异,土面小生境大多数形态性状的变异系数最大,其次是石槽和石缝,显示不同的生境条件促进了小蓬竹的形态分化。聚类分析结果显示,聚为一类的生境都具有相似的生态因子(如土壤厚度、维持水分的能力等)。主成分分析结果显示,一级分枝叶数、节数、一级分枝数、二级分枝叶数可作为其形态性状的重要主成分。相关性分析结果显示,株高和节平均值呈显著负相关,一级、二级分枝形态及一级、二级分枝叶片形态相互之间都存在正相关性。表明小蓬竹对生境变化反应灵敏,形态可塑性大,对环境适应能力强,遗传多样性比较丰富,可作为喀斯特地区的适生竹种,本研究结果对石漠化地区的植被选择和小蓬竹保护有积极意义。
关键词: 小蓬竹;小生境;形态多样性;喀斯特地区;相关分析
中图分类号: Q945 79 文献标志码: A
文章编号:1002-1302(2015)08-0191-04
小蓬竹[Drepanostachyum luodianense (Yi et R S Wang) Keng f ]属竹亚科(Bambusoideae) 镰序竹属(Drepanostaehyum Keng f ) [1],主产于贵州省黔南州、黔西南州、安顺市一带,其外形美观,有栽培观赏价值,同时又可作为良好的纸浆原料,具有经济价值。但是,近年来小蓬竹无性系种群严重退化,种群数量急剧减少,已被列为极危物种 [2-4]。
形态多样性是遗传多样性与环境多样性的综合体现,是基因组DNA编码序列与其调控序列多样性的具体表现,众多研究集中在种内群体在其分布区内各种环境条件下的表型变异 [5-8]。竹类植物在不同的土壤养分、水分和光照资源水平下,可以通过形态塑造来适应不同的资源水平 [9]。已有的研究表明,小蓬竹可在贵州喀斯特地貌的多种极端生境下生长,是喀斯特石山地区的适生性竹种,对土壤保水、保肥、固土能力效果显著,可作为喀斯特石漠化地区生态恢复与重建的植被 [10-13]。为明确小蓬竹资源的适应性及遗传特点,本研究通过对不同小生境的小蓬竹进行形态指标的变异测定,以明晰该种质的自然表型变异情况,揭示变异程度、变异格局及与地理环境的关系,为筛选喀斯特石漠化地区生态恢复与重建植被的优良变异种质、建立核心种质库、为制定资源保护及利用策略提供参考依据。
1 材料与方法
1 1 研究地概况
研究地点设在贵州省罗甸县,贵州省南部边缘,境内以山地为主,有岩溶、丘陵、盆地及石炭岩等喀斯特地貌,属典型的亚热带温暖湿润季风气候,冬无严寒、夏无酷暑、雨热同季,年均气温为13 6~19 6 ℃,雨量充沛,年均降水量1 100~1 400 mm。调查地设置在罗甸县董架乡东跃村,位于 106°53′~106°54′E、25°37′N,平均海拔997 m。在研究地段的各样地内小蓬竹均占绝对优势,其盖度65%~75%,位于群落中上层,植株上部基本匍甸生长在灌木之上。在群落中小蓬竹与一些乔木、灌木和少量草本混生在一起。森林植被主要类型为亚热带常绿阔叶林,群落乔木层优势种常由壳斗科(Fagaceae)、樟科(Lauraceae)、山茶科(Theaceae)植物组成。
各类喀斯特小生境选取类型为石台小生境、土面小生境、石沟小生境、石缝小生境、石槽小生境、石洞小生境 [14-15]。
1 2 采样与测定
选取贵州省罗甸县董架乡东跃村典型小蓬竹种群,随机在种群中选取正常生长于土面、石沟、石缝、石槽、石洞和石台小生境的小蓬竹进行测量和采样,每类小生境选取3处,每处取1株。叶片指标带回实验室进行测量,其他指标测定在野外完成。测量其株高、节平均直径、节数、一级分枝数、一级分枝叶数、一级分枝长度、一级分枝直径、一级分枝叶长/宽、一级分枝平均叶面积、二级分枝数、二级分枝直径、二级分枝叶数、二级分枝长度、二级分枝平均叶面积共14项指标 [16]。
1 3 统计与分析
计算各性状的变异系数,以变异系数作为生境间各形态特征变异度的量度 [17],变异系数反映遗传变异占均值的大小,以衡量不同变量的变异幅度。变异系数越大,样本间的差异越大。
利用Excel进行变异系数的计算,利用SPSS 18 0对数据进行相关性、主成分及聚类分析。
2 结果与分析
2 1 小生境内小蓬竹形态变异分析
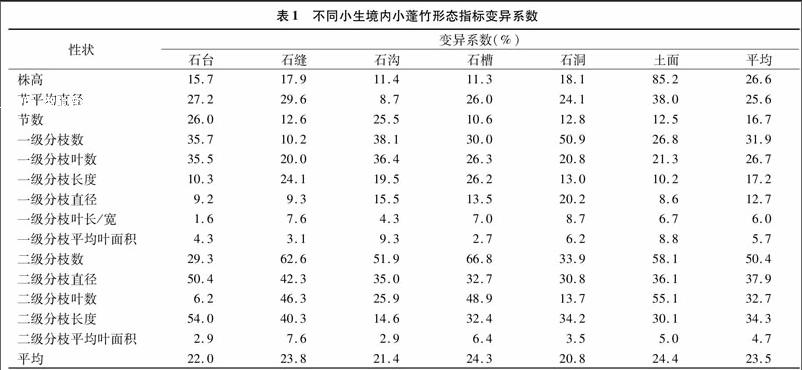
小蓬竹小生境内形态存在着差异,6个小生境的外部形态变异见表1。石台小生境的14个形态指标变异系数除了一级分枝直径、一级分枝叶长/宽、一级与二级枝平均叶面积,其他10个形态指标大于10%。变异系数最大的是二级分枝长度,为54%;其次是二级分枝直径,为50 4%;最小的是一级分枝叶长/宽,为1 6%;平均变异值为22%。
石缝小生境的14个形态指标变异系数除了一级分枝直径、一级分枝叶长/宽、一级分枝平均叶面积、二级分枝平均叶面积,其他10个形态指标变异系数都大于10%。变异系数最大的是二级分枝数,为62 6%;最小的是一级分枝平均叶面积,为3 1%;平均变异值为23 8%。
石沟小生境的14个形态指标变异系数除了节平均直径、一级分枝叶长/宽、一级分枝平均叶面积、二级分枝平均叶面积,其他10个形态指标变异系数都大于10%。最大的是二级分枝数,为51 9%;最小的是二级分枝平均叶面积,为29%;平均变异值为21 4%。
石槽小生境的14个形态指标变异系数除了一级分枝叶长/宽、一级分枝平均叶面积、二级分枝平均叶面积,其他11个形态指标变异系数都大于10%。最大的是二级分枝数,为66 8%;最小的是一级分枝平均叶面积,为2 7%;平均变异值为24 3%。
石洞小生境的14个形态指标变异系数除了一级分枝叶长/宽、一级分枝平均叶面积、二级分枝平均叶面积,其他11个形态指标变异系数都大于10%。最大的一级分枝数,为50 9%;最小的二级枝平均叶面积,为3 5%;平均变异值为20 8%。
土面小生境的14个形态指标变异系数除了一级分枝直径、一级分枝叶长/宽、一级分枝平均叶面积、二级分枝平均叶面积,其他10个形态指标变异系数都大于10%。最大株高,为85 2%;其次二级分枝数,为58 1%;最小二级分枝平均叶面积,为5%;平均变异值为24 4%。
株高变异系数变化范围为11 3%~85 2%,均值22 6,变异系数从大到小依次为土面>石洞>石缝>石台>石沟>石槽;节平均直径变异系数变化范围为8 7%~38 0%,均值 25 6%,变异系数从大到小依次为土面>石缝>石台> 石槽>石洞>石沟;节数变异系数变化范围为10 6%~26 0%,均值16 7%,变异系数从大到小依次为石台>石沟>石洞>石缝>土面>石槽;一级分枝数变异系数变化范围为10 2%~ 50 9%,均值31 9%,变异系数从大到小依次为石洞> 石沟>石台>石槽>土面>石缝;一级分枝叶数变异系数变化范围为20 0%~36 4%,均值26 7%,变异系数从大到小依次为石沟>石台>石槽>土面>石洞>石缝;一级分枝长度变异系数变化范围为10 2%~26 2%,均值17 2%,变异系数从大到小依次为石槽>石缝>石沟>石洞>石台>土面;一级分枝直径变异系数变化范围为9 2%~20 2%,均值127%,变异系数从大到小依次为石洞>石沟>石槽>石缝>石台>土面;一级分枝叶(长/宽)变异系数变化范围为16%~87%,均值6 0%,变异系数从大到小依次为石洞>石缝>石槽>土面>石沟>石台;一级分枝平均叶面积变异系数变化范围为2 7%~9 3%,均值5 7%,变异系数从大到小依次为石沟>土面>石洞>石台>石缝>石槽;二级分枝数变异系数变化范围为29 3%~66 8%,均值50 4%,变异系数从大到小依次为石槽>石缝>土面>石沟>石洞>石台;二级分支直径变异系数变化范围为30 8%~50 4%,均值 379%,变异系数从大到小依次为石台>石缝>土面>石 沟>石槽>石洞;二级分枝叶数变异系数变化范围为6 2%~551%,均值32 7%, 变异系数从大到小依次为土面>石槽>石缝>石沟>石洞>石台;二级分支长度变异系数变化范围为14 6%~54 0%,均值34 3%,变异系数从大到小依次为石台>石缝>石洞>石槽>土面>石沟;二级分支平均叶面积变异系数变化范围为2 9%~7 6%,均值47%,变异系数从大到小依次为石缝>石槽>土面>石洞>石台>石沟;各小生境的所有形态指标的平均变异系数变化范围为 20 8%~24 4%,均值为23 5%,变异系数均值从大到小依次为为土面>石槽>石缝>石台>石沟>石洞。
分析结果表明,土面在很多性状中的变异最大,其次是石槽、石缝。
2 2 小生境间小蓬竹形态性状分析
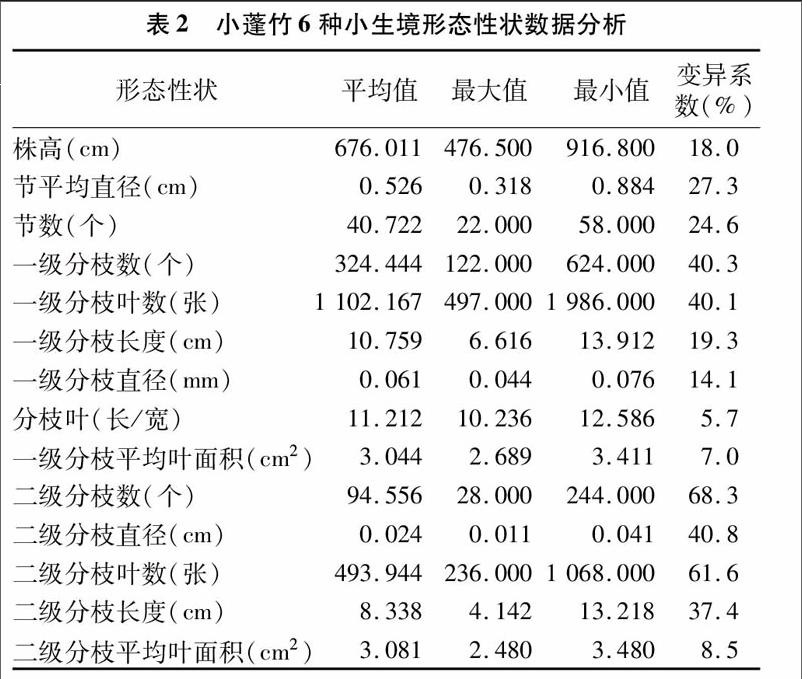
2 2 1 形态性状分析 对6个小生境小蓬竹的14个形态性状的统计分析可知,小蓬竹株高变化范围为476 5~916 8 cm;节平均直径从0 318~0 884 cm;最大节数为58个,最小节数为22个,一级、二级分枝数变化范围分别为122~624、28~244个;一级、二级分枝叶片数变化范围分别为497~1 986、236~1 068张;一级、二级分枝长度变化范围分别为6 616~13 912、4 412~13 218 cm;一级、二级分枝直径变化范围分别为0 044~0 076、0011~0 041 cm;一级、二级分枝叶面积均值变化范围分别为2 689~3 411、2 480~3 480 mm2;分枝叶的长/宽变化范围为10 236~12 586。
从表2看出,小蓬竹各生境间存在形态多样性,变异系数最大的是二级分枝数,达68 3%,变异系数较大的包括二级分枝叶数、一级分枝数、一级分枝叶数、二级分枝直径,变异系数分别为61 6%、40 3%、40 1%、40 8%;变异系数较小的包括株高、一级分枝长度、一级分枝直径,变异系数分别为18 0%、19 3%、14 1%,变异系数最小的是叶长/宽、一级与二级枝叶面积,分别为5 7%、7 0%,8 5%。
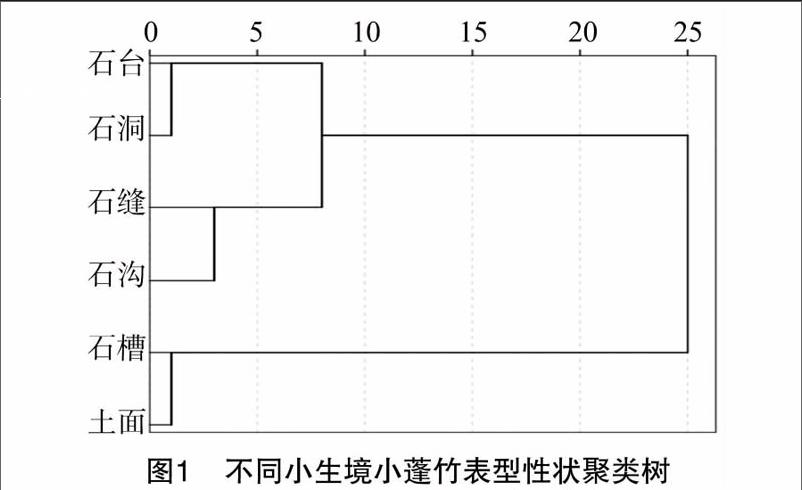
2 2 2 形态性状聚类分析 根据小蓬竹生境形态性状的平均值计算欧式距离,采用UPGMA法进行聚类,结果见图1,小蓬竹生境间形态变异的欧式平均遗传距离变异范围为0~25,从聚类图可以看出,距离3处将6个生境分为3类,石台和石洞聚为一类,石缝和石沟聚为一类,石槽和土面聚为一类。聚为一类的生境条件相似,可见小蓬竹的形态分化受生境条件影响很大,同时也可以说明不同的生境条件促进了小蓬竹的形态分化。
2 2 3 形态性状主成分分析 对小蓬竹的14个形态性状进行主成分分析,结果见表3,前5个主成分的累积贡献率8207%,[CM(22]可以代表原始因子所代表的大部分信息,其中第1主成分占41 139%,一级分枝叶数、节数、一级分枝数、二级分枝叶数对第1主成分的贡献较大,除节数外,其余3个性状都属于一级分枝的形态特征,表明一级分枝形态对小蓬竹形态特征影响较大;第2主成分的贡献率为14 241%,二级分枝叶数、二级分枝数、二级分支长度以及一级分枝长度对第2主成分的贡献较大,此成分只包括一级和二级分枝的性状;第3主成分的贡献率为10 121%,株高和节平均直径对此成分的贡献较大;第4主成分的贡献率为9 328%,一级分枝叶长/宽对第4主成分的贡献较大;第5主成分的贡献率为7240%,一级分枝平均叶面积对此成分的贡献较大,对5个主成分贡献率较大的形态指标表明其受环境影响较大。
2 2 4 形态性状相关性分析 计算小蓬竹形态性状指标的平均值,进行相关性分析与显著性检验,结果见表4,株高与节平均直径存在显著负相关;节平均直径与二级分枝数和二级分枝长度呈正相关;节数与一级分枝数、一级分枝叶数呈极显著正相关,与二级分枝叶数呈显著正相关;一级分枝数与一级分枝叶数、二级分枝数、二级分枝叶数呈极显著正相关,与一级分枝直径、二级分枝直径呈显著正相关;一级分枝叶数与一级分枝直径、二级分枝数以及二级分枝叶数呈极显著正相关;一级分枝长度与二级分枝直径呈显著正相关;一级分枝直径与二级分枝叶数呈极显著正相关,与二级分枝数、二级分枝长度呈显著性正相关;二级分枝数与二级分枝叶数、二级分枝长度呈极显著正相关;二级分枝直径与二级分枝叶数呈极显著正相关,与二级分枝长度呈显著正相关;二级分枝叶数与二级分枝长度呈极显著正相关。
3 结论与讨论
小蓬竹小生境内的形态变异在6个生境间存在差异,其中土面小生境大多数形态性状的变异系数最大,其次是石槽和石缝,表明不同的生境条件促进了小蓬竹的形态分化。通过对小蓬竹6个小生境的形态标记指标进行统计分析,各表型性状存在不同程度的分化,14个表型性状变异系数均值变化范围为5 7%~68 3%,变异系数较低都是叶片的相关指标,说明小蓬竹叶片性状受外界环境影响相对较小,遗传稳定性较高;变异系数最高的是二级分枝数,为68300%,表明小蓬竹生境间的形态变异大,遗传多样性很丰富,有利于小蓬竹的保护。聚类分析结果表明,6个小生境按形态性状可归为3类,石台和石洞聚为一类,石缝和石沟聚为一类,石槽和土面聚为一类,根据各小生境特征可以看出,聚为一类的生境都具有相似的生态因子(如土壤厚度、维持水分的能力等),说明小[CM(25]蓬竹的表型性状受环境的影响较大。主成分分析结果表明,前5个主成分的累积贡献率82 070%,可以代表原始因子所代表的大部分信息,其中第1主成分占41 139%,一级分枝叶数、节数、一级分枝数、二级分枝叶数的特征向量较大大于0 749,可作为其形态性状的重要主成分。相关性分析结果表明,株高和节平均值呈显著负相关,一级、二级分枝形态及一级、二级分枝叶片形态相互之间都存在着正相关性。
研究植物在其分布区域内各种生境下的形态变异,是了解遗传变异的重要线索,往往具有适应和进化上的意义 [18-20]。形态变异越大,可能存在的遗传变异越大,自然群体中保持大的变异贮存对群体是有利的,群体内多种基因型所对应的表型范围越广,群体在整体上适应环境的能力更强 [21-25]。根据小蓬竹丰富的表型变异,可以推断小蓬竹作为喀斯特的适生竹种,其应用和遗传改良前景是广阔的,这对开展小蓬竹天然群体遗传多样性和种质资源的保护、评价与利用具有重要的意义,为喀斯特造林材料的选育提供了较大的理论基础。本试验旨在对小蓬竹不同小生境下的天然居群的外部形态形状进行统计分析,但选取的指标有限,因此,今后对小蓬竹遗传改良品种的选择和应用中,应结合贵州喀斯特复杂的地形地貌,深入到分子水平,选取更多的变异指标进行系统的评价。
参考文献:
[1] 耿伯介 镰序竹属竹类植物在我国的新发现[J] 竹子研究汇刊,1986,5(2):28-40
[2]李永康,蓝开敏 贵州植物志[M] 成都:四川民族出版社,1988:271-272
[3]刘济明 贵州喀斯特地区小蓬竹生态特性研究[D] 北京:北京林业大学,2010
[4]蒙朝阳,谢元贵,刘济明,等 小蓬竹水土保持效应的研究[J] 安徽农业科学,2008,36(4):1431-1433
[5]Noel F,Machon N,Porcher E No genetic diversity at molecular markers and strong phenotypic plasticity in populations of Ranunculus nodiflorus,an endangered plant species in France[J] Annals of Botany,2007,99(6):1203-1212
[6] 沈 浩,刘登义 遗传多样性概述[J] 生物学杂志,2001,18(3):5-7
[7]King J N,Yeh F C,Heaman J H Selection of growth and yield traits in controlled crosses of coastal Douglas-fir[J] Silvae Genetica,1998,37(3/4):158-164
[8]Gomez O J,Blair M W,Frankow-Lindberg B E,et al Molecular and phenotypic diversity of common bean landraces from Nicaragua[J] Crop Science,2004,44(4):1412-1418
[9]施建敏,叶学华,陈伏生,等 竹类植物对异质生境的适应——表型可塑性[J] 生态学报,2014,34(20):5687-5695
[10] 周 超 小蓬竹克隆种群在不同喀斯特生境下的适应性[D] 贵阳:贵州大学,2008
[11]周 超,刘济明,蒙朝阳,等 不同喀斯特生境条件下小蓬竹构件生物量研究[J] 安徽农业科学,2008,36(5):1835-1836
[12]刘济明,池 馨,王 敏,等 喀斯特地区濒危小蓬竹的小生境变化特征[J] 贵州农业科学,2013,41(9):53-57
[13]徐雪娇,刘济明,徐国瑞,等 不同小生境中小蓬竹的含水率及生物量分配规律[J] 贵州农业科学,2010,38(10):163-166
[14]朱守谦 喀斯特森林生态研究:Ⅰ[M] 贵阳:贵州科技出版社,1993
[15]王 敏,刘济明,徐雪娇,等 濒危竹种小蓬竹群落不同小生境的小气候变化特征[J] 贵州农业科学,2013,41(6):78-82
[16]谢元贵,刘济明,陈 洪,等 不同喀斯特生境小蓬竹无性系构件研究[J] [HJ1 81mm]西南大学学报:自然科学版,2009,31(6):46-50
[17]潘庆杰 地涌金莲种群遗传多样性研究[D] 北京:中国林业科学研究院,2007
[18]杨树华,郭 宁,葛维亚,等 新疆东天山地区宽刺蔷薇居群表型多样性分析[J] 植物遗传资源学报,2013,14(3):455-461
[19]阎爱民,陈文新 苜蓿、草木樨、锦鸡儿根瘤菌的表型多样性分析[J] 生物多样性,1999,7(2):112-118
[20]杨 艳,刘 强,尹 翔,等 湖南白檀居群形态多样性及与环境的相关性[J] 植物遗传资源学报,2015,16(1):80-86
[21]马玉敏,陈学森,何天明,等 中国板栗3个野生居群部分表型性状的遗传多样性[J] 园艺学报,2008,35(12):1717-1726
[22]Baeilieri R,Ducousso A,Kremer A Genetic morphological ecological and phonological differentiation between Quercus petraea (Matt ) Label and Quercus robur L in a mixed stand of north-west of France[J] Silvae Genetica,1995,44(1):1-10
[23]王娅丽,李 毅,陈晓阳 祁连山青海云杉天然群体表型性状遗传多样性分析[J] 林业科学,2008,44(2):70-77
[24]李俊清,臧润国,蒋有绪 欧洲水青冈(Fagus sylvatical L )构筑型与形态多样性研究[J] 生态学报,2001,21(1):151-155
[25]Peterson B J,Graves W R,Sharma J Phenotypic and genotypic diversity of eastern leatherwood in five populations that span its geographic distribution[J] The American Midland Naturalist,2011,165(1):1-21
猜你喜欢
——以贵阳花溪公园为例
山地农业生物学报(2022年3期)2022-05-13
热带农业科学(2016年10期)2016-12-12
中国集体经济(2016年26期)2016-11-19
现代防御技术(2016年1期)2016-06-01
计算机工程(2015年8期)2015-07-03
通化师范学院学报(2014年8期)2014-06-12
计算机工程(2014年6期)2014-02-28
湖北社会科学(2011年11期)2011-10-30
