
苦荞EMS诱变群体的创建及农艺性状分析
2015-09-09 08:44邓琳琼张以忠
湖北农业科学 2015年14期
邓琳琼 张以忠

摘要:利用化学诱变剂甲基磺酸乙酯 (ethyl methane sulfonate,EMS) 诱变处理苦荞(Fagopyrum tataricum (L.) Gaertn)种子,对M2代突变材料进行生物学性状与农艺性状鉴定和分析,M3代材料进一步验证。结果表明,试验获得了叶片肥大、早熟、晚熟、小粒、矮秆和黄化6种稳定遗传的突变体。叶片肥大、早熟和晚熟突变体的株高、主茎分枝数、主茎节数、株粒数、株粒重和千粒重与对照间均无显著差异。矮秆突变体的株高和主茎分支数以及小粒突变体的株粒重和千粒重显著低于对照,其他性状与对照间差异不显著。黄化突变体的株高与对照无显著差异,主茎分枝数、主茎节数、株粒数、株粒重和千粒重均显著低于对照。
关键词:苦荞(Fagopyrum tataricum (L.) Gaertn);甲基磺酸乙酯;突变体;农艺性状
中图分类号:S517 文献标识码:A 文章编号:0439-8114(2015)14-3343-05
DOI:10.14088/j.cnki.issn0439-8114.2015.14.004
Construction and Agronomic Traits Analysis of Mutant Population from Tartary Buckwheat Induced by Ethyl Methane Sulfonate(EMS)
DENG Lin-qiong,ZHANG Yi-zhong
(Ecological Engineering School,Guizhou University of Engineering Science, Bijie 551700, Guizhou, China)
Abstract: The seeds of tartary buckwheat were treated by chemica1 mutagen ethyl methane sulfonate (EMS) to construct a mutant population. The biological and agronomic characters were identified and analyzed in M2 generation. M3 generation materials were further verified. The results showed that six types of stable genetic mutants(big leaf, premature, late maturity, small grain, dwarf and albino plant) were gotten. There were no significant differences for the plant height, the branches of main stem, the nodes of main stem, the seeds per plant, the weight per plant and the 1000-seeds weight between the big leaf mutant, premature mutant and late maturing mutant. The plant height and the branches of main stem of the dwarf mutant and the weight per plant and the 1000-seeds weight of the small grain mutant were significant lower than the control materials, whereas the other characters were no significant differences with the control materials. Though the plant height of the albino plant was no significant differences with the control materials, the branches of main stem, the nodes of main stem, the seeds per plant, the weight per plant and the 1000-seeds weight were significant lower than the control materials.
Key words: tartary buckwheat(Fagopyrum tataricum (L.) Gaertn);ethyl methane sulfonate;mutants;agronomic traits
突变体是某个性状发生可遗传变异或某个基因发生突变的材料,是遗传育种的重要资源,也是开展功能基因研究的重要材料,而且当今功能基因组学所取得的一些成果,也是建立在大量不同类型突变体的基础之上[1-3]。由于自发突变产生突变体的频率非常低,传统育种技术周期长,不能满足当前荞麦生产对优良品种的需求。化学诱变可以大大提高突变频率,加快新种质资源的创制速率。甲基磺酸乙酯(EMS)是目前使用最广泛、效果最好的化学诱变剂,由于其诱变方法简便、诱变频率高、诱变范围广、染色体畸变相对较少、可以对作物的某一种特殊性状及其品质进行改良,因多为显性突变体,易于进行筛选,因此在其他作物上已被广泛利用[4-6]。
苦荞(Fagopyrum. tataricum(L.) Gaertn)属蓼科(Polygnaceae)荞麦属(Fagopyrum Mill)2个栽培种之一,为一年生草本植物,其营养价值居粮食作物之首,而且还具有较高的药用价值,属药食两用作物,被营养学家们称为21世纪最有前途的绿色食品[7-9]。近年来,随着人们生活水平的不断提高,具有保健疗效的粮用药用作物荞麦和荞麦产品日益受到人们的喜爱和重视,但荞麦的产量和品质严重阻碍了荞麦的生产及发展。虽然诱发突变技术已广泛应用于植物品种改良,在水稻(Oryza sativa)[10-12]、小麦(Triticum aestivum Linn.)[13-15]、玉米(Zea mays)[16,17]等作物上获得了一批常规种质资源库中少见的突变材料,大大加快了传统作物育种的进程,也为功能作物育种和产品开发提供了重要技术手段和种质资源。但在荞麦育种研究方面,突变体研究报道很少,且均为物理方法进行的诱变,主要有李国柱等[18]、田小丽[19]、Yao等[20]、Samo等[21]、Breznik等[22]、王安虎等[23]的报道,这些利用物理方法对荞麦进行的诱变产生了一些较好的突变体,为荞麦的进一步研究及开发提供了一些材料。而利用EMS对荞麦的诱变研究目前尚未见报道。鉴于此,本研究利用化学诱变剂EMS对贵州地方栽培苦荞进行处理,对其突变体进行筛选和鉴定,创建苦荞突变体库,并对其农艺性状进行研究,以期为荞麦遗传育种、开发及利用提供理论依据。
1 材料与方法
1.1 供试材料
供试苦荞材料为贵州毕节苦荞(编号TA20111008001),来源于贵州工程应用技术学院生态工程学院荞麦种子资源库。
1.2 方法
1.2.1 突变材料创建 选用均匀一致的2 000粒饱满苦荞种子,室温下蒸馏水中浸泡18 h,滤纸吸干表面水分,室温下用质量分数为0.9%的化学诱变剂EMS(Sigma公司)处理17 h,自来水冲洗15 h,晾干,2012年7月25日播种于毕节市七星关区鸭池试验地(地理位置为东经105°16′571″,北纬27°13′322″,海拔1 550 m)。行距0.40 m,粒距0.30 m,田间常规管理,2012年11月28日单株收获M1代种子共793份。
1.2.2 M2代的种植与表型性状调查 2013年3月31日将793份M2代按株系播种于毕节市七星关区鸭池试验地,并设野生型苦荞作为对照,行距0.40 m,粒距0.30 m,全生育期每2~4 d进行田间调查一次,观察记载幼苗、叶、茎、花、育性、成熟期等性状,筛选各种形态突变体。并分别考察其株高、主茎分枝数、主茎节数、株粒数、株粒重、千粒重等。筛选各种形态性状变异材料,记录相应变异性状并编号,成熟后于2013年7月21日单株收获。
1.2.3 M3代种植与表型性状调查 2013年7月31日将在M2代筛选到的形态变异植株种子按株系(M3)播种于毕节市七星关区鸭池镇试验地,每个株系4株。在苦荞全生育期,根据M2代变异株的特点进行M3代株系突变性状的重复观测和鉴定,筛选记录遗传稳定的突变体。对仍然产生分离的M3代株系,分单株收获并对其M4代进行鉴定。
1.2.4 数据处理 采用Excel 2007软件计算平均数,采用SPSS11.5统计软件进行多重比较。
2 结果与分析
2.1 EMS诱变处理后Ml代性状的变异
经EMS处理后的2 000粒苦荞种子播种于试验地,出苗793株,出苗率为39.65%。在其全生育期,发现植株长势与对照差异较大(图1B)。主要表现在79.82%植株明显矮化、93.49%植株成熟期改变、6.90%的植株出现了部分叶片色泽、叶形异常,出现了1.92%小粒及2.30%不育及蚂蚁危害死亡等变异植株。另外,发现M1代表型变异多数不能遗传给M2代,最终得到的稳定遗传的突变体与所观察到的M1代的某些变异性状变化较大,可能是由于经EMS诱变后,EMS对M1代种子有一定的伤害,导致M1代植株出现性状的变化,这些变化多数不能遗传,在M2代得到了恢复。
2.2 M2代突变体的筛选及农艺性状分析
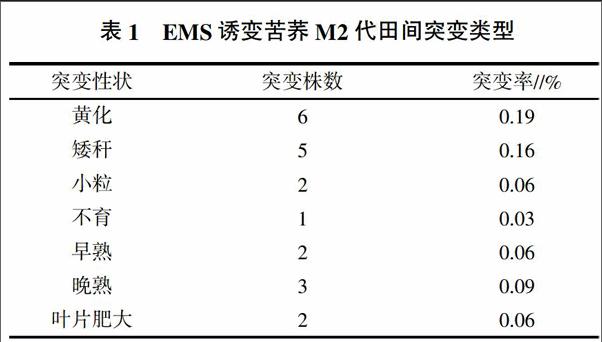
以野生型苦荞作为对照,对793份M2代按株系播种,每个株系4株,共3 172个单株,其中21株出现了突变,发生表型突变的频率为0.66%。主要表现在株高、叶片、种子、育性、成熟期等性状(表1)。
2.2.1 营养器官突变 在诱变群体中共观察到13株荞麦的营养器官发生了突变,突变频率为0.41%。叶色突变共6株,突变频率为0.19%,包括整片叶片黄化突变(图1C)、部分叶片黄化突变(图1D)和叶片镶嵌黄化突变(图1E)。矮秆突变(图1B)共5株,突变频率为0.16%,株高在20~47 cm之间,平均株高仅为36.33 cm。叶片肥大突变(图1F)共2株,突变频率为0.06%,叶片明显比正常植株叶片大且厚。
2.2.2 成熟期突变 在诱变群体中共观察到5株荞麦的成熟期发生了突变,突变频率为0.15%。包括早熟突变和晚熟突变。其中早熟突变共2株,突变频率为0.06%,成熟期提前15 d。晚熟突变共3株,突变频率为0.09%,成熟期推后20 d。
2.2.3 生殖器官突变 诱变群体中共观察到3株荞麦的生殖器官发生了突变,突变频率为0.09%。包括小粒突变和不育突变。其中小粒突变共2株(图1G),突变频率为0.06%,种子明显比正常植株种子小。不育突变1株,突变频率为0.03%,植株在苗期生长正常,可以开花,但花枯萎,无种子。
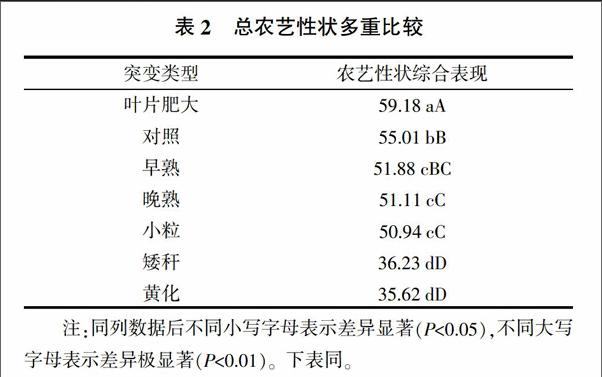
2.2.4 M2代突变体间及与对照间的总农艺性状分析 M2代突变体间及与对照间的株高、主茎分枝数、主茎节数、株粒数、株粒重和千粒重总农艺性状表现如表2所示。由表2可知,叶片肥大突变体总农艺性状值最高,为59.18,极显著高于对照及其他突变体。对照的总农艺性状值为55.01,极显著高于晚熟、小粒、矮秆和黄化突变体,显著高于早熟突变体。早熟、晚熟和小粒突变体间差异不显著,但均极显著高于矮秆和黄化突变体,而矮秆和黄化突变体间差异不显著。综合以上分析发现,就总农艺性状表现而言,叶片肥大突变体表现值最大,而黄化突变体的表现值最小。在0.05水平上,其农艺性状综合表现可将其排序为叶片肥大>对照>早熟、晚熟、小粒>矮秆、黄化。
2.2.5 M2代突变体间及与对照间的农艺性状分析 M2代突变体间及与对照间株高、主茎分枝数、主茎节数、株粒数、株粒重和千粒重的表现如表3所示。由表3可知,株高方面,叶片肥大、对照、早熟、晚熟、小粒和黄化突变体的株高差异不显著,但均极显著高于矮秆突变体。主茎分枝数方面,早熟、晚熟、小粒、叶片肥大突变体和对照间差异不显著,但早熟、晚熟、小粒、叶片肥大突变体均显著高于矮秆和黄化突变体,对照、矮秆和黄化突变体间差异不显著,且早熟突变体与矮秆和黄化突变体,晚熟突变体与黄化突变体之间的差异达到极显著水平。主茎节数方面,对照、晚熟、小粒、叶片肥大、早熟和矮秆突变体之间的差异不显著,而对照和晚熟突变体均显著高于黄化突变体,但未达到极显著水平。株粒数方面,叶片肥大、小粒突变体和对照之间差异不显著,但均显著高于黄化突变体,而早熟、晚熟、矮秆和黄化突变体间差异不显著。株粒重方面,叶片肥大、早熟、晚熟突变体和对照之间以及矮秆、小粒和黄化突变体之间差异均不显著,但叶片肥大突变体极显著高于矮秆、小粒和黄化突变体,对照显著高于小粒和黄化突变体,另外早熟和晚熟突变体也显著高于黄化突变体。千粒重方面,叶片肥大突变体和对照之间差异不显著,显著高于黄化和小粒突变体,极显著高于小粒突变体。矮秆、早熟、晚熟和黄化突变体间差异不显著,但均显著高于小粒突变体,且矮秆、早熟、晚熟与小粒突变体间的差异达极显著水平。
2.3 M3代突变体的表型鉴定
在M3代(部分M4代)再次对突变表型进行鉴定,有80.95%从M2代中筛选到的具有典型特征的突变体能够稳定遗传而不再发生分离。通过对这些类型的突变体M2代和M3代(部分M4代)突变表型遗传特点的观察分析,在最终得到的突变体中,包括黄化、矮秆、小粒、早熟、晚熟、叶片肥大6种突变体类型。
3 小结与讨论
突变体是某个性状发生可遗传变异或某个基因发生突变的材料,突变体的创制是进行植物功能基因组学和遗传育种研究的重要途径[1,24]。EMS诱变由于方法及条件简单,诱变频率高,染色体畸变相对较少,并能够在短时间内创制出大量突变体。因此其作为人工创造变异的一种手段越来越受到研究者的重视[25]。笔者首次利用化学诱变剂EMS对贵州地方栽培苦荞进行处理,对其产生的突变体进行筛选和鉴定,创制了一些具有价值的突变体。在M2代获得了黄化、矮秆、小粒、早熟、晚熟、叶片肥大6种类型的突变体,突变频率为0.66%,其中黄化突变频率最高,为0.19%,叶片肥大、早熟和小粒的突变频率次之,均为0.06%,而不育突变频率最小,仅为0.03%。进一步对M3代突变表型进行观察,发现80.95%从M2代中筛选到的突变体能够稳定遗传而未发生分离,最终得到黄化、矮秆、小粒、早熟、晚熟、叶片肥大6种类型的突变体。这6种类型突变体在株高、主茎节数、主茎分枝数、株粒数、株粒重和千粒重等农艺性状综合表现方面,叶片肥大突变体的综合表现值最大,对照次之,早熟、晚熟和小粒第三,而矮秆和黄化的最小。
在苦荞育种过程中,对农艺性状的选择十分重要,因为农艺性状的优劣对苦荞的高产性和稳产性有重要的影响,但是不同农艺性状对苦荞的产量影响不同。因此,本研究在对农艺性状进行综合分析的基础上,进一步对突变体间以及突变体与对照间的各农艺性状进行分析。结果表明,本研究所获得的6种类型的突变体中,叶片肥大突变体、早熟突变体与对照间在株高、主茎分枝数、主茎节数、株粒数、株粒重和千粒重等农艺性状方面均无显著差异。晚熟突变体除千粒重显著低于叶片肥大突变体外,本研究中的其他农艺性状均与叶片肥大突变体、早熟突变体和对照间无显著差异。矮秆突变体的株高显著低于对照,其他农艺性状与对照间差异不显著。小粒突变体株粒重和千粒重显著低于对照,株高等其他农艺性状与对照无显著差异。黄化突变体株高和主茎分枝数与对照间差异不显著,主茎节数、株粒数、株粒重和千粒重均显著低于对照。前人报道[26,27]发现,千粒重、单株粒数能有效地提高苦荞的产量水平,千粒重对株粒重的直接效应较大,是荞麦品种选育的主要目标性状和高产栽培的主攻方向。本研究中,叶片肥大突变体、早熟突变体与对照间的株粒数、千粒重和株粒重均无显著差异。因此该两种类型突变体在苦荞产量选育上可能优于其他突变体,而突变体之间以及突变体与对照之间的品质性状方面是否具有与农艺性状方面相同或相似的差异,有待进一步研究。
参考文献:
[1] 钱 前,程式华.水稻遗传学和功能基因组学[M].北京:科学出版社,2006.
[2] 张晓勤,薛大伟,周伟辉,等.用甲基磺酸乙酯(EMS)诱变的大麦浙农大3号突变体的筛选和鉴定[J].浙江大学学报(农业与生命科学版),2011,37(2):169-174.
[3] 郭建秋,雷全奎,杨小兰,等.植物突变体库的构建及突变体检测研究进展[J].河南农业科学,2010(6):150-155.
[4] ALONSO J M,STEPANOVA A N,LEISSE T J,et al.Genome-wide insertional mutagenesis of Arabidopsis thaliana[J].Science,2003,301:653-657.
[5] JEON J S,JANG S,LEE S,et al.Leafy hull sterile is a homeotic mutation in a rice MADS box gene affecting rice flower development[J]. Plant Cell,2000,12:871-884
[6] 樊双虎,郭文柱,路小铎,等.玉米EMS突变体库构建及突变体初步鉴定[J].安徽农业科学,2014, 42(11):3162-3165,3185.
[7] 张以忠,陈庆富.荞麦研究的现状与展望[J].种子,2004,23(3):39-42.
[8] 姜 涛,孔令聪,王光宇,等.安徽省苦荞麦种质资源引种观察及鉴定[J].农学学报,2013,3(7):8-10.
[9] 侯雅君,张宗文,吴 斌,等.苦荞种质资源AFLP标记遗传多样性分析[J].中国农业科学,2009,42(12):4166-4174.
[10] 李燕群,蒲 翔,李春梅,等.水稻507ys黄绿叶突变体的遗传鉴定与候选基因分析[J].中国农业科学,2014,47(2):221-229.
[11] 陈竹锋,卢嘉威,卢启清,等.优质水稻黄华占雄性不育突变体的筛选及初步分析[J].广东农业科学,2014,41(19):1-4.
[12] 罗洪发,查仁明,杨洪海,等.水稻突变体的创制[J].中国农学通报,2011,27(27):45-49.
[13] 张纪元,张平平,姚金保,等.以EMS诱变创制软质小麦宁麦9号高分子量谷蛋白亚基突变体[J].作物学报,2014,40(9):1579-1584.
[14] 杜丽芬,李明飞,刘录祥,等.一个化学诱变的小麦斑点叶突变体的生理和遗传分析[J].作物学报,2014,40(6):1020-1026.
[15] ERMELINDA B,FRANCESCO S,ANTONIO H L,et al.High resolution melting analysis for the detection of EMS induced mutations in wheat SBEIIa genes[J]. BMC Plant Biol,2011,11:156.
[16] 刘祥久,刘晓丽.利用EMS诱发玉米性状变异的效果[J].杂粮作物,2010,30(2):88-90.
[17] 杨 镇,刘晓丽,李 刚.EMS诱变剂对玉米自交系改造效果的研究[J].辽宁农业科学,2006(5):7-10.
[18] 李国柱,董艳辉,申慧芳,等.苦荞辐照突变体黄酮含量与PAL活性和农艺性状研究[J].激光生物学报,2010,19(4):546-551.
[19] 田小丽.60Co-γ射线辐照诱发苦荞突变体筛选高黄酮含量的种质资源[J].科技情报开发与经济,2008,18(23):150-152.
[20] YAO Y N,XUAN Z Y, HE Y M,et al. Principal component analysis of intraspecific responses of tartary buckwheat to UV-B radiation under field conditions[J]. Environ Exp Bot,2007,61(3):237-245.
[21] SAMO K,?譒TRUKELJ B,GABER?譒IK A. Rutin in buckwheat herbs grown at different UV-B radiation levels: comparison of two UV spectrophotometric and an HPLC method[J]. J Exp Bot, 2002, 53: 1801-1804.
[22] BREZNIK B,GERM M, GABERSCIK A,et al.Combined effects of elevated UV-B radiation and the addition of selenium on common (Fagopyrum esculentum Moench) and tartary [Fagopyrum tataricum(L.) Gaertn.] buckwheat[J]. Photosynthetica,2005,43(4):583-589.
[23] 王安虎,戴红燕,邓建平.60Co-γ射线辐射苦荞麦籽粒M2至M5代变异性状不稳定性研究[J].杂粮作物,2005,25(6):364-366.
[24] MIYAO A,IWASAKI Y,KITANO H,et al.A large-scale collection of phenotypic data describing an insertional mutant population to facilitate functional analysis of rice genes[J]. Plant Molecular Biology,2007,63(5):625-635.
[25] 罗洪发,查仁明,杨洪海,等.水稻突变体的创制[J].中国农学通报,2011,27(27):45-49.
[26] 杨明君,杨 嫒,郭忠贤,等.旱作苦荞麦籽粒产量与主要性状的相关分析[J].内蒙古农业科技,2010,(2):49-50.
[27] 杨玉霞,吴 卫,郑有良,等.苦荞主要农艺性状与单株籽粒产量的相关和通径分析[J].安徽农业科学,2008,36(16):6719-6721,6746.
猜你喜欢
农业科技通讯(2023年1期)2023-02-12
作物学报(2022年3期)2022-01-22
麦类作物学报(2021年5期)2021-09-23
课堂内外(高中版)(2021年9期)2021-01-17
商品与质量(2019年36期)2019-04-15
种子(2018年10期)2018-11-14
麦类作物学报(2018年9期)2018-10-09
中国果业信息(2017年4期)2017-05-09
大众文艺(2017年1期)2017-03-08
中国果菜(2016年9期)2016-03-01
